
The Effect of an Essential Oil Blend on Growth Performance, Intestinal Health, and Microbiota in Early-Weaned Piglets

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Design
2.2. Sample Collection
2.3. Growth Performance, Fecal Score, and Organ Index
2.4. Determination of Hematology and Plasma Biochemical Parameters
2.5. Phenotype of the T-Lymphocytes
2.6. Intestinal Morphology
2.7. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.8. Western Blotting Analysis
2.9. Determination of Volatile Fatty Acids (VFA)
2.10. Colonic Digesta Microbiota Analysis
2.11. Statistical Analysis
3. Results
3.1. Effect of EO on Piglet Growth Performance
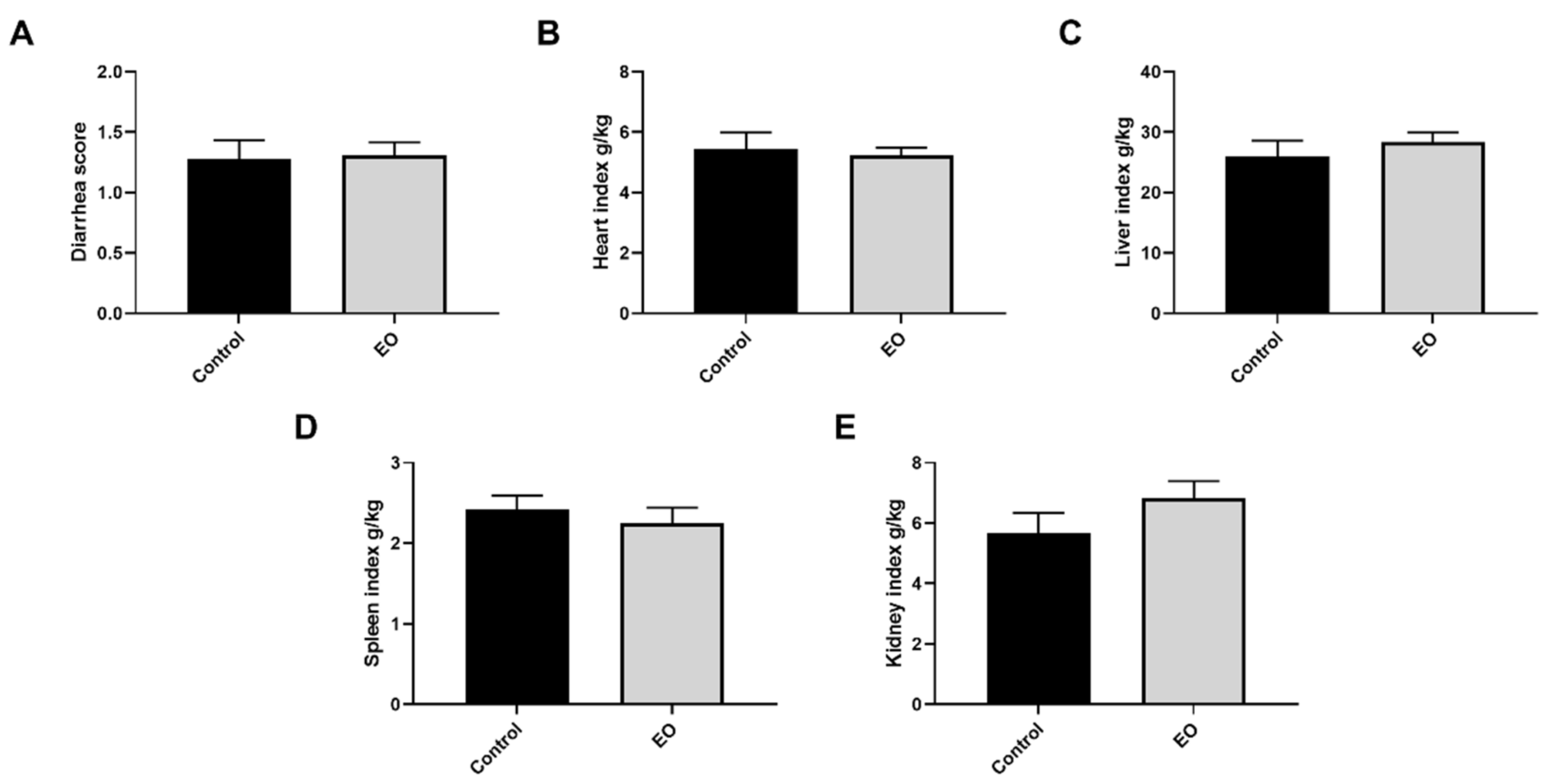
3.2. Effect of EO on Piglet Diarrhea Score and Organ Index
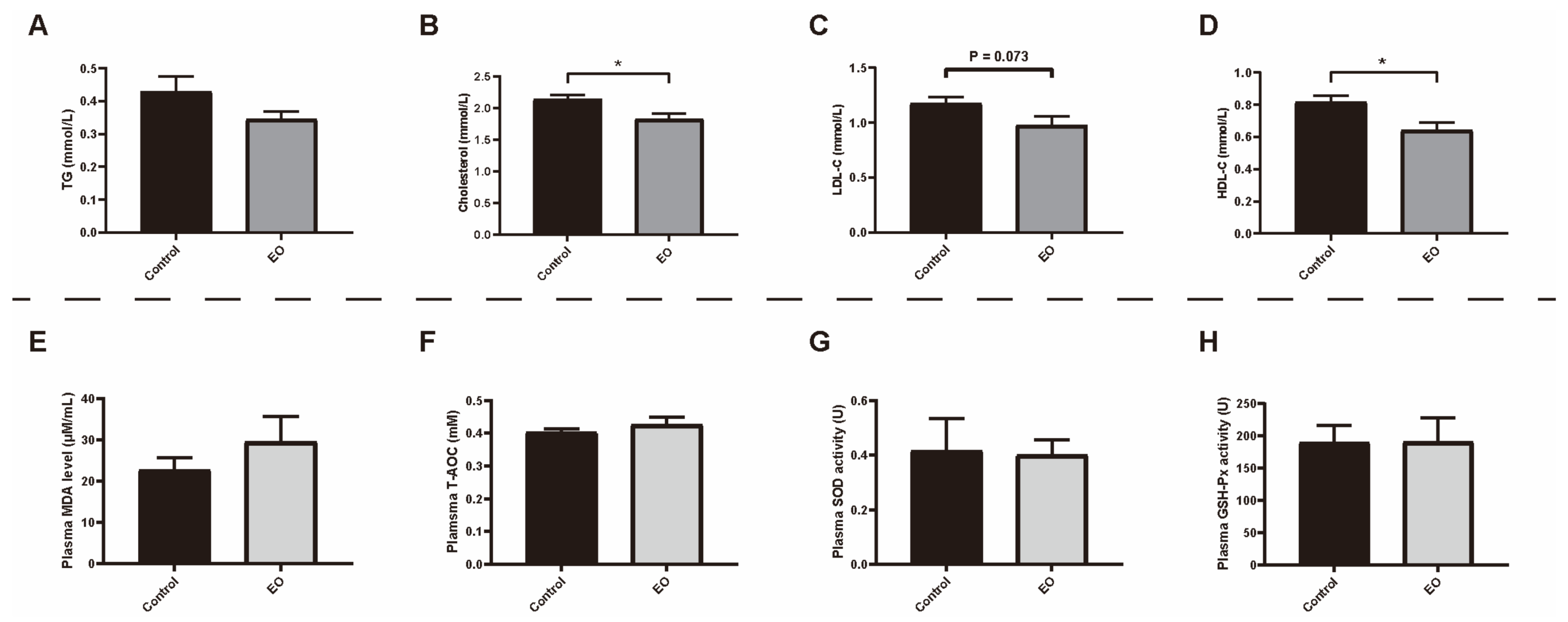
3.3. Effect of EO on Piglet Hematology, Plasma Biochemical Index, and Antioxidant Capacity
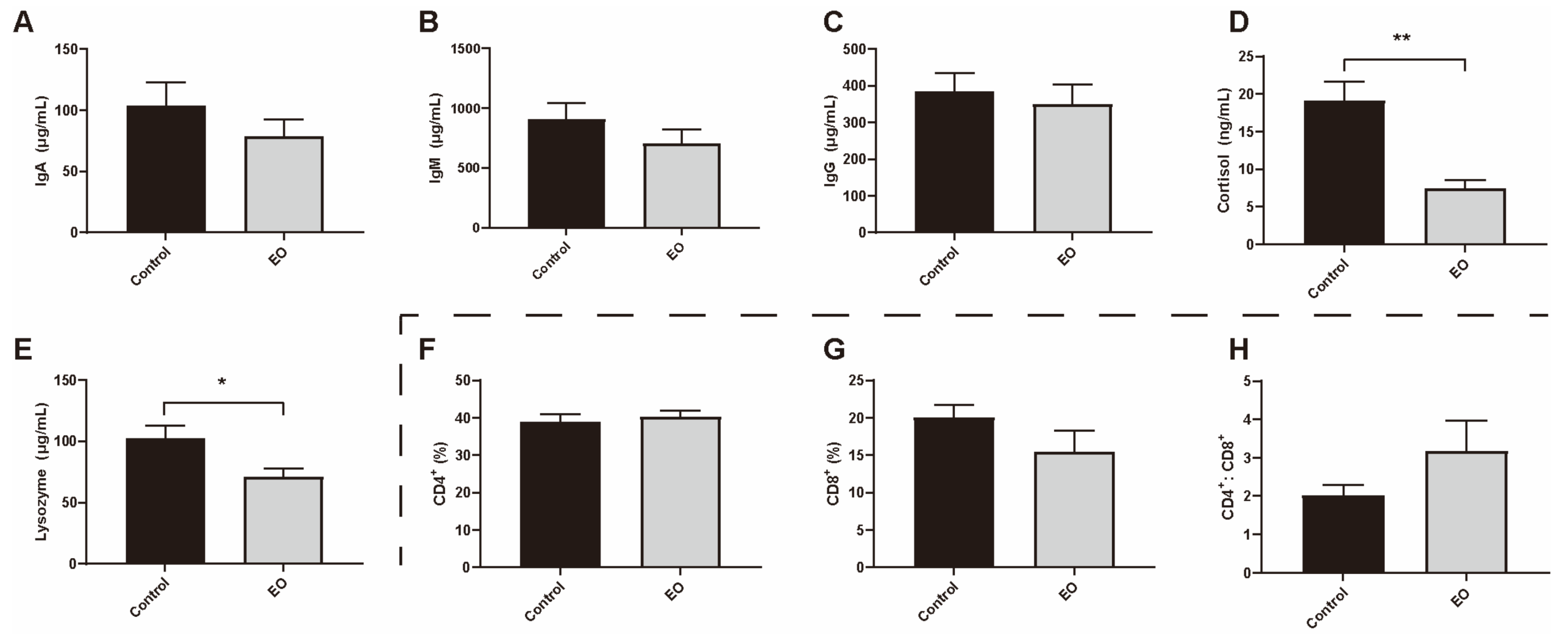
3.4. Effect of EO on Piglet Immunity
3.5. Effect of EO on Piglet Intestinal Epithelial Morphological Structure
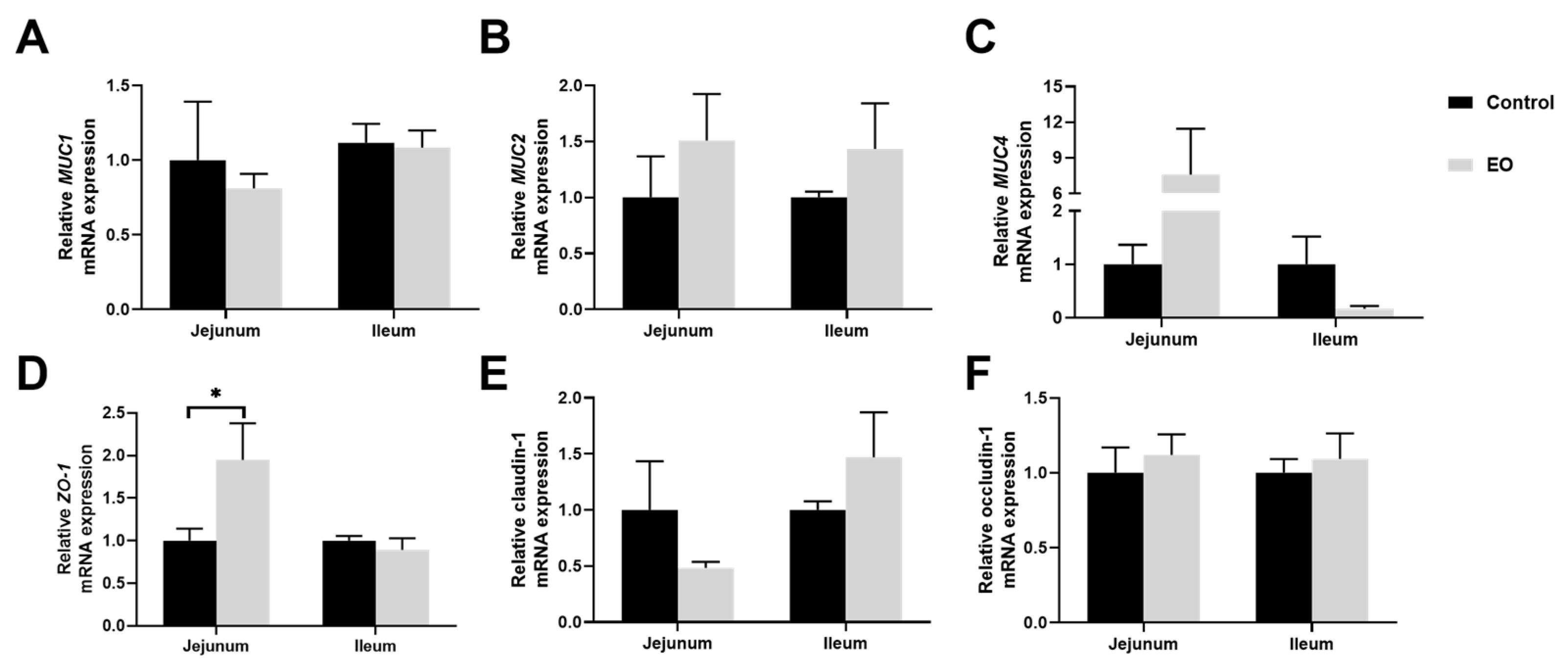
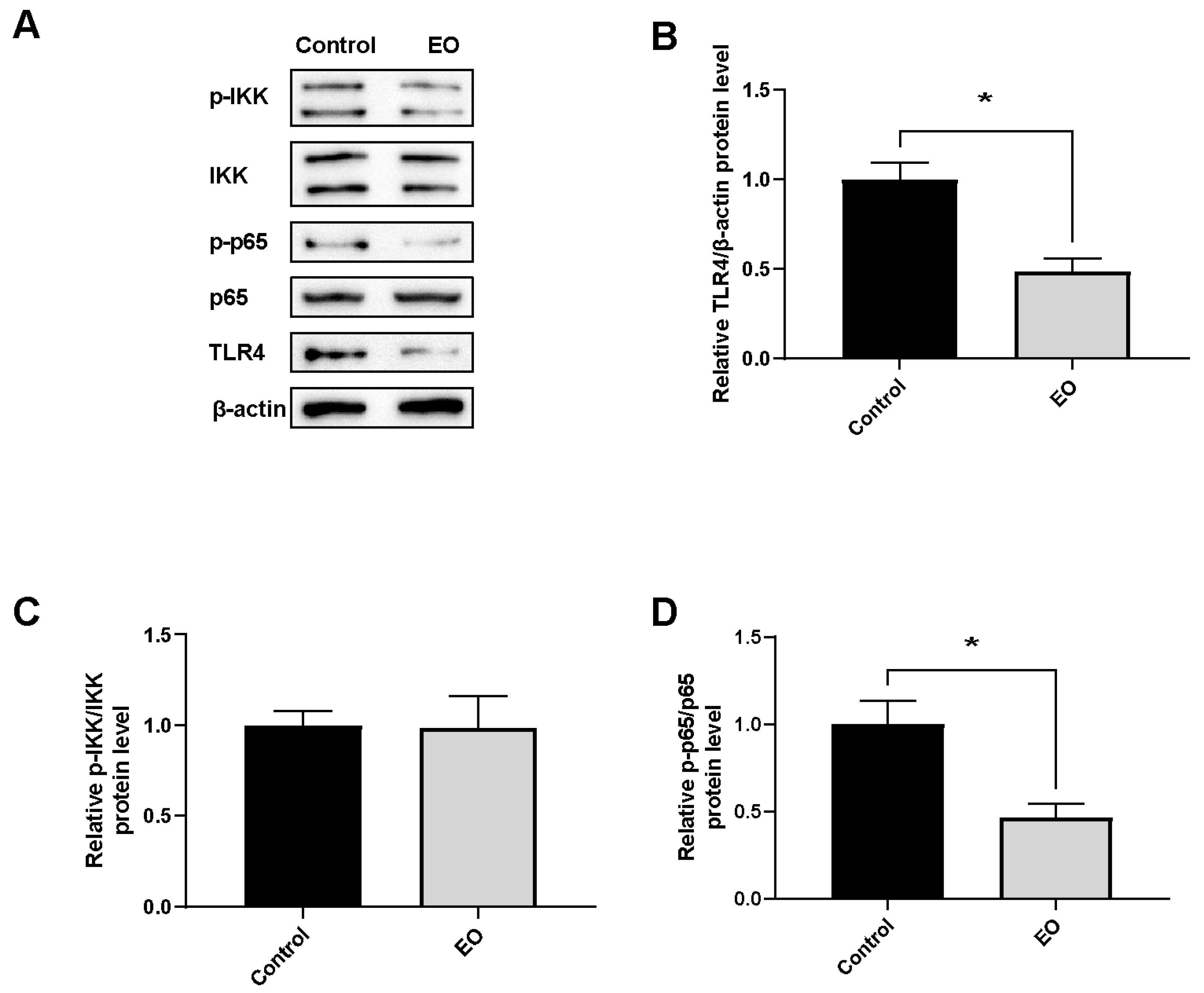
3.6. Effect of EO on Piglet Intestinal Barrier Function
3.7. Effect of EO on Piglet Ileum Immune Function
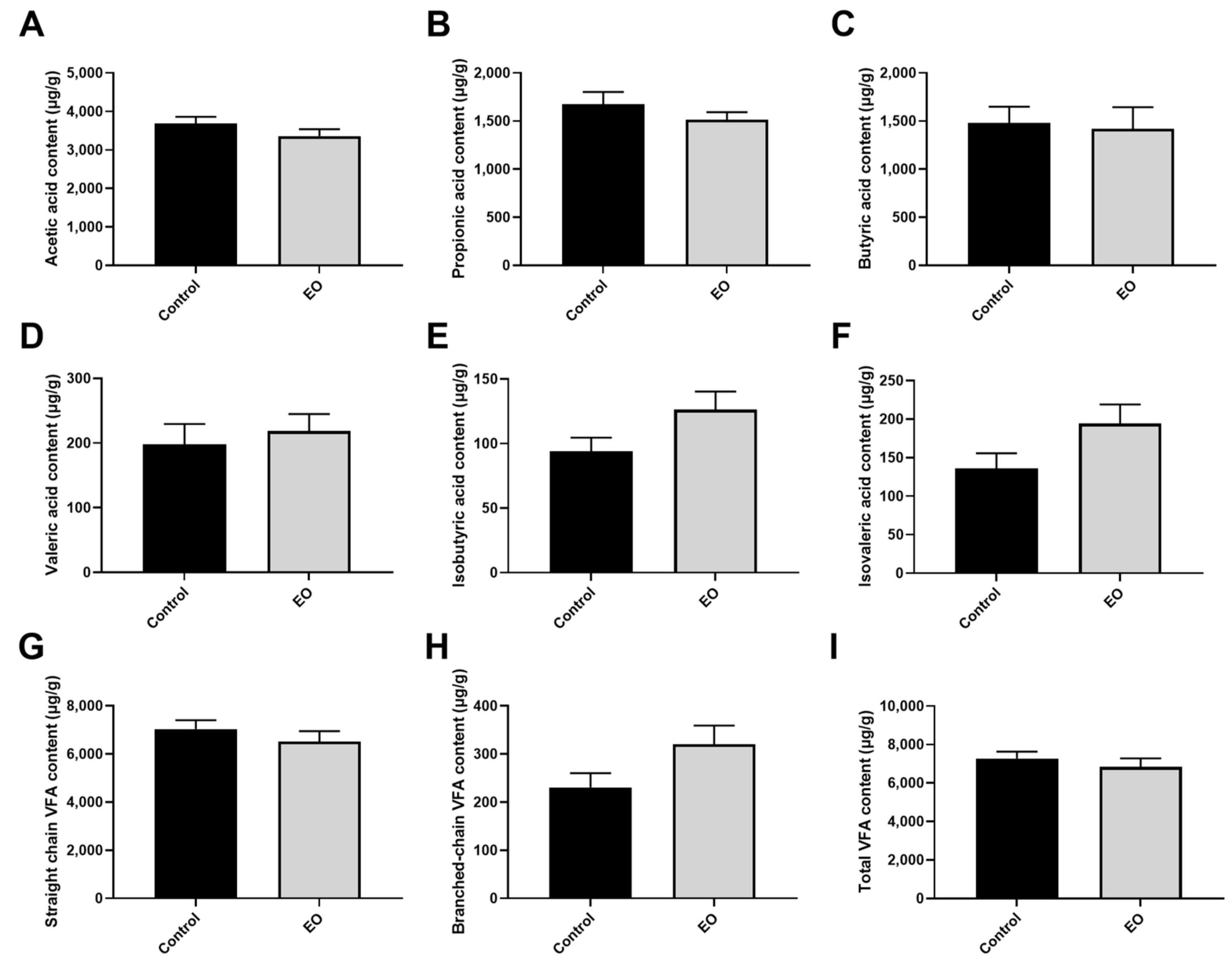
3.8. Effect of EO on VFA Concentrations and Microbiome in Piglet Colonic Content
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lallès, J.P.; Boudry, G.; Favier, C.; Le Floc’h, N.; Luron, I.; Montagne, L.; Oswald, I.P.; Pié, S.; Piel, C.; Sève, B. Gut function and dysfunction in young pigs: Physiology. Anim. Res. 2004, 53, 301–316. [Google Scholar] [CrossRef] [Green Version]
- Lewis, N.J. Transport of early weaned piglets. Appl. Anim. Behav. Sci. 2008, 110, 128–135. [Google Scholar] [CrossRef]
- Parratt, C.A.; Chapman, K.J.; Turner, C.; Jones, P.H.; Mendl, M.T.; Miller, B.G. The fighting behaviour of piglets mixed before and after weaning in the presence or absence of a sow. Appl. Anim. Behav. Sci. 2006, 101, 54–67. [Google Scholar] [CrossRef]
- Sutherland, M.A.; Backus, B.L.; McGlone, J.J. Effects of transport at weaning on the behavior, physiology and performance of pigs. Animals 2014, 4, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Zhao, K.; Chen, X.; Xu, J. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J. Anim. Sci. 2012, 90, 2581–2589. [Google Scholar] [CrossRef]
- Wijtten, P.J.; van der Meulen, J.; Verstegen, M.W. Intestinal barrier function and absorption in pigs after weaning: A review. Br. J. Nutr. 2011, 105, 967–981. [Google Scholar] [CrossRef]
- Yin, J.; Wu, M.M.; Xiao, H.; Ren, W.K.; Duan, J.L.; Yang, G.; Li, T.J.; Yin, Y.L. Development of an antioxidant system after early weaning in piglets2. J. Anim. Sci. 2014, 92, 612–619. [Google Scholar] [CrossRef]
- Pohl, C.S.; Medland, J.E.; Mackey, E.; Edwards, L.L.; Bagley, K.D.; DeWilde, M.P.; Williams, K.J.; Moeser, A.J. Early weaning stress induces chronic functional diarrhea, intestinal barrier defects, and increased mast cell activity in a porcine model of early life adversity. Neurogastroenterol. Motil. 2017, 29, e13118. [Google Scholar] [CrossRef] [Green Version]
- Novais, A.K.; Deschene, K.; Martel-Kennes, Y.; Roy, C.; Laforest, J.P.; Lessard, M.; Matte, J.J.; Lapointe, J. Weaning differentially affects mitochondrial function, oxidative stress, inflammation and apoptosis in normal and low birth weight piglets. PLoS ONE 2021, 16, e0247188. [Google Scholar] [CrossRef]
- Brenes, A.; Roura, E. Essential oils in poultry nutrition: Main effects and modes of action. Anim. Feed Sci. Technol. 2010, 158, 1–14. [Google Scholar] [CrossRef]
- Zeng, Z.; Zhang, S.; Wang, H.; Piao, X. Essential oil and aromatic plants as feed additives in non-ruminant nutrition: A review. J. Anim. Sci. Biotechnol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Chen, S.; Wang, L. Purification and identification of carvacrol from the root of Stellera chamaejasme and research on its insecticidal activity. Nat. Prod. Res. 2011, 25, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Mancini, E.; Senatore, F.; Del Monte, D.; De Martino, L.; Grulova, D.; Scognamiglio, M.; Snoussi, M.; De Feo, V. Studies on chemical composition, antimicrobial and antioxidant activities of five Thymus vulgaris L. essential oils. Molecules 2015, 20, 12016–12028. [Google Scholar] [CrossRef] [Green Version]
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential oils as alternatives to antibiotics in swine production. Anim. Nutr. 2018, 4, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, A.; Przychodna, M.; Sopata, S.; Bodalska, A.; Fecka, I. Thymol and Thyme Essential Oil—New Insights into Selected Therapeutic Applications. Molecules 2020, 25, 4125. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Mishra, A.P.; Shukla, I.; Sharifi-Rad, M.; Contreras, M.d.M.; Segura-Carretero, A.; Fathi, H.; Nasrabadi, N.N.; Kobarfard, F.; Sharifi-Rad, J. Thymol, thyme, and other plant sources: Health and potential uses. Phytother. Res. 2018, 32, 1688–1706. [Google Scholar] [CrossRef]
- Suntres, Z.E.; Coccimiglio, J.; Alipour, M. The Bioactivity and Toxicological Actions of Carvacrol. Crit. Rev. Food Sci. Nutr. 2015, 55, 304–318. [Google Scholar] [CrossRef] [PubMed]
- Hajinejad, M.; Ghaddaripouri, M.; Dabzadeh, M.; Forouzanfar, F.; Sahab-Negah, S. Natural Cinnamaldehyde and Its Derivatives Ameliorate Neuroinflammatory Pathways in Neurodegenerative Diseases. BioMed Res. Int. 2020, 2020, 1034325. [Google Scholar] [CrossRef]
- Li, P.; Piao, X.; Ru, Y.; Han, X.; Xue, L.; Zhang, H. Effects of Adding Essential Oil to the Diet of Weaned Pigs on Performance, Nutrient Utilization, Immune Response and Intestinal Health. Asian-Aust. J. Anim. Sci. 2012, 25, 1617–1626. [Google Scholar] [CrossRef]
- Zeng, Z.; Xu, X.; Zhang, Q.; Li, P.; Zhao, P.; Li, Q.; Liu, J.; Piao, X. Effects of essential oil supplementation of a low-energy diet on performance, intestinal morphology and microflora, immune properties and antioxidant activities in weaned pigs. Anim. Sci. J. 2015, 86, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Piao, X. Essential oil blend could decrease diarrhea prevalence by improving antioxidative capability for weaned pigs. Animals 2019, 9, 847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.T.; Liu, L.; Long, S.F.; Pan, L.; Piao, X.S. Effect of organic acids and essential oils on performance, intestinal health and digestive enzyme activities of weaned pigs. Anim. Feed Sci. Technol. 2018, 235, 110–119. [Google Scholar] [CrossRef]
- Mo, K.B.; Li, J.; Liu, F.F.; Xu, Y.; Huang, X.H.; Ni, H.J. Superiority of Microencapsulated Essential Oils Compared with Common Essential Oils and Antibiotics: Effects on the Intestinal Health and Gut Microbiota of Weaning Piglet. Front Nutr. 2022, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Swine; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Wang, Q.; Xiong, X.; Li, J.; Tu, Q.; Yang, H.; Yin, Y. Energy metabolism in the intestinal crypt epithelial cells of piglets during the suckling period. Sci. Rep. 2018, 8, 12948. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Song, Y.; Yu, B.; He, J.; Zheng, P.; Mao, X.; Huang, Z.; Luo, Y.; Luo, J.; Yan, H.; et al. Tannic acid prevents post-weaning diarrhea by improving intestinal barrier integrity and function in weaned piglets. J. Anim. Sci. Biotechnol. 2020, 11, 87. [Google Scholar] [CrossRef]
- Li, J.; Zhong, Y.; Li, H.; Zhang, N.; Ma, W.; Cheng, G.; Liu, F.; Liu, F.; Xu, J. Enhancement of Astragalus polysaccharide on the immune responses in pigs inoculated with foot-and-mouth disease virus vaccine. Int. J. Biol. Macromol. 2011, 49, 362–368. [Google Scholar] [CrossRef]
- Deng, Q.; Shao, Y.; Wang, Q.; Li, J.; Li, Y.; Ding, X.; Huang, P.; Yin, J.; Yang, H.; Yin, Y. Effects and interaction of dietary electrolyte balance and citric acid on the intestinal function of weaned piglets. J. Anim. Sci. 2020, 98, skaa106. [Google Scholar] [CrossRef]
- Wang, J.; Zeng, L.; Tan, B.; Li, G.; Huang, B.; Xiong, X.; Li, F.; Kong, X.; Liu, G.; Yin, Y. Developmental changes in intercellular junctions and Kv channels in the intestine of piglets during the suckling and post-weaning periods. J. Anim. Sci. Biotechnol. 2016, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Z.; Liu, S.; Zhou, Y.; Mi, S.; Liu, G.; Wu, X.; Wu, X.; Yao, K.; Assaad, H.; Deng, Z.; et al. Chlorogenic acid decreases intestinal permeability and increases expression of intestinal tight junction proteins in weaned rats challenged with LPS. PLoS ONE 2014, 9, e97815. [Google Scholar] [CrossRef]
- Qi, M.; Tan, B.; Wang, J.; Li, J.; Liao, S.; Yan, J.; Liu, Y.; Yin, Y. Small intestinal transcriptome analysis revealed changes of genes involved in nutrition metabolism and immune responses in growth retardation piglets1. J. Anim. Sci. 2019, 97, 3795–3808. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Wu, M.M.; Tan, B.E.; Yin, Y.L.; Li, T.J.; Xiao, D.F.; Li, L. Effects of composite antimicrobial peptides in weanling piglets challenged with deoxynivalenol: II. Intestinal morphology and function1. J. Anim. Sci. 2013, 91, 4750–4756. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Tan, B.; Wang, J.; Liao, S.; Li, J.; Cui, Z.; Shao, Y.; Ji, P.; Yin, Y. Postnatal growth retardation is associated with deteriorated intestinal mucosal barrier function using a porcine model. J. Cell Physiol. 2021, 236, 2631–2648. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.F.; Blisklager, A.T.; Moeser, A.J. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Gastroint. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surai, P.; Fisinin, V. Antioxidant-prooxidant balance in the intestine: Applications in chick placement and pig weaning. J. Vet. Sci. Med. 2015, 3, 66–84. [Google Scholar]
- Li, S.Y.; Ru, Y.J.; Liu, M.; Xu, B.; Péron, A.; Shi, X.G. The effect of essential oils on performance, immunity and gut microbial population in weaner pigs. Livest. Sci. 2012, 145, 119–123. [Google Scholar] [CrossRef]
- Ogawa, K.; Honda, M.; Tanigawa, A.; Hatase, A.; Ito, A.; Higa, Y.; Morinaga, O. Appetite-enhancing effects of inhaling cinnamon, clove, and fennel essential oils containing phenylpropanoid analogues. J. Nat. Med. 2020, 74, 710–721. [Google Scholar] [CrossRef]
- Pluske, J.R. Psychological and Nutritional Stress in Pigs at Weaning: Production Parameters, the Stress Response, and Histology and Biochemistry of the Small Intestine; University of Western Australia: Crawley, Australia, 1993. [Google Scholar]
- Van Beers-Schreurs, H.M.G.; Nabuurs, M.J.A.; Vellenga, L.; Valk HJK-vd Wensing, T.; Breukink, H.J. Weaning and the Weanling Diet Influence the Villous Height and Crypt Depth in the Small Intestine of Pigs and Alter the Concentrations of Short-Chain Fatty Acids in the Large Intestine and Blood. J. Nutr. 1998, 128, 947–953. [Google Scholar] [CrossRef] [Green Version]
- Van der Peet-Schwering, C.; Jansman, A.; Smidt, H.; Yoon, I. Effects of yeast culture on performance, gut integrity, and blood cell composition of weanling pigs. J. Anim. Sci. 2007, 85, 3099–3109. [Google Scholar] [CrossRef] [PubMed]
- Michiels, J.; Missotten, J.; Van Hoorick, A.; Ovyn, A.; Fremaut, D.; De Smet, S.; Dierick, N. Effects of dose and formulation of carvacrol and thymol on bacteria and some functional traits of the gut in piglets after weaning. Arch. Anim. Nutr. 2010, 64, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Fu, C.; Yan, M.; Xie, H.; Li, S.; Yu, Q.; He, S.; He, J. Resveratrol modulates intestinal morphology and HSP70/90, NF-κB and EGF expression in the jejunal mucosa of black-boned chickens on exposure to circular heat stress. Food Funct. 2016, 7, 1329–1338. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, X.H.; Yang, L.; Chen, X.Y.; Jiang, R.S.; Jin, S.H.; Geng, Z.Y. Resveratrol alleviates heat stress-induced impairment of intestinal morphology, microflora, and barrier integrity in broilers. Poult. Sci. 2017, 96, 4325–4332. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Song, M.; Yun, W.; Lee, J.; Lee, C.; Kwak, W.; Han, N.; Kim, H.; Cho, J. Effects of oral administration of different dosages of carvacrol essential oils on intestinal barrier function in broilers. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Elson, C.E. Suppression of mevalonate pathway activities by dietary isoprenoids: Protective roles in cancer and cardiovascular disease. J. Nutr. 1995, 125 (Suppl. S6), 1666S–1672S. [Google Scholar]
- Moeser, A.J.; Klok, C.V.; Ryan, K.A.; Wooten, J.G.; Little, D.; Cook, V.L.; Blikslager, A. Stress signaling pathways activated by weaning mediate intestinal dysfunction in the pig. Am. J. Physiol. Gastroint. Liver Physiol. 2007, 292, G173–G181. [Google Scholar] [CrossRef] [PubMed]
- Bergantini, L.; Bianchi, F.; Cameli, P.; Mazzei, M.A.; Fui, A.; Sestini, P.; Rottoli, P.; Bargagli, E. Prognostic Biomarkers of Sarcoidosis: A Comparative Study of Serum Chitotriosidase, ACE, Lysozyme, and KL-6. Dis. Markers 2019, 2019, 8565423. [Google Scholar] [CrossRef]
- Klass, H.; Neale, G. Serum and faecal lysozyme in inflammatory bowel disease. Gut 1978, 19, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Pié, S.; Lalles, J.P.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I.P. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Sève, B. Effects of underfeeding during the weaning period on growth, metabolism, and hormonal adjustments in the piglet. Domest. Anim. Endocrinol. 2000, 19, 63–74. [Google Scholar]
- Boudry, G.; Péron, V.; Le Huërou-Luron, I.; Lalles, J.P.; Sève, B. Weaning induces both transient and long-lasting modifications of absorptive, secretory, and barrier properties of piglet intestine. J. Nutr. 2004, 134, 2256–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocana-Fuentes, A.; Arranz-Gutierrez, E.; Senorans, F.; Reglero, G. Supercritical fluid extraction of oregano (Origanum vulgare) essentials oils: Anti-inflammatory properties based on cytokine response on THP-1 macrophages. Food Chem. Toxicol. 2010, 48, 1568–1575. [Google Scholar] [CrossRef] [PubMed]
- Ballabeni, V.; Tognolini, M.; Giorgio, C.; Bertoni, S.; Bruni, R.; Barocelli, E. Ocotea quixos Lam. essential oil: In vitro and in vivo investigation on its anti-inflammatory properties. Fitoterapia 2010, 81, 289–295. [Google Scholar] [CrossRef]
- Maynard, C.L.; Elson, C.O.; Hatton, R.D.; Weaver, C.T. Reciprocal interactions of the intestinal microbiota and immune system. Nature 2012, 489, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Luo, J.; Yang, S.; Xiao, Q.; Wang, X.; Zhou, Z.; Xiao, Y.; Shi, D. Different Responses of Microbiota across Intestinal Tract to Enterococcus faecium HDRsEf1 and Their Correlation with Inflammation in Weaned Piglets. Microorganisms 2021, 9, 1767. [Google Scholar] [CrossRef]
- Jiang, Z.; Su, W.; Li, W.; Wen, C.; Du, S.; He, H.; Zhang, Y.; Gong, T.; Wang, X.; Wang, Y.; et al. Bacillus amyloliquefaciens 40 regulates piglet performance, antioxidant capacity, immune status and gut microbiota. Anim. Nutr. 2022, 12, 116–127. [Google Scholar] [CrossRef]
- Hu, R.; He, Z.; Liu, M.; Tan, J.; Zhang, H.; Hou, D.-X.; Wu, S. Dietary protocatechuic acid ameliorates inflammation and up-regulates intestinal tight junction proteins by modulating gut microbiota in LPS-challenged piglets. J. Anim. Sci. Biotechnol. 2020, 11, 92. [Google Scholar] [CrossRef]
- Pujo, J.; Petitfils, C.; Le Faouder, P.; Eeckhaut, V.; Payros, G.; Maurel, S.; Perez-Berezo, T.; van Hul, M.; Barreau, F.; Blanpied, C.; et al. Bacteria-derived long chain fatty acid exhibits anti-inflammatory properties in colitis. Gut 2021, 70, 1088–1097. [Google Scholar] [CrossRef]
- Ritchie, J.M.; Rui, H.; Zhou, X.; Iida, T.; Kodoma, T.; Ito, S.; Davis, B.M.; Bronson, R.T.; Waldor, M.K. Inflammation and disintegration of intestinal villi in an experimental model for Vibrio parahaemolyticus-induced diarrhea. PLoS Pathog. 2012, 8, e1002593. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Li, Y.; Han, H.; Liu, Z.; Zeng, X.; Li, T.; Yin, Y. Long-term effects of lysine concentration on growth performance.; intestinal microbiome.; and metabolic profiles in a pig model. Food Funct. 2018, 9, 4153–4163. [Google Scholar] [CrossRef] [PubMed]
- Pieper, R.; Kröger, S.; Richter, J.F.; Wang, J.; Martin, L.; Bindelle, J.; Htoo, J.; Von Smolinski, D.; Vahjen, W.; Zentek, J. Fermentable fiber ameliorates fermentable protein-induced changes in microbial ecology, but not the mucosal response, in the colon of piglets. J. Nutr. 2012, 142, 661–667. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhou, X.; Huang, N.; Li, H.; Cui, Z.; Tian, J.; Jiang, Q.; Liu, S.; Wu, J.; Li, T. Administration of alpha-ketoglutarate improves epithelial restitution under stress injury in early-weaning piglets. Oncotarget 2017, 8, 91965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.; Bai, Y.; Li, T.; Li, N.; Wang, J. Original low birth weight deteriorates the hindgut epithelial barrier function in pigs at the growing stage. FASEB J. 2019, 33, 9897–9912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Sun, J.; Ge, L.; Liu, Z.; Chen, H.; Yu, B.; Chen, D. Exogenous infusion of short-chain fatty acids can improve intestinal functions independently of the gut microbiota. J Anim Sci. 2020, 98, skaa371. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition, % | Phase 1, Days 1–10 | Phase 2, Days 11–28 |
|---|---|---|
| Extruded corn | 30.53 | 30 |
| Corn | 25 | 32.42 |
| Soybean meal | 8.55 | 10.1 |
| Soybean protein concentrate | 8 | 8 |
| Whey powder | 8 | 5 |
| Peru fish meal | 6 | 5 |
| Limestone | 0.2 | 0.2 |
| Soybean oil | 2 | 2 |
| Sucrose | 2 | - |
| Glucose | 5 | 3 |
| Calcium formate | 0.6 | 0.6 |
| Calcium hydrogen phosphate | 0.5 | 0.4 |
| Choline chloride | 0.1 | 0.1 |
| Premix 1 | 1 | 1 |
| Antioxidant | 0.05 | 0.05 |
| Citric acid | 0.8 | 0.8 |
| ZnO | 0.2 | 0.02 |
| Salt | 0.4 | 0.4 |
| Lysine | 0.62 | 0.55 |
| DL-methionine | 0.09 | 0.07 |
| Threonine | 0.25 | 0.2 |
| Tryptophan | 0.06 | 0.04 |
| Fungicide | 0.05 | 0.05 |
| Total | 100 | 100 |
| Calculated levels, % | ||
| CP | 19 | 18.5 |
| Ca | 0.79 | 0.7 |
| AP | 0.44 | 0.38 |
| Lysine | 1.35 | 1.23 |
| Methionine | 0.39 | 0.36 |
| Threonine | 0.79 | 0.73 |
| Tryptophan | 0.22 | 0.2 |
| DE MJ/kg | 14.67 | 14.57 |
| Control | EO | p-Value | |
|---|---|---|---|
| Body weight (kg) | |||
| Day 0 | 7.37 ± 0.21 | 7.77 ± 0.45 | 0.436 |
| Day 14 | 9.74 ± 0.39 | 10.43 ± 0.62 | 0.364 |
| Day 28 | 15.30 ± 0.62 | 17.82 ± 1.12 | 0.077 |
| ADG (g/d) | |||
| Day 0 to 14 | 169.35 ± 16.16 | 190.18 ± 14.86 | 0.365 |
| Day 14 to 28 | 430.00 ± 11.59 | 527.38 ± 39.88 | 0.082 |
| Day 0 to 28 | 283.33 ± 16.49 | 358.78 ± 26.49 | 0.036 |
| ADFI (g/d) | |||
| Day 0 to 14 | 373.39 ± 16.39 | 488.54 ± 26.72 | 0.004 |
| Day 14 to 28 | 892.56 ± 38.65· | 1092.76 ± 51.73 | 0.011 |
| Day 0 to 28 | 632.98 ± 25.11 | 790.65 ± 37.94 | 0.006 |
| F/G | |||
| Day 0 to 14 | 2.33 ± 0.27 | 2.63 ± 0.21 | 0.399 |
| Day 14 to 28 | 2.13 ± 0.04 | 2.10 ± 0.10 | 0.807 |
| Day 0 to 28 | 2.25 ± 0.08 | 2.24 ± 0.12 | 0.921 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, Y.; Peng, Q.; Wu, Y.; Peng, C.; Wang, S.; Zou, L.; Qi, M.; Peng, C.; Liu, H.; Li, R.; et al. The Effect of an Essential Oil Blend on Growth Performance, Intestinal Health, and Microbiota in Early-Weaned Piglets. Nutrients 2023, 15, 450. https://doi.org/10.3390/nu15020450
Shao Y, Peng Q, Wu Y, Peng C, Wang S, Zou L, Qi M, Peng C, Liu H, Li R, et al. The Effect of an Essential Oil Blend on Growth Performance, Intestinal Health, and Microbiota in Early-Weaned Piglets. Nutrients. 2023; 15(2):450. https://doi.org/10.3390/nu15020450
Chicago/Turabian StyleShao, Yirui, Qingyun Peng, Yuliang Wu, Changfeng Peng, Shanshan Wang, Lijun Zou, Ming Qi, Can Peng, Hongnan Liu, Rui Li, and et al. 2023. "The Effect of an Essential Oil Blend on Growth Performance, Intestinal Health, and Microbiota in Early-Weaned Piglets" Nutrients 15, no. 2: 450. https://doi.org/10.3390/nu15020450
APA StyleShao, Y., Peng, Q., Wu, Y., Peng, C., Wang, S., Zou, L., Qi, M., Peng, C., Liu, H., Li, R., Xiong, X., & Yin, Y. (2023). The Effect of an Essential Oil Blend on Growth Performance, Intestinal Health, and Microbiota in Early-Weaned Piglets. Nutrients, 15(2), 450. https://doi.org/10.3390/nu15020450