Revisiting One-Carbon Metabolites in Human Breast Milk: Focus on S-Adenosylmethionine

 ,
, 
Abstract
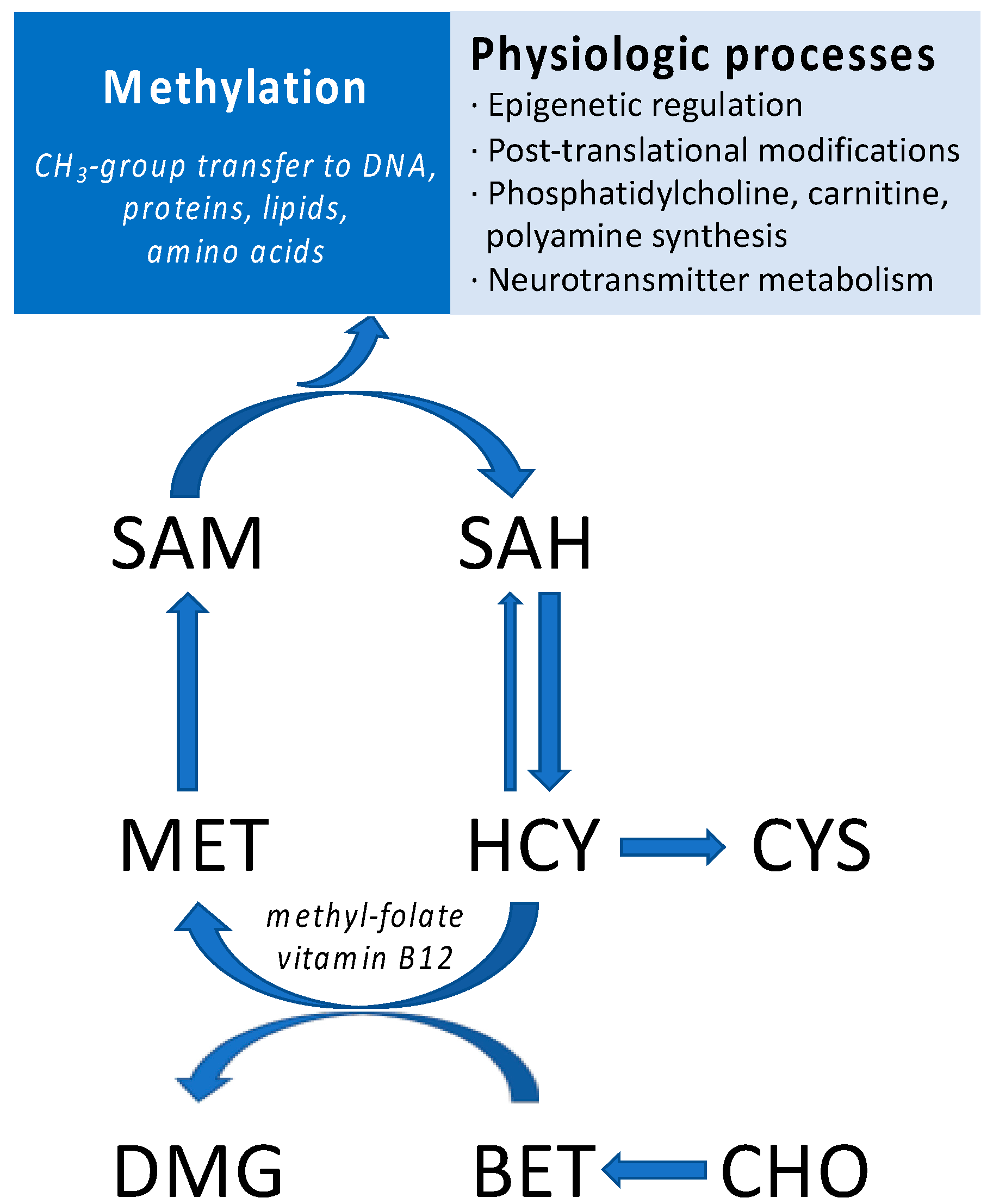
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Metabolite Analysis
2.3. Statistical Analysis
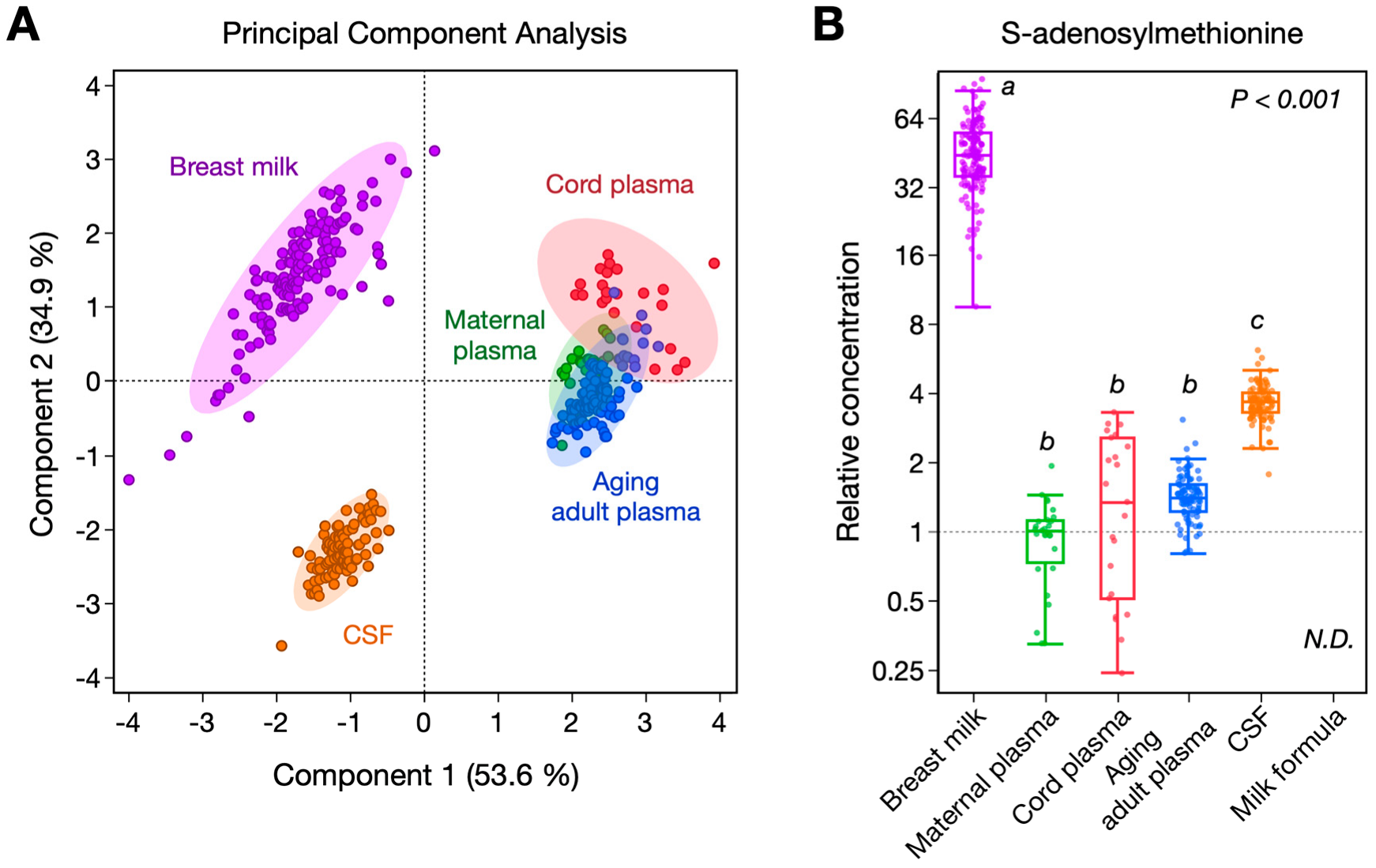
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Association of Pediatrics. Breastfeeding and the use of human milk. Pediatrics 2012, 129, e827–e841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuebe, A.M.; Schwarz, E.B. The risks and benefits of infant feeding practices for women and their children. J. Perinatol. 2010, 30, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Breastfeeding. Available online: https://www.who.int/health-topics/breastfeeding (accessed on 1 December 2022).
- Kalhan, S.C. One carbon metabolism in pregnancy: Impact on maternal, fetal and neonatal health. Mol. Cell Endocrinol. 2016, 435, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBreairty, L.E.; Bertolo, R.F. The dynamics of methionine supply and demand during early development. Appl. Physiol. Nutr. Metab. 2016, 41, 581–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeisel, S.H. Choline: Critical Role During Fetal Development and Dietary Requirements in Adults. Ann. Rev. Nutr. 2006, 26, 229–250. [Google Scholar] [CrossRef] [Green Version]
- Fischer, L.M.; da Costa, K.A.; Galanko, J.; Sha, W.; Stephenson, B.; Vick, J.; Zeisel, S.H. Choline intake and genetic polymorphisms influence choline metabolite concentrations in human breast milk and plasma. Am. J. Clin. Nutr. 2010, 92, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Mato, J.M.; Martínez-Chantar, M.L.; Lu, S.C. Methionine Metabolism and Liver Disease. Ann. Rev. Nutr. 2008, 28, 273–293. [Google Scholar] [CrossRef] [Green Version]
- Ullah, H.; Khan, A.; Rengasamy, K.R.R.; Di Minno, A.; Sacchi, R.; Daglia, M. The Efficacy of S-Adenosyl Methionine and Probiotic Supplementation on Depression: A Synergistic Approach. Nutrients 2022, 14, 2751. [Google Scholar] [CrossRef]
- Sharma, A.; Gerbarg, P.; Bottiglieri, T.; Massoumi, L.; Carpenter, L.L.; Lavretsky, H.; Muskin, P.R.; Brown, R.P.; Mischoulon, D. S-Adenosylmethionine (SAMe) for Neuropsychiatric Disorders: A Clinician-Oriented Review of Research. J. Clin. Psychiatry 2017, 78, e656–e667. [Google Scholar] [CrossRef] [Green Version]
- Pascale, R.M.; Simile, M.M.; Calvisi, D.F.; Feo, C.F.; Feo, F. S-Adenosylmethionine: From the Discovery of Its Inhibition of Tumorigenesis to Its Use as a Therapeutic Agent. Cells 2022, 11, 409. [Google Scholar] [CrossRef]
- Noureddin, M.; Mato, J.M.; Lu, S.C. Nonalcoholic fatty liver disease: Update on pathogenesis, diagnosis, treatment and the role of S-adenosylmethionine. Exp. Biol. Med. 2015, 240, 809–820. [Google Scholar] [CrossRef]
- Rutjes, A.W.; Nüesch, E.; Reichenbach, S.; Jüni, P. S-Adenosylmethionine for osteoarthritis of the knee or hip. Cochrane Database Syst. Rev. 2009, 2009, CD007321. [Google Scholar] [CrossRef] [Green Version]
- Soeken, K.L.; Lee, W.L.; Bausell, R.B.; Agelli, M.; Berman, B.M. Safety and efficacy of S-adenosylmethionine (SAMe) for osteoarthritis. J. Fam. Pract. 2002, 51, 425–430. [Google Scholar]
- Ribo, S.; Sánchez-Infantes, D.; Martinez-Guino, L.; García-Mantrana, I.; Ramon-Krauel, M.; Tondo, M.; Arning, E.; Nofrarías, M.; Osorio-Conles, Ó.; Fernández-Pérez, A.; et al. Increasing breast milk betaine modulates Akkermansia abundance in mammalian neonates and improves long-term metabolic health. Sci. Transl. Med. 2021, 13, eabb0322. [Google Scholar] [CrossRef]
- Rooney, M.; Bottiglieri, T.; Wasek-Patterson, B.; McMahon, A.; Hughes, C.F.; McCann, A.; Horigan, G.; Strain, J.J.; McNulty, H.; Ward, M. Impact of the MTHFR C677T polymorphism on one-carbon metabolites: Evidence from a randomised trial of riboflavin supplementation. Biochimie 2020, 173, 91–99. [Google Scholar] [CrossRef]
- Inoue-Choi, M.; Nelson, H.H.; Robien, K.; Arning, E.; Bottiglieri, T.; Koh, W.P.; Yuan, J.M. One-carbon metabolism nutrient status and plasma S-adenosylmethionine concentrations in middle-aged and older Chinese in Singapore. Int. J. Mol. Epidemiol. Genet. 2012, 3, 160–173. [Google Scholar]
- García-Mantrana, I.; Alcántara, C.; Selma-Royo, M.; Boix-Amorós, A.; Dzidic, M.; Gimeno-Alcañiz, J.; Úbeda-Sansano, I.; Sorribes-Monrabal, I.; Escuriet, R.; Gil-Raga, F.; et al. MAMI: A birth cohort focused on maternal-infant microbiota during early life. BMC Pediatr. 2019, 19, 140. [Google Scholar] [CrossRef] [Green Version]
- Ubbink, J.B.; Hayward Vermaak, W.J.; Bissbort, S. Rapid high-performance liquid chromatographic assay for total homocysteine levels in human serum. J. Chromatogr. 1991, 565, 441–446. [Google Scholar] [CrossRef]
- Jeong, H.; Suh, Y.; Chung, Y.-J. Choline and Betaine Concentrations in Breast Milk of Korean Lactating Women and the Choline and Betaine Intakes of Their Infants. Korean J. Nutr. 2010, 43, 588–596. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, A.; Ono, H.; Mizoguchi, N.; Sakura, N. Betaine and homocysteine concentrations in infant formulae and breast milk. Pediatr. Int. 2001, 43, 637–640. [Google Scholar] [CrossRef]
- Bardanzellu, F.; Puddu, M.; Peroni, D.G.; Fanos, V. The Human Breast Milk Metabolome in Overweight and Obese Mothers. Front. Immunol. 2020, 11, 1533. [Google Scholar] [CrossRef] [PubMed]
- Hampel, D.; Shahab-Ferdows, S.; Hossain, M.; Islam, M.M.; Ahmed, T.; Allen, L.H. Validation and Application of Biocrates AbsoluteIDQ® p180 Targeted Metabolomics Kit Using Human Milk. Nutrients 2019, 11, 1733. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.S.; Lewis, A.N.; Gong, Q.; Sollome, J.J.; DeWitt, O.N.; Williams, R.D.; Good, M. Untargeted Metabolomic Analysis of Human Milk from Mothers of Preterm Infants. Nutrients 2021, 13, 3604. [Google Scholar] [CrossRef] [PubMed]
- Isganaitis, E.; Venditti, S.; Matthews, T.J.; Lerin, C.; Demerath, E.W.; Fields, D.A. Maternal obesity and the human milk metabolome: Associations with infant body composition and postnatal weight gain. Am. J. Clin. Nutr. 2019, 110, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Bottiglieri, T. S-Adenosyl-L-methionine (SAMe): From the bench to the bedside—Molecular basis of a pleiotrophic molecule. Am. J. Clin. Nutr. 2002, 76, 1151S–1157S. [Google Scholar] [CrossRef] [Green Version]
- Cunliffe, V.T. Experience-sensitive epigenetic mechanisms, developmental plasticity, and the biological embedding of chronic disease risk. Wiley Interdiscip. Rev. Syst. Biol. Med. 2015, 7, 53–71. [Google Scholar] [CrossRef]
- Hao, G.; Youssef, N.A.; Davis, C.L.; Su, S. The role of DNA methylation in the association between childhood adversity and cardiometabolic disease. Int. J. Cardiol. 2018, 255, 168–174. [Google Scholar] [CrossRef]
- Wikenius, E.; Moe, V.; Smith, L.; Heiervang, E.R.; Berglund, A. DNA methylation changes in infants between 6 and 52 weeks. Sci. Rep. 2019, 9, 17587. [Google Scholar] [CrossRef] [Green Version]
- Mato, J.M.; Martínez-Chantar, M.L.; Lu, S.C. S-adenosylmethionine metabolism and liver disease. Ann. Hepatol. 2013, 12, 183–189. [Google Scholar] [CrossRef]
- Yang, J.; He, Y.; Du, Y.X.; Tang, L.L.; Wang, G.J.; Fawcett, J.P. Pharmacokinetic properties of S-adenosylmethionine after oral and intravenous administration of its tosylate disulfate salt: A multiple-dose, open-label, parallel-group study in healthy Chinese volunteers. Clin. Ther. 2009, 31, 311–320. [Google Scholar] [CrossRef]
- Freeman, M.P. Complementary and alternative medicine for perinatal depression. J. Affect. Disord. 2009, 112, 1–10. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Breast Milk (n = 142) | Maternal Plasma (n = 28) | Cord Plasma (n = 23) | Adult Plasma (n = 91) | Cerebrospinal Fluid (n = 92) | Milk Formula (n = 11) | p Value | |
|---|---|---|---|---|---|---|---|
| CHO, µmol/L | 123.5 (92.8) a | 11.8 (4.2) b | 56.8 (36.0) a,b | 7.1 (2.2) b | 2.7 (1.0) c | 1510 (208) d | <0.0001 |
| BET, µmol/L | 3.6 (3.2) a | 12.2 (5.5) b | 30.9 (16.7) b,c | 35.9 (14.8) c | 1.9 (0.6) d | 16.0 (15.7) a,b | <0.0001 |
| MET, µmol/L | 4.2 (2.1) a | 21.4 (8.6) b,c | 31.0 (5.7) b | 21.1 (4.5) b,c | 3.2 (1.0) d | 272 (358) c | <0.0001 |
| SAM, nmol/L | 1830 (805) a | 42 (16) b,c | 56 (86) b,c | 59 (16) b | 154 (29) d | <0.1 c | <0.0001 |
| SAH, nmol/L | 263 (192) a | 41 (14) b | 43 (39) b,c | 26 (8) b,c | 10 (6) d | 18.6 (27.7) c,d | <0.0001 |
| tHCY, µmol/L * | 0.27 (0.15) a | 7.38 (2.60) b | 7.05 (2.03) b | 5.70 (2.90) b | 0.09 (0.03) c | N.A. | <0.0001 |
| CYS, µmol/L | 91 (91) a | 199 (107) b | 334 (187) b | 105 (61) a | 43 (20) c | 20.9 (11.3) c | <0.0001 |
| CHO | BET | MET | SAM | SAH | CYS | tHCY | |
|---|---|---|---|---|---|---|---|
| CHO | 1 | ||||||
| BET | 0.58 (<0.001) | 1 | |||||
| MET | 0.43 (<0.001) | 0.25 (0.003) | 1 | ||||
| SAM | 0.01 (0.904) | −0.18 (0.035) | 0.02 (0.851) | 1 | |||
| SAH | 0.25 (0.003) | 0.45 (<0.001) | 0.09 (0.262) | −0.20 (0.015) | 1 | ||
| CYS | 0.46 (<0.001) | 0.45 (<0.001) | 0.32 (<0.001) | 0.32 (<0.001) | 0.42 (<0.001) | 1 | |
| tHCY * | 0.30 (<0.001) | 0.22 (0.009) | 0.25 (0.003) | 0.08 (0.376) | 0.26 (0.002) | 0.41 (<0.001) | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lerin, C.; Collado, M.C.; Isganaitis, E.; Arning, E.; Wasek, B.; Demerath, E.W.; Fields, D.A.; Bottiglieri, T. Revisiting One-Carbon Metabolites in Human Breast Milk: Focus on S-Adenosylmethionine. Nutrients 2023, 15, 282. https://doi.org/10.3390/nu15020282
Lerin C, Collado MC, Isganaitis E, Arning E, Wasek B, Demerath EW, Fields DA, Bottiglieri T. Revisiting One-Carbon Metabolites in Human Breast Milk: Focus on S-Adenosylmethionine. Nutrients. 2023; 15(2):282. https://doi.org/10.3390/nu15020282
Chicago/Turabian StyleLerin, Carles, María Carmen Collado, Elvira Isganaitis, Erland Arning, Brandi Wasek, Ellen W. Demerath, David A. Fields, and Teodoro Bottiglieri. 2023. "Revisiting One-Carbon Metabolites in Human Breast Milk: Focus on S-Adenosylmethionine" Nutrients 15, no. 2: 282. https://doi.org/10.3390/nu15020282
APA StyleLerin, C., Collado, M. C., Isganaitis, E., Arning, E., Wasek, B., Demerath, E. W., Fields, D. A., & Bottiglieri, T. (2023). Revisiting One-Carbon Metabolites in Human Breast Milk: Focus on S-Adenosylmethionine. Nutrients, 15(2), 282. https://doi.org/10.3390/nu15020282