
Daily Early-Life Exposures to Diet Soda and Aspartame Are Associated with Autism in Males: A Case-Control Study

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Materials
2.2.1. Demographic and Neurodevelopmental Data Collected
2.2.2. Early-Life Exposures to Diet Sodas, Other Diet Drinks, Aspartame, and Other NNSs
2.3. Procedure
2.3.1. Case Definitions
2.3.2. Exposure Variables
2.4. Statistical Analyses
2.5. Sensitivity Analyses
3. Results
Sensitivity Analyses
4. Discussion
4.1. Non-Regressive Cases
4.2. Results from Earlier Studies in Adult Consumers and Gestationally Exposed Offspring
4.3. Sex Differences in Developmental Responses to Early-Life Exposures from Earlier Studies
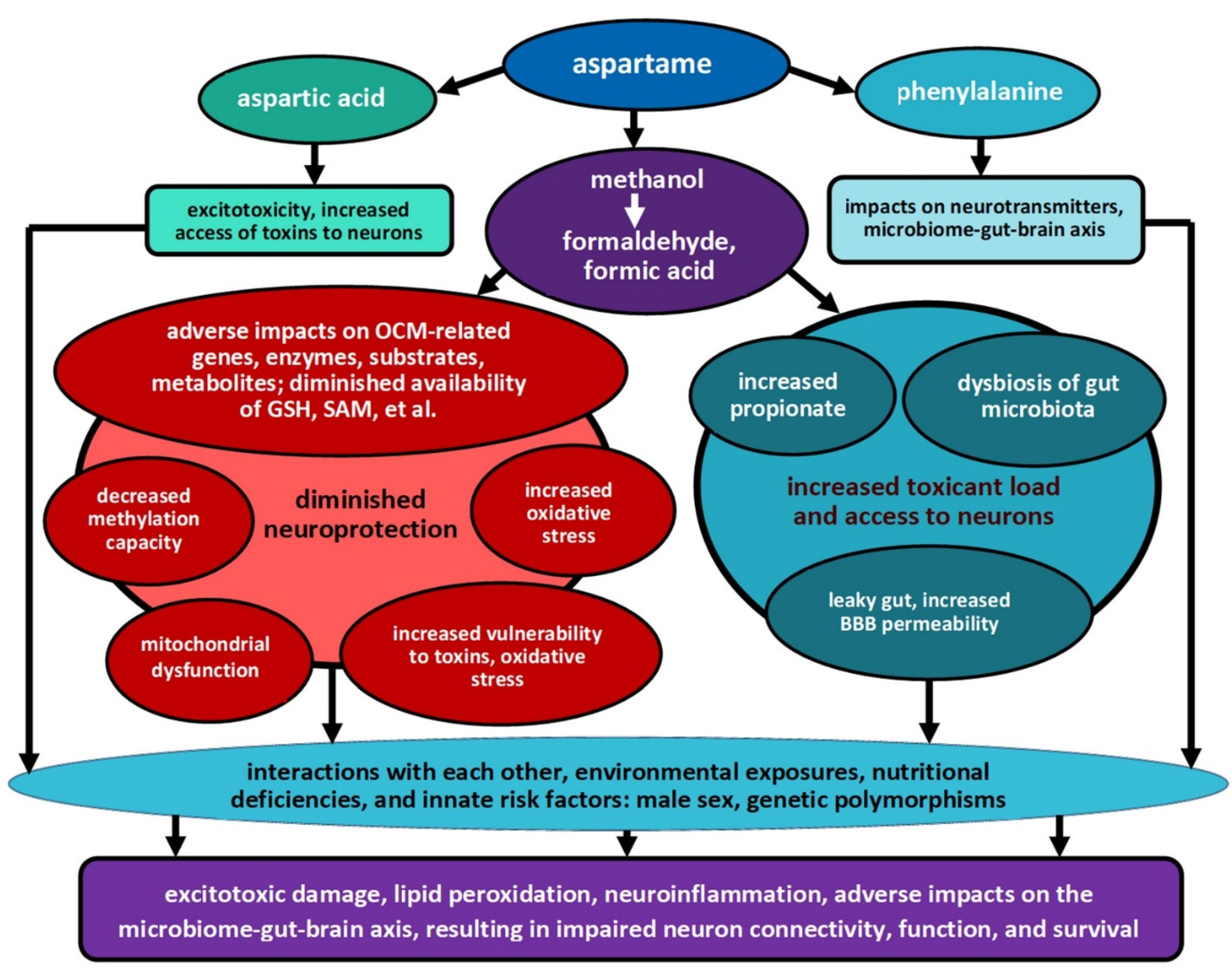
4.4. Is Dietary Methanol from Aspartame Contributing to These Associations?
4.5. Metabolic Impacts of Aspartame, Methanol, and Their Metabolites on Availability of Reduced Glutathione (GSH) and Other Products of One-Carbon Metabolism
4.6. Sensitivity Analyses
4.7. Future Directions
4.8. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nevison, C.; Blaxill, M.; Zahorodny, W. California Autism Prevalence Trends from 1931 to 2014 and Comparison to National ASD Data from IDEA and ADDM. J. Autism Dev. Disord. 2018, 48, 4103–4117. [Google Scholar] [CrossRef]
- Frye, R.E.; Rossignol, D.A. Mitochondrial Dysfunction Can Connect the Diverse Medical Symptoms Associated with Autism Spectrum Disorders. Pediatr. Res. 2011, 69, 41R–47R. [Google Scholar] [CrossRef]
- Maenner, M.J.; Warren, Z.; Williams, A.R.; Amoakohene, E.; Bakian, A.V.; Bilder, D.A.; Durkin, M.S.; Fitzgerald, R.T.; Furnier, S.M.; Hughes, M.M.; et al. Prevalence and Characteristics of Autism Spectrum Disorder among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2020. MMWR Surveill. Summ. 2023, 72, 1–14. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Neurobehavioural effects of developmental toxicity. Lancet Neurol. 2014, 13, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Tessing, J.; Lee, B.K.; Lyall, K. Maternal Dietary Factors and the Risk of Autism Spectrum Disorders: A Systematic Review of Existing Evidence. Autism Res. 2020, 13, 1634–1658. [Google Scholar] [CrossRef] [PubMed]
- Peretti, S.; Mariano, M.; Mazzocchetti, C.; Mazza, M.; Pino, M.C.; Verrotti Di Pianella, A.; Valenti, M. Diet: The keystone of autism spectrum disorder? Nutr. Neurosci. 2019, 22, 825–839. [Google Scholar] [CrossRef]
- Walton, R.G.; Monte, W.C. Dietary methanol and autism. Med. Hypotheses 2015, 85, 441–446. [Google Scholar] [CrossRef]
- Butchko, H.H.; Stargel, W. Aspartame: Scientific Evaluation in the Postmarketing Period. Regul. Toxicol. Pharmacol. 2001, 34, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Putnam, J.J.; Allshouse, J.E. Food Consumption, Prices, and Expenditures, 1970–1997; Economic Research Service, U.S. Department of Agriculture: Washington, DC, USA, 1999. [Google Scholar]
- Centers for Disease Control. Evaluation of Consumer Complaints Related to Aspartame Use. Morb. Mortal. Wkly. Rep. 1984, 33, 605–607. [Google Scholar]
- Bradstock, M.K.; Serdula, M.K.; Marks, J.S.; Barnard, R.J.; Crane, N.T.; Remington, P.L.; Trowbridge, F.L. Evaluation of reactions to food additives: The aspartame experience. Am. J. Clin. Nutr. 1986, 43, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Wurtman, R. Aspartame: Possible effect on seizure susceptibility. Lancet 1985, 326, 1060. [Google Scholar] [CrossRef]
- Walton, R.G. Seizure and mania after high intake of aspartame. Psychosomatics 1986, 27, 218–220. [Google Scholar] [CrossRef]
- Spiers, P.S.D.; Sabounjian, L.; Lieberman, H.; Wurtman, R.; Duguid, J.; McCarten, R.; Lyden, M. Aspartame and human behavior: Cognitive and behavioral observations. In Dietary Phenylalanine and Brain Function; Wurtman, R.-W.E., Ed.; Burkhäuser Boston: Boston, MA, USA, 1988; pp. 169–178. [Google Scholar]
- Walton, R.G. The possible role of aspartame in seizure induction. In Dietary Phenylalanine and Brain Function; Wurtman, R.J., Ritter-Walker, E., Eds.; Birkhauser: Boston, MA, USA, 1988; pp. 159–162. [Google Scholar]
- Lipton, R.B.; Newman, L.C.; Cohen, J.S.; Solomon, S. Aspartame as a Dietary Trigger of Headache. Headache J. Head Face Pain 1989, 29, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Walton, R.G.; Hudak, R.; Green-Waite, R.J. Adverse reactions to aspartame: Double-blind challenge in patients from a vulnerable population. Biol. Psychiatry 1993, 34, 13–17. [Google Scholar] [CrossRef]
- Van den Eeden, S.K.; Koepsell, T.D.; Longstreth, W.T.; Van Belle, G.; Daling, J.R.; McKnight, B. Aspartame ingestion and headaches: A randomized crossover trial. Neurology 1994, 44, 1787–1793. [Google Scholar] [CrossRef]
- Roberts, H.J. Aspartame Disease: An Ignored Epidemic; Sunshine Sentinel Press, Inc.: West Palm Beach, FL, USA, 2001. [Google Scholar]
- Lindseth, G.N.; Coolahan, S.E.; Petros, T.V.; Lindseth, P.D. Neurobehavioral Effects of Aspartame Consumption. Res. Nurs. Health 2014, 37, 185–193. [Google Scholar] [CrossRef]
- Guo, X.; Park, Y.; Freedman, N.D.; Sinha, R.; Hollenbeck, A.R.; Blair, A.; Chen, H. Sweetened Beverages, Coffee, and Tea and Depression Risk among Older US Adults. PLoS ONE 2014, 9, e94715. [Google Scholar] [CrossRef]
- Pase, M.P.; Himali, J.J.; Beiser, A.S.; Aparicio, H.J.; Satizabal, C.L.; Vasan, R.S.; Seshadri, S.; Jacques, P.F. Sugar- and Artificially Sweetened Beverages and the Risks of Incident Stroke and Dementia: A Prospective Cohort Study. Stroke 2017, 48, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Sylvetsky, A.C.; Jin, Y.; Clark, E.J.; Welsh, J.A.; Rother, K.I.; Talegawkar, S.A. Consumption of Low-Calorie Sweeteners among Children and Adults in the United States. J. Acad. Nutr. Diet. 2017, 117, 441–448.e2. [Google Scholar] [CrossRef] [PubMed]
- Christian, B.; McConnaughey, K.; Bethea, E.; Brantley, S.; Coffey, A.; Hammond, L.; Harrell, S.; Metcalf, K.; Muehlenbein, D.; Spruill, W.; et al. Chronic aspartame affects T-maze performance, brain cholinergic receptors and Na+, K+-ATPase in rats. Pharmacol. Biochem. Behav. 2004, 78, 121–127. [Google Scholar] [CrossRef]
- Abdel-Salam, O.M.; Salem, N.A.; El-Shamarka, M.E.; Hussein, J.S.; Ahmed, N.A.; El-Nagar, M.E. Studies on the effects of aspartame on memory and oxidative stress in brain of mice. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 2092–2101. [Google Scholar]
- Abu-Taweel, G.M.; Zyadah, M.A.; Ajarem, J.S.; Ahmad, M. Cognitive and biochemical effects of monosodium glutamate and aspartame, administered individually and in combination in male albino mice. Neurotoxicol. Teratol. 2014, 42, 60–67. [Google Scholar] [CrossRef]
- Erbaş, O.; Erdoğan, M.A.; Khalilnezhad, A.; Solmaz, V.; Gürkan, F.T.; Yiğittürk, G.; Eroglu, H.A.; Taskiran, D. Evaluation of long-term effects of artificial sweeteners on rat brain: A biochemical, behavioral, and histological study. J. Biochem. Mol. Toxicol. 2018, 32, e22053. [Google Scholar] [CrossRef]
- Onaolapo, A.Y.; Onaolapo, O.J.; Nwoha, P.U. Alterations in behaviour, cerebral cortical morphology and cerebral oxidative stress markers following aspartame ingestion. J. Chem. Neuroanat. 2016, 78, 42–56. [Google Scholar] [CrossRef]
- Choudhary, A.K.; Pretorius, E. Revisiting the safety of aspartame. Nutr. Rev. 2017, 75, 718–730. [Google Scholar] [CrossRef] [PubMed]
- Onaolapo, A.Y.; Abdusalam, S.Z.; Onaolapo, O.J. Silymarin attenuates aspartame-induced variation in mouse behaviour, cerebrocortical morphology and oxidative stress markers. Pathophysiology 2017, 24, 51–62. [Google Scholar] [CrossRef]
- Butchko, H.H.; Stargel, W.W.; Comer, C.P.; Mayhew, D.A.; Benninger, C.; Blackburn, G.L.; de Sonneville, L.M.J.; Geha, R.S.; Hertelendy, Z.; Koestner, A.; et al. Aspartame: Review of Safety. Regul. Toxicol. Pharmacol. 2002, 35, S1–S93. [Google Scholar] [CrossRef]
- Magnuson, B.A.; Burdock, G.A.; Doull, J.; Kroes, R.M.; Marsh, G.M.; Pariza, M.W.; Spencer, P.S.; Waddell, W.J.; Walker, R.; Williams, G.M. Aspartame: A Safety Evaluation Based on Current Use Levels, Regulations, and Toxicological and Epidemiological Studies. Crit. Rev. Toxicol. 2007, 37, 629–727. [Google Scholar] [CrossRef] [PubMed]
- Marinovich, M.; Galli, C.L.; Bosetti, C.; Gallus, S.; La Vecchia, C. Aspartame, low-calorie sweeteners and disease: Regulatory safety and epidemiological issues. Food Chem. Toxicol. 2013, 60, 109–115. [Google Scholar] [CrossRef] [PubMed]
- EFSA ANS Panel (EFSA Panel on Food Additives and Nutrient Sources added to Food). Scientific Opinion on the re-evaluation of aspartame (E 951) as a food additive. EFSA J. 2013, 11, 263. [Google Scholar]
- Soffritti, M.; Belpoggi, F.; Tibaldi, E.; Degli Esposti, D.; Lauriola, M. Life-Span Exposure to Low Doses of Aspartame Beginning during Prenatal Life Increases Cancer Effects in Rats. Environ. Health Perspect. 2007, 115, 1293–1297. [Google Scholar] [CrossRef]
- Abdo, K.M.; Camargo Jr, C.A.; Davis, D.; Egilman, D.; Epstein, S.S.; Froines, J.; Hattis, D.; Hooper, K.; Huff, J.; Infante, P.F.; et al. Letter to U.S. FDA commissioner. Questions about the safety of the artificial sweetener aspartame. Int. J. Occup. Env. Environ. Health 2007, 13, 449–450. [Google Scholar]
- Millstone, E.P.; Dawson, E. EFSA’s toxicological assessment of aspartame: Was it even-handedly trying to identify possible unreliable positives and unreliable negatives? Arch. Public Health 2019, 77, 34. [Google Scholar] [CrossRef]
- Humphries, P.; Pretorius, E.; Naudé, H. Direct and indirect cellular effects of aspartame on the brain. Eur. J. Clin. Nutr. 2007, 62, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Wurtman, R.J. Aspartame effects on brain serotonin. Am. J. Clin. Nutr. 1987, 45, 799–801. [Google Scholar] [CrossRef] [PubMed]
- Olney, J.W.; Ho, O.-L. Brain Damage in Infant Mice following Oral Intake of Glutamate, Aspartate or Cysteine. Nature 1970, 227, 609–611. [Google Scholar] [CrossRef]
- Blaylock, R.L. Excitotoxins: The Taste That Kills; Health Press: Santa Fe, NM, USA, 1997. [Google Scholar]
- Borkum, J.M. Migraine Triggers and Oxidative Stress: A Narrative Review and Synthesis. Headache J. Head Face Pain 2016, 56, 12–35. [Google Scholar] [CrossRef]
- Monte, W.C. Methanol: A chemical Trojan horse as the root of the inscrutable U. Med. Hypotheses 2010, 74, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Iyyaswamy, A.; Rathinasamy, S. Effect of chronic exposure to aspartame on oxidative stress in the brain of albino rats. J. Biosci. 2012, 37, 679–688. [Google Scholar] [CrossRef]
- Ashok, I.; Sheeladevi, R. Biochemical responses and mitochondrial mediated activation of apoptosis on long-term effect of aspartame in rat brain. Redox Biol. 2014, 2, 820–831. [Google Scholar] [CrossRef]
- Ashok, I.; Sheeladevi, R.; Wankhar, D. Acute effect of aspartame-induced oxidative stress in Wistar albino rat brain. J. Biomed. Res. 2015, 29, 390–396. [Google Scholar] [PubMed]
- Songur, A.; Ozen, O.A.; Sarsilmaz, M. The Toxic Effects of Formaldehyde on the Nervous System. Rev. Env. Environ. Contam. Toxicol. 2010, 203, 105–118. [Google Scholar]
- Tulpule, K.; Dringen, R. Formaldehyde in brain: An overlooked player in neurodegeneration? J. Neurochem. 2013, 127, 7–21. [Google Scholar] [CrossRef]
- Abhilash, M.; Paul, M.V.S.; Varghese, M.V.; Nair, R.H. Long-term consumption of aspartame and brain antioxidant defense status. Drug Chem. Toxicol. 2013, 36, 135–140. [Google Scholar] [CrossRef]
- Finamor, I.A.; Ourique, G.M.; Pês, T.S.; Saccol, E.M.H.; Bressan, C.A.; Scheid, T.; Baldisserotto, B.; Llesuy, S.F.; Partata, W.A.; Pavanato, M.A. The Protective Effect of N-Acetylcysteine on Oxidative Stress in the Brain Caused by the Long-Term Intake of Aspartame by Rats. Neurochem. Res. 2014, 39, 1681–1690. [Google Scholar] [CrossRef] [PubMed]
- Adaramoye, O.A.; Akanni, O.O. Effects of long-term administration of aspartame on biochemical indices, lipid profile and redox status of cellular system of male rats. J. Basic Clin. Physiol. Pharmacol. 2016, 27, 29–37. [Google Scholar] [CrossRef]
- Finamor, I.; Pérez, S.; Bressan, C.A.; Brenner, C.E.; Rius-Pérez, S.; Brittes, P.C.; Cheiran, G.; Rocha, M.I.; da Veiga, M.; Sastre, J.; et al. Chronic aspartame intake causes changes in the trans-sulphuration pathway, glutathione depletion and liver damage in mice. Redox Biol. 2017, 11, 701–707. [Google Scholar] [CrossRef]
- Frye, R.E.; James, S.J. Metabolic pathology of autism in relation to redox metabolism. Biomark. Med. 2014, 8, 321–330. [Google Scholar] [CrossRef]
- Lu, S.C. Regulation of glutathione synthesis. Mol. Asp. Med. 2009, 30, 42–59. [Google Scholar] [CrossRef]
- Main, P.A.; Angley, M.T.; O’Doherty, C.E.; Thomas, P.; Fenech, M. The potential role of the antioxidant and detoxification properties of glutathione in autism spectrum disorders: A systematic review and meta-analysis. Nutr. Metab. 2012, 9, 35. [Google Scholar] [CrossRef]
- García-Giménez, J.L.; Ibañez-Cabellos, J.S.; Seco-Cervera, M.; Pallardó, F.V. Glutathione and cellular redox control in epigenetic regulation. Free. Radic. Biol. Med. 2014, 75 (Suppl. S1), S3. [Google Scholar] [CrossRef]
- Horio, Y.; Sun, Y.; Liu, C.; Saito, T.; Kurasaki, M. Aspartame-induced apoptosis in PC12 cells. Environ. Toxicol. Pharmacol. 2014, 37, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Ashok, I.; Sheeladevi, R. Oxidant stress evoked damage in rat hepatocyte leading to triggered nitric oxide synthase (NOS) levels on long term consumption of aspartame. J. Food Drug Anal. 2015, 23, 679–691. [Google Scholar] [CrossRef]
- Iyaswamy, A.; Kammella, A.K.; Thavasimuthu, C.; Wankupar, W.; Dapkupar, W.; Shanmugam, S.; Rajan, R.; Rathinasamy, S. Oxidative stress evoked damages leading to attenuated memory and inhibition of NMDAR-CaMKII-ERK/CREB signalling on consumption of aspartame in rat model. J. Food Drug Anal. 2018, 26, 903–916. [Google Scholar] [CrossRef]
- Choudhary, A.K.; Lee, Y.Y. Neurophysiological symptoms and aspartame: What is the connection? Nutr. Neurosci. 2018, 21, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.K. Aspartame: Should Individuals with Type II Diabetes be Taking it? Curr. Diabetes Rev. 2018, 14, 350–362. [Google Scholar] [CrossRef]
- Collison, K.S.; Inglis, A.; Shibin, S.; Saleh, S.; Andres, B.; Ubungen, R.; Thiam, J.; Mata, P.; Al-Mohanna, F.A. Effect of developmental NMDAR antagonism with CGP 39551 on aspartame-induced hypothalamic and adrenal gene expression. PLoS ONE 2018, 13, e0194416. [Google Scholar]
- Abdel-Salam, O.M.E.; Salem, N.A.; Hussein, J.S. Effect of Aspartame on Oxidative Stress and Monoamine Neurotransmitter Levels in Lipopolysaccharide-Treated Mice. Neurotox. Res. 2012, 21, 245–255. [Google Scholar] [CrossRef]
- Halldorsson, T.I.; Strøm, M.; Petersen, S.B.; Olsen, S.F. Intake of artificially sweetened soft drinks and risk of preterm delivery: A prospective cohort study in 59,334 Danish pregnant women. Am. J. Clin. Nutr. 2010, 92, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Palmnäs, M.S.A.; Cowan, T.E.; Bomhof, M.R.; Su, J.; Reimer, R.A.; Vogel, H.J.; Hittel, D.S.; Shearer, J. Low-Dose Aspartame Consumption Differentially Affects Gut Microbiota-Host Metabolic Interactions in the Diet-Induced Obese Rat. PLoS ONE 2014, 9, e109841. [Google Scholar]
- Gul, S.S.; Hamilton, A.R.L.; Munoz, A.R.; Phupitakphol, T.; Liu, W.; Hyoju, S.K.; Economopoulos, K.P.; Morrison, S.; Hu, D.; Zhang, W.; et al. Inhibition of the gut enzyme intestinal alkaline phosphatase may explain how aspartame promotes glucose intolerance and obesity in mice. Appl. Physiol. Nutr. Metab. 2017, 42, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhao, Y.; Jiang, X.; Li, R.; Xie, H.; Ge, L.; Xie, B.; Yang, X.; Zhang, L. Exposure to Formaldehyde Perturbs the Mouse Gut Microbiome. Genes 2018, 9, 192. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, R.; Shehreen, S.; Shahriar, S.; Rahman, S.; Akhteruzzaman, S.; Sajib, A.A. Non-Caloric Artificial Sweeteners Modulate the Expression of Key Metabolic Genes in the Omnipresent Gut Microbe Escherichia coli. Microb. Physiol. 2019, 29, 43–56. [Google Scholar] [CrossRef]
- Frankenfeld, C.L.; Sikaroodi, M.; Lamb, E.; Shoemaker, S.; Gillevet, P.M. High-intensity sweetener consumption and gut microbiome content and predicted gene function in a cross-sectional study of adults in the United States. Ann. Epidemiol. 2015, 25, 736–742.e4. [Google Scholar] [CrossRef] [PubMed]
- Gerasimidis, K.; Bryden, K.; Chen, X.; Papachristou, E.; Verney, A.; Roig, M.; Hansen, R.; Nichols, B.; Papadopoulou, R.; Parrett, A. The impact of food additives, artificial sweeteners and domestic hygiene products on the human gut microbiome and its fibre fermentation capacity. Eur. J. Nutr. 2019, 59, 3213–3230. [Google Scholar] [CrossRef]
- Nettleton, J.E.; Cho, N.A.; Klancic, T.; Nicolucci, A.C.; Shearer, J.; Borgland, S.L.; Johnston, L.A.; Ramay, H.R.; Tuplin, E.N.; Chleilat, F.; et al. Maternal low-dose aspartame and stevia consumption with an obesogenic diet alters metabolism, gut microbiota and mesolimbic reward system in rat dams and their offspring. Gut 2020, 69, 1807–1817. [Google Scholar] [CrossRef]
- Nettleton, J.E.; Reimer, R.A.; Shearer, J. Reshaping the gut microbiota: Impact of low calorie sweeteners and the link to insulin resistance? Physiol. Behav. 2016, 164, 488–493. [Google Scholar] [CrossRef]
- Suez, J.; Cohen, Y.; Valdés-Mas, R.; Mor, U.; Dori-Bachash, M.; Federici, S.; Zmora, N.; Leshem, A.; Heinemann, M.; Linevsky, R.; et al. Personalized microbiome-driven effects of non-nutritive sweeteners on human glucose tolerance. Cell 2022, 185, 3307–3328.e19. [Google Scholar]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef]
- MacFabe, D.F. Enteric short-chain fatty acids: Microbial messengers of metabolism, mitochondria, and mind: Implications in autism spectrum disorders. Microb. Ecol. Health Dis. 2015, 26, 28177. [Google Scholar] [CrossRef]
- El-Ansary, A.K.; Ben Bacha, A.; Kotb, M. Etiology of autistic features: The persisting neurotoxic effects of propionic acid. J. Neuroinflammation 2012, 9, 74. [Google Scholar] [PubMed]
- El-Ansary, A.; Al-Salem, H.S.; Asma, A.; Al-Dbass, A. Glutamate excitotoxicity induced by orally administered propionic acid, a short chain fatty acid can be ameliorated by bee pollen. Lipids Health Dis. 2017, 16, 96. [Google Scholar] [PubMed]
- Frye, R.E.; Nankova, B.; Bhattacharyya, S.; Rose, S.; Bennuri, S.C.; MacFabe, D.F. Modulation of Immunological Pathways in Autistic and Neurotypical Lymphoblastoid Cell Lines by the Enteric Microbiome Metabolite Propionic Acid. Front. Immunol. 2017, 8, 1670. [Google Scholar] [PubMed]
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.D.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 2013, 18, 666–673. [Google Scholar]
- Clarke, G.; O’Mahony, S.; Dinan, T.; Cryan, J. Priming for health: Gut microbiota acquired in early life regulates physiology, brain and behaviour. Acta Paediatr. 2014, 103, 812–819. [Google Scholar]
- The Interactive Autism Network: Research Overview: Kennedy Krieger Institute. 2023. Available online: https://iancommunity.org/cs/ian_research/overview (accessed on 6 August 2023).
- Interactive Autism Network: Patient-Centered Outcomes Research Institute. 2023. Available online: https://www.pcori.org/research-results/2015/interactive-autism-network#section_professional_abstract (accessed on 6 August 2023).
- Heilbrun, L.P.; Palmer, R.F.; Jaen, C.R.; Svoboda, M.D.; Perkins, J.; Miller, C.S. Maternal Chemical and Drug Intolerances: Potential Risk Factors for Autism and Attention Deficit Hyperactivity Disorder (ADHD). J. Am. Board Fam. Med. 2015, 28, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Palmer, R.F.; Heilbrun, L.; Camann, D.; Yau, A.; Schultz, S.; Elisco, V.; Tapia, B.; Garza, N.; Miller, C. Organic Compounds Detected in Deciduous Teeth: A Replication Study from Children with Autism in Two Samples. J. Environ. Public Health 2015, 2015, 862414. [Google Scholar]
- Franz, M. Diet Soft Drinks; Diabetes Self-Management, Rapaport Publishing, Inc.: New York, NY, USA, 2010. [Google Scholar]
- Stein, P.J. The Sweetness of Aspartame: A Biochemistry Lab for Health Science Chemistry Courses. J. Chem. Educ. 1997, 74, 1112. [Google Scholar] [CrossRef]
- Noss, A. Household Income for States: 2009 and 2010; American Community Survey Briefs, Report Number ACSBR/10-02; U.S. Census Bureau, Department, Economics and Statistics Administration: Washington, DC, USA, 2011; p. 5. Available online: https://www.census.gov/library/publications/2011/acs/acsbr10-02.html (accessed on 6 August 2023).
- Sylvetsky, A.C.; Figueroa, J.; Rother, K.I.; Goran, M.I.; Welsh, J.A. Trends in Low-Calorie Sweetener Consumption among Pregnant Women in the United States. Curr. Dev. Nutr. 2019, 3, nzz004. [Google Scholar]
- Azad, M.B.; Sharma, A.K.; de Souza, R.J.; Dolinsky, V.W.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; Lefebvre, D.L.; Sears, M.R.; et al. Association Between Artificially Sweetened Beverage Consumption During Pregnancy and Infant Body Mass Index. JAMA Pediatr. 2016, 170, 662–670. [Google Scholar] [CrossRef]
- Palatnik, A.; Moosreiner, A.; Stichelen, S.O.-V. Consumption of non-nutritive sweeteners during pregnancy. Am. J. Obstet. Gynecol. 2020, 223, 211–218. [Google Scholar] [PubMed]
- Boterberg, S.; Charman, T.; Marschik, P.B.; Bölte, S.; Roeyers, H. Regression in autism spectrum disorder: A critical overview of retrospective findings and recommendations for future research. Neurosci. Biobehav. Rev. 2019, 102, 24–55. [Google Scholar]
- Ozonoff, S.; Iosif, A.-M. Changing conceptualizations of regression: What prospective studies reveal about the onset of autism spectrum disorder. Neurosci. Biobehav. Rev. 2019, 100, 296–304. [Google Scholar]
- Buso, M.E.C.; Brouwer-Brolsma, E.M.; Naomi, N.D.; Ngo, J.; Soedamah-Muthu, S.S.; Mavrogianni, C.; Harrold, J.A.; Halford, J.C.G.; Raben, A.; Geleijnse, J.M.; et al. Sugar and low/no-calorie-sweetened beverage consumption and associations with body weight and waist circumference changes in five European cohort studies: The SWEET project. Eur. J. Nutr. 2023; Online ahead of print. [Google Scholar]
- Naomi, N.D.; Ngo, J.; Brouwer-Brolsma, E.M.; Buso, M.E.C.; Soedamah-Muthu, S.S.; Pérez-Rodrigo, C.; Harrold, J.A.; Halford, J.C.G.; Raben, A.; Geleijnse, J.M.; et al. Sugar-sweetened beverages, low/no-calorie beverages, fruit juice and non-alcoholic fatty liver disease defined by fatty liver index: The SWEET project. Nutr. Diabetes 2023, 13, 6. [Google Scholar] [PubMed]
- Azad, M.B.; Abou-Setta, A.M.; Chauhan, B.F.; Rabbani, R.; Lys, J.; Copstein, L.; Mann, A.; Jeyaraman, M.M.; Reid, A.E.; Fiander, M.; et al. Nonnutritive Sweeteners and Cardiometabolic Health: A Systematic Review and Meta-Analysis of Randomized Controlled Trials and Prospective Cohort Studies. CMAJ 2017, 189, E929–E939. [Google Scholar] [CrossRef] [PubMed]
- Halasa, B.C.; Sylvetsky, A.C.; Conway, E.M.; Shouppe, E.L.; Walter, M.F.; Walter, P.J.; Cai, H.; Hui, L.; Rother, K.I. Non-Nutritive Sweeteners in Human Amniotic Fluid and Cord Blood: Evidence of Transplacental Fetal Exposure. Am. J. Perinatol. 2021, 40, 1286–1291. [Google Scholar]
- Zhu, Y.; Olsen, S.F.; Mendola, P.; Halldorsson, T.I.; Rawal, S.; Hinkle, S.N.; Yeung, E.H.; Chavarro, J.E.; Grunnet, L.G.; Granström, C.; et al. Maternal consumption of artificially sweetened beverages during pregnancy, and offspring growth through 7 years of age: A prospective cohort study. Leuk. Res. 2017, 46, 1499–1508. [Google Scholar]
- Azad, M.B.; Archibald, A.; Tomczyk, M.M.; Head, A.; Cheung, K.G.; de Souza, R.J.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Moraes, T.J.; et al. Nonnutritive sweetener consumption during pregnancy, adiposity, and adipocyte differentiation in offspring: Evidence from humans, mice, and cells. Int. J. Obes. 2020, 44, 2137–2148. [Google Scholar]
- Plows, J.F.; Aris, I.M.; Rifas-Shiman, S.L.; Goran, M.I.; Oken, E. Associations of maternal non-nutritive sweetener intake during pregnancy with offspring body mass index and body fat from birth to adolescence. Int. J. Obes. 2022, 46, 186–193. [Google Scholar]
- Kern, J.K.; Geier, D.A.; Homme, K.G.; King, P.G.; Bjørklund, G.; Chirumbolo, S.; Geier, M.R. Developmental neurotoxicants and the vulnerable male brain: A systematic review of suspected neurotoxicants that disproportionally affect males. Acta Neurobiol. Exp. 2017, 77, 269–296. [Google Scholar]
- Bale, T.L. The placenta and neurodevelopment: Sex differences in prenatal vulnerability. Dialog. Clin. Neurosci. 2016, 18, 459–464. [Google Scholar] [CrossRef]
- James, S.J. Autism and Folate-dependent One-carbon Metabolism: Serendipity and Critical Branch-point Decisions in Science. Glob. Adv. Health Med. 2013, 2, 48–51. [Google Scholar] [CrossRef]
- James, S.J.; Melnyk, S.; Jernigan, S.; Cleves, M.A.; Halsted, C.H.; Wong, D.H.; Cutler, P.; Bock, K.; Boris, M.; Bradstreet, J.J.; et al. Metabolic endophenotype and related genotypes are associated with oxidative stress in children with autism. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2006, 141, 947–956. [Google Scholar] [CrossRef]
- Abhilash, M.; Paul, M.S.; Varghese, M.V.; Nair, R.H. Effect of long term intake of aspartame on antioxidant defense status in liver. Food Chem. Toxicol. 2011, 49, 1203–1207. [Google Scholar] [CrossRef] [PubMed]
- Ashok, I.; Poornima, P.S.; Wankhar, D.; Ravindran, R.; Sheeladevi, R. Oxidative stress evoked damages on rat sperm and attenuated antioxidant status on consumption of aspartame. Int. J. Impot. Res. 2017, 29, 164–170. [Google Scholar] [CrossRef]
- Mezzelani, A.; Landini, M.; Facchiano, F.; Raggi, M.E.; Villa, L.; Molteni, M.; De Santis, B.; Brera, C.; Caroli, A.M.; Milanesi, L.; et al. Environment, dysbiosis, immunity and sex-specific susceptibility: A translational hypothesis for regressive autism pathogenesis. Nutr. Neurosci. 2015, 18, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Vuong, H.E.; Hsiao, E.Y. Emerging Roles for the Gut Microbiome in Autism Spectrum Disorder. Biol. Psychiatry 2017, 81, 411–423. [Google Scholar] [PubMed]
- Ghanizadeh, A.; Akhondzadeh, S.; Hormozi, M.; Makarem, A.; Abotorabi-Zarchi, M.; Firoozabadi, A. Glutathione-Related Factors and Oxidative Stress in Autism, A Review. Curr. Med. Chem. 2012, 19, 4000–4005. [Google Scholar] [CrossRef]
- Li, M.; Fallin, M.D.; Riley, A.; Landa, R.; Walker, S.O.; Silverstein, M.; Caruso, D.; Pearson, C.; Kiang, S.; Dahm, J.L.; et al. The Association of Maternal Obesity and Diabetes with Autism and Other Developmental Disabilities. Pediatrics 2016, 137, e20152206. [Google Scholar] [CrossRef]
- Lei, X.Y.; Li, Y.J.; Ou, J.J.; Li, Y.M. Association between parental body mass index and autism spectrum disorder: A systematic review and meta-analysis. Eur. Child Adolesc. Psychiatry 2018, 28, 933–947. [Google Scholar] [CrossRef]
- Carpita, B.; Muti, D.; Dell’Osso, L. Oxidative Stress, Maternal Diabetes, and Autism Spectrum Disorders. Oxid. Med. Cell. Longev. 2018, 2018, 3717215. [Google Scholar] [CrossRef]
- Wan, H.; Zhang, C.; Li, H.; Luan, S.; Liu, C. Association of maternal diabetes with autism spectrum disorders in offspring: A systemic review and meta-analysis. Medicine 2018, 97, e9438. [Google Scholar] [CrossRef]
- Rowland, J.; Wilson, C.A. The association between gestational diabetes and ASD and ADHD: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 5136. [Google Scholar] [CrossRef] [PubMed]
- Krakowiak, P.; Walker, C.K.; Bremer, A.A.; Baker, A.S.; Ozonoff, S.; Hansen, R.L.; Hertz-Picciotto, I. Maternal Metabolic Conditions and Risk for Autism and Other Neurodevelopmental Disorders. Pediatrics 2012, 129, e1121–e1128. [Google Scholar] [CrossRef]
- Ogden, C.L.; Fakhouri, T.H.; Carroll, M.D.; Hales, C.M.; Fryar, C.D.; Li, X.; Freedman, D.S. Prevalence of Obesity among Adults, by Household Income and Education-United States, 2011–2014. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 1369–1373. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.M.; Fryar, C.D.; Carroll, M.D.; Freedman, D.S.; Aoki, Y.; Ogden, C.L. Differences in Obesity Prevalence by Demographic Characteristics and Urbanization Level among Adults in the United States, 2013–2016. JAMA 2018, 319, 2419–2429. [Google Scholar] [CrossRef] [PubMed]
- Okosun, I.S.; Annor, F.B.; Seale, J.P.; Eriksen, M.P. Abdominal adiposity and family income-to-poverty ratio in American women. Obes. Res. Clin. Pract. 2014, 8, e201–e211. [Google Scholar] [CrossRef] [PubMed]
- Diehl, A.K.; Stern, M.P. Special health problems of Mexican-Americans: Obesity, gallbladder disease, diabetes mellitus, and cardiovascular disease. Adv. Intern. Med. 1989, 34, 73–96. [Google Scholar] [PubMed]
- Ogden, C.L.; Carroll, M.D.; Kit, B.K.; Flegal, K.M. Prevalence of Childhood and Adult Obesity in the United States, 2011–2012. JAMA J. Am. Med. Assoc. 2014, 311, 806–814. [Google Scholar] [CrossRef]
- Persson, E. What are the core ideas behind the Precautionary Principle? Sci. Total Environ. 2016, 557–558, 134–141. [Google Scholar] [CrossRef]
- Fowler, S.P.; Palmer, R.F. Early-Life Exposures to Diet Soda and Aspartame Are Associated with Autism in Males; dataset ed.; Dryad: Davis, CA, USA, 2020. [Google Scholar]


{kind=link}
| Characteristic | Boys (n = 257) | Girls (n = 99) | ||||||
|---|---|---|---|---|---|---|---|---|
| Cases | Cases | |||||||
| Controls (n = 54) | Any ASD (n = 203) | Autism (n = 140) | Non-Regressive Autism (n = 86) | Controls (n = 67) | Any ASD (n = 32) | Autism (n = 28) | Non-Regressive Autism (n = 19) | |
| Birth year of child (Mean (SD)) | 2001.3 (4.1) | 2001.4 (4.3) | 2001.5 (4.2) | 2001.5 (4.1) | 2002.2 (3.8) | 2001.8 (4.3) | 2001.9 (4.3) | 2001.4 (4.6) |
| Education of mother (n) | 54 | 202 | 139 | 86 | 67 | 32 | 28 | 19 |
| <High school (%) | 0 | 4 | 3 | 3 | 3 | 0 | 0 | 0 |
| High school graduate (%) | 13 | 5 | 8 | 9 | 4 | 9 | 7 | 5 |
| <4 yrs college (%) | 20 | 31 | 29 | 19 | 28 | 25 | 29 | 21 |
| ≥4 yrs college (%) | 67 | 60 | 60 | 69 | 64 | 66 | 64 | 74 |
| Family income (n) | 50 | 198 | 135 | 83 | 66 | 32 | 28 | 19 |
| <USD 25K (%) | 12 | 18 | 19 | 16 | 14 | 9 | 7 | 5 |
| USD 25K to <USD 50K (%) | 10 | 16 | 14 | 12 | 17 | 28 | 29 | 16 |
| USD 50K to <USD 100K (%) | 32 | 35 | 36 | 30 | 29 | 34 | 39 | 53 |
| USD 100K to <USD 200K (%) | 32 | 25 | 25 | 34 | 29 | 19 | 18 | 21 |
| ≥USD 200K (%) | 14 | 7 | 7 | 8 | 12 | 9 | 7 | 5 |
| Ethnicity of child (n) | 53 | 202 | 139 | 85 | 66 | 32 | 28 | 19 |
| Hispanic (%) | 25 | 16 | 15 | 13 | 32 | 9 | 7 | 5 |
| Non-Hispanic white (%) | 64 | 70 | 68 | 74 | 58 | 88 | 93 | 95 |
| Other/mixed (%) | 11 | 14 | 17 | 13 | 11 | 3 | 0 | 0 |
| Recruitment source (n) | 54 | 203 | 140 | 86 | 67 | 32 | 28 | 19 |
| IAN (%) | 69 | 74 | 73 | 72 | 60 | 87 | 89 | 89 |
| SA/STX (%) | 31 | 26 | 27 | 28 | 40 | 13 | 11 | 11 |
| Boys (n = 257) | Girls (n = 99) | |||||
|---|---|---|---|---|---|---|
| n | ≥1 Diet Soda/Day | ≥177 mg/Day of Aspartame | n | ≥1 Diet Soda/Day | ≥177 mg/Day of Aspartame | |
| % | % | % | % | |||
| Controls | 54 | 7.4 | 7.4 | 67 | 17.9 | 19.4 |
| ASD cases, excluding autism | 63 | 9.5 | 9.5 | 4 | 25.0 | 25.0 |
| Non-regressive ASD cases, excluding autism | 45 | 11.1 | 11.1 | 4 | 25.0 | 25.0 |
| All ASD cases, combined | 203 | 16.3 | 17.2 | 32 | 12.5 | 12.5 |
| Non-regressive ASD cases, combined | 131 | 18.3 | 19.1 * | 23 | 17.4 | 17.4 |
| Autism cases | 140 | 19.3 * | 20.7 * | 28 | 10.7 | 10.7 |
| Non-regressive autism cases | 86 | 22.1 * | 23.3 * | 19 | 15.8 | 15.8 |
| Condition | Boys | Girls | All Participants Combined c | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | OR | 95% CI | n | OR | 95% CI | n | OR | 95% CI | |
| ASD: all cases | |||||||||
| Unadjusted c | 257 | 2.4 | 0.8 to 7.2 | 99 | 0.7 | 0.2 to 2.2 | 356 | 1.4 | 0.7 to 2.8 |
| Adjusted d | 246 | 2.4 | 0.8 to 7.3 | 98 | 0.5 | 0.1 to 1.8 | 344 | 1.4 | 0.7 to 2.9 |
| Non-regressive ASD | |||||||||
| Unadjusted c | 185 | 2.8 | 0.9 to 8.5 | 90 | 1.0 | 0.3 to 3.4 | 275 | 1.8 | 0.8 to 3.8 |
| Adjusted d | 177 | 2.7 | 0.9 to 8.4 | 89 | 0.7 | 0.2 to 3.0 | 266 | 1.8 | 0.8 to 3.8 |
| Autism | |||||||||
| Unadjusted c | 194 | 3.0 | 0.99 to 9.0 | 95 | 0.6 | 0.1 to 2.1 | 289 | 1.5 | 0.7 to 3.1 |
| Adjusted d | 183 | 3.1 * | 1.02 to 9.7 | 94 | 0.3 | 0.1 to 1.3 | 277 | 1.5 | 0.7 to 3.2 |
| Non-regressive autism | |||||||||
| Unadjusted c | 140 | 3.5 * | 1.1 to 11.1 | 86 | 0.9 | 0.2 to 3.4 | 226 | 2.0 | 0.9 to 4.3 |
| Adjusted d | 132 | 3.5 * | 1.1 to 11.1 | 85 | 0.5 | 0.1 to 2.3 | 217 | 2.0 | 0.9 to 4.4 |
| Condition in Males | n | Daily Exposure | ORs | 95% CI |
|---|---|---|---|---|
| ASD: all cases | 246 | ≥1 serving/day of any NNS | 1.6 | 0.7 to 3.7 |
| ≥1 serving/day of aspartame | 1.9 | 0.8 to 5.0 | ||
| ≥177 mg/day of aspartame | 2.6 | 0.9 to 7.8 | ||
| Non-regressive ASD | 177 | ≥1 serving/day of any NNS | 1.7 | 0.7 to 4.1 |
| ≥1 serving/day of aspartame | 2.1 | 0.8 to 5.7 | ||
| ≥177 mg/day of aspartame | 2.9 | 0.9 to 8.8 | ||
| Autism | 183 | ≥1 serving/day of any NNS | 2.0 | 0.8 to 4.7 |
| ≥1 serving/day of aspartame | 2.5 | 0.95 to 6.5 | ||
| ≥177 mg/day of aspartame | 3.4 * | 1.1 to 10.4 | ||
| Non-regressive autism | 132 | ≥1 serving/day of any NNS | 2.1 | 0.8 to 5.2 |
| ≥1 serving/day of aspartame | 2.6 | 0.97 to 7.2 | ||
| ≥177 mg/day of aspartame | 3.7 * | 1.2 to 11.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fowler, S.P.; Gimeno Ruiz de Porras, D.; Swartz, M.D.; Stigler Granados, P.; Heilbrun, L.P.; Palmer, R.F. Daily Early-Life Exposures to Diet Soda and Aspartame Are Associated with Autism in Males: A Case-Control Study. Nutrients 2023, 15, 3772. https://doi.org/10.3390/nu15173772
Fowler SP, Gimeno Ruiz de Porras D, Swartz MD, Stigler Granados P, Heilbrun LP, Palmer RF. Daily Early-Life Exposures to Diet Soda and Aspartame Are Associated with Autism in Males: A Case-Control Study. Nutrients. 2023; 15(17):3772. https://doi.org/10.3390/nu15173772
Chicago/Turabian StyleFowler, Sharon Parten, David Gimeno Ruiz de Porras, Michael D. Swartz, Paula Stigler Granados, Lynne Parsons Heilbrun, and Raymond F. Palmer. 2023. "Daily Early-Life Exposures to Diet Soda and Aspartame Are Associated with Autism in Males: A Case-Control Study" Nutrients 15, no. 17: 3772. https://doi.org/10.3390/nu15173772
APA StyleFowler, S. P., Gimeno Ruiz de Porras, D., Swartz, M. D., Stigler Granados, P., Heilbrun, L. P., & Palmer, R. F. (2023). Daily Early-Life Exposures to Diet Soda and Aspartame Are Associated with Autism in Males: A Case-Control Study. Nutrients, 15(17), 3772. https://doi.org/10.3390/nu15173772