
Maternal Protein Restriction in Rats Alters Postnatal Growth and Brain Lipid Sensing in Female Offspring


Abstract
:1. Introduction
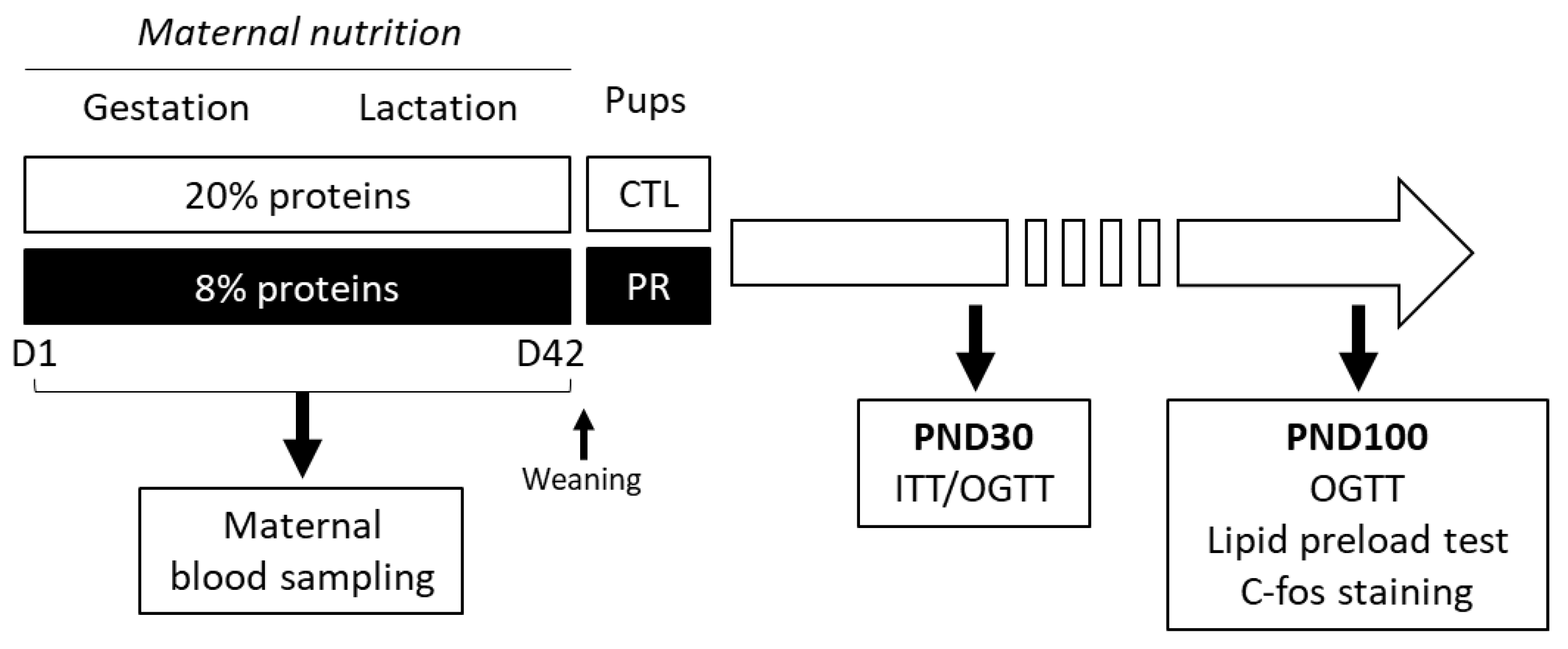
2. Materials and Methods
3. Results
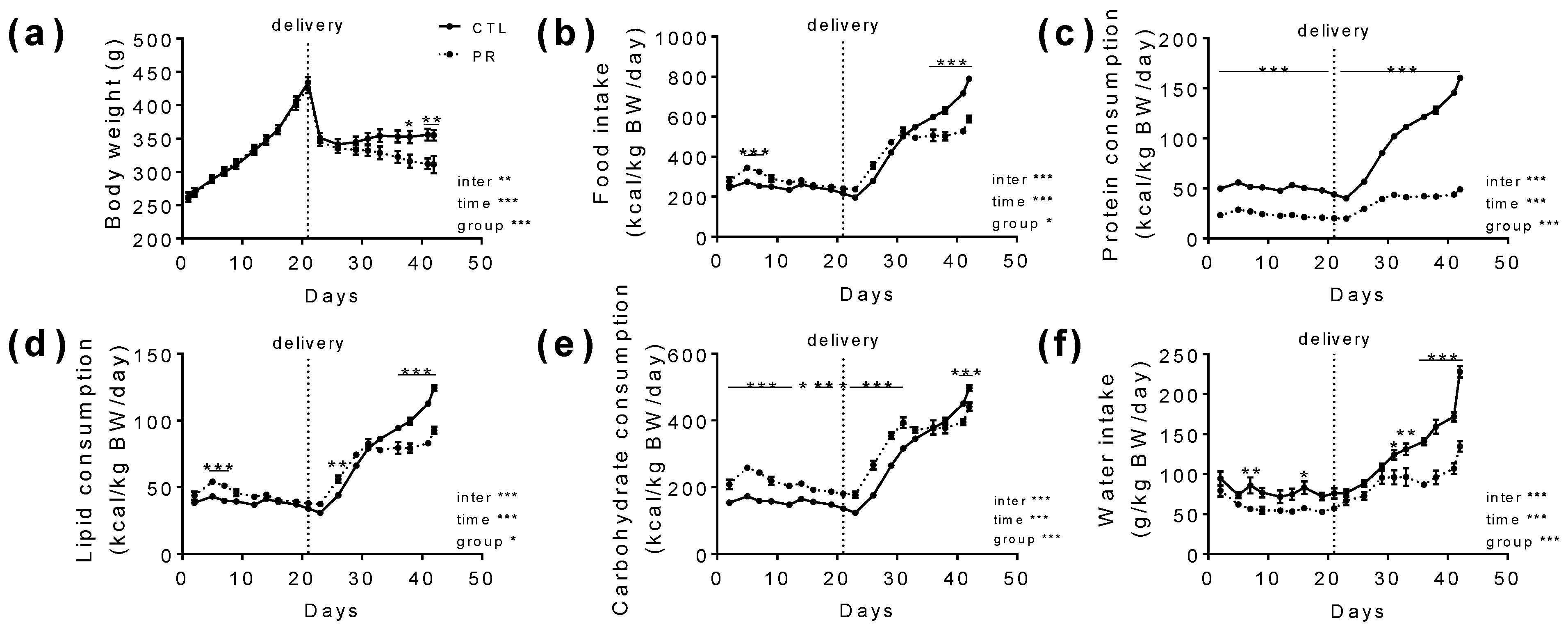
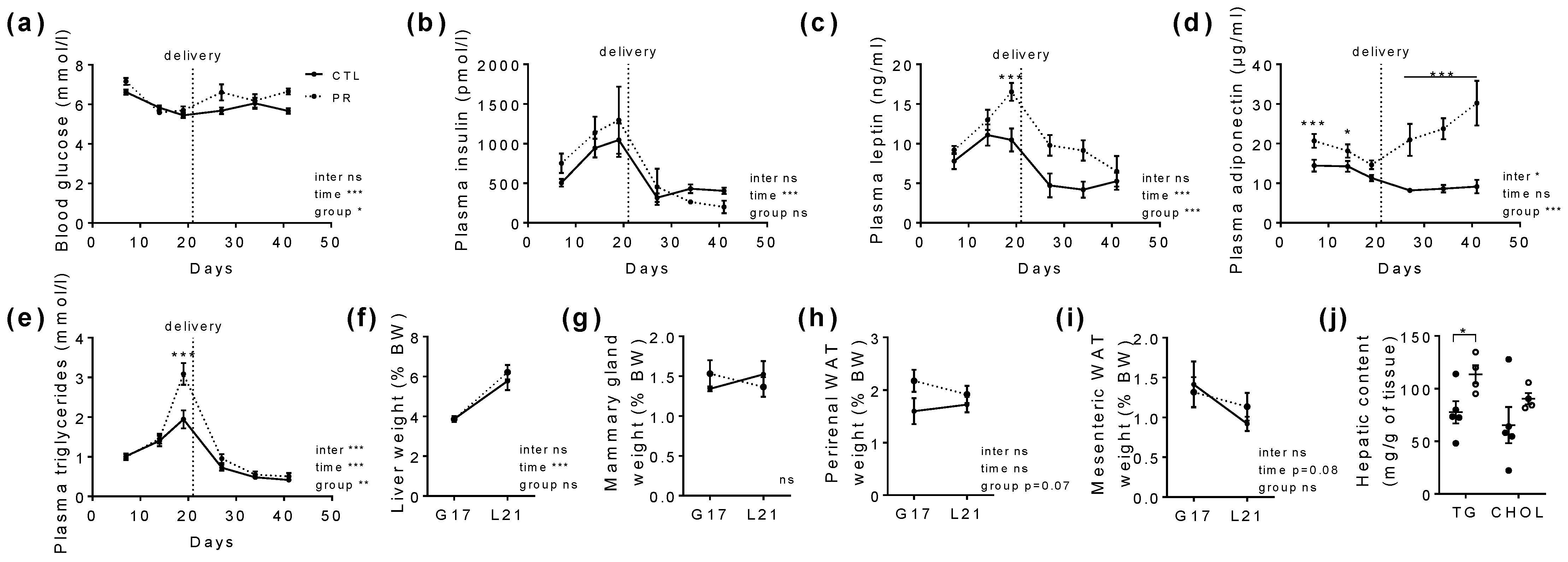
3.1. Maternal PR Does Alter Maternal Food Intake, Body Weight Gain, and Hormonal Status during Gestation and Lactation
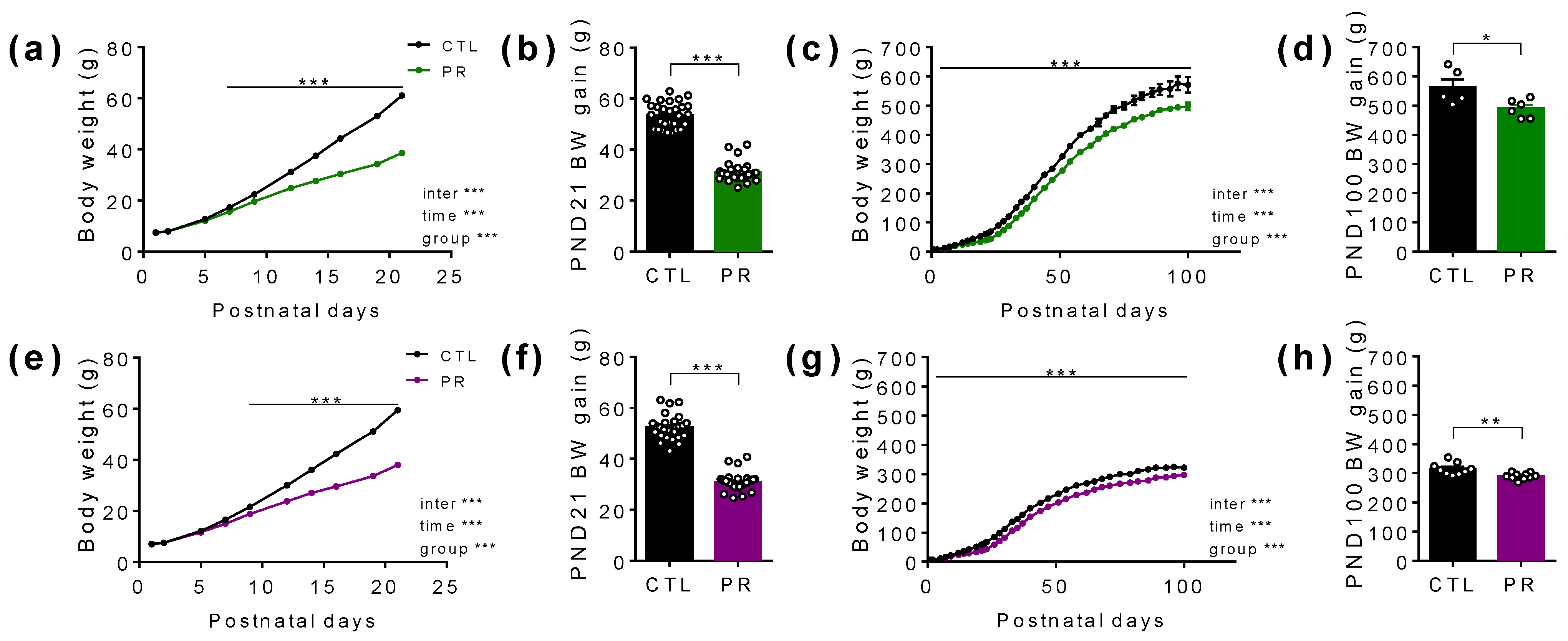
3.2. Despite Unchanged Birth Weight, Maternal PR Has Long Lasting Effects after Birth on the Body Weight of the Offspring
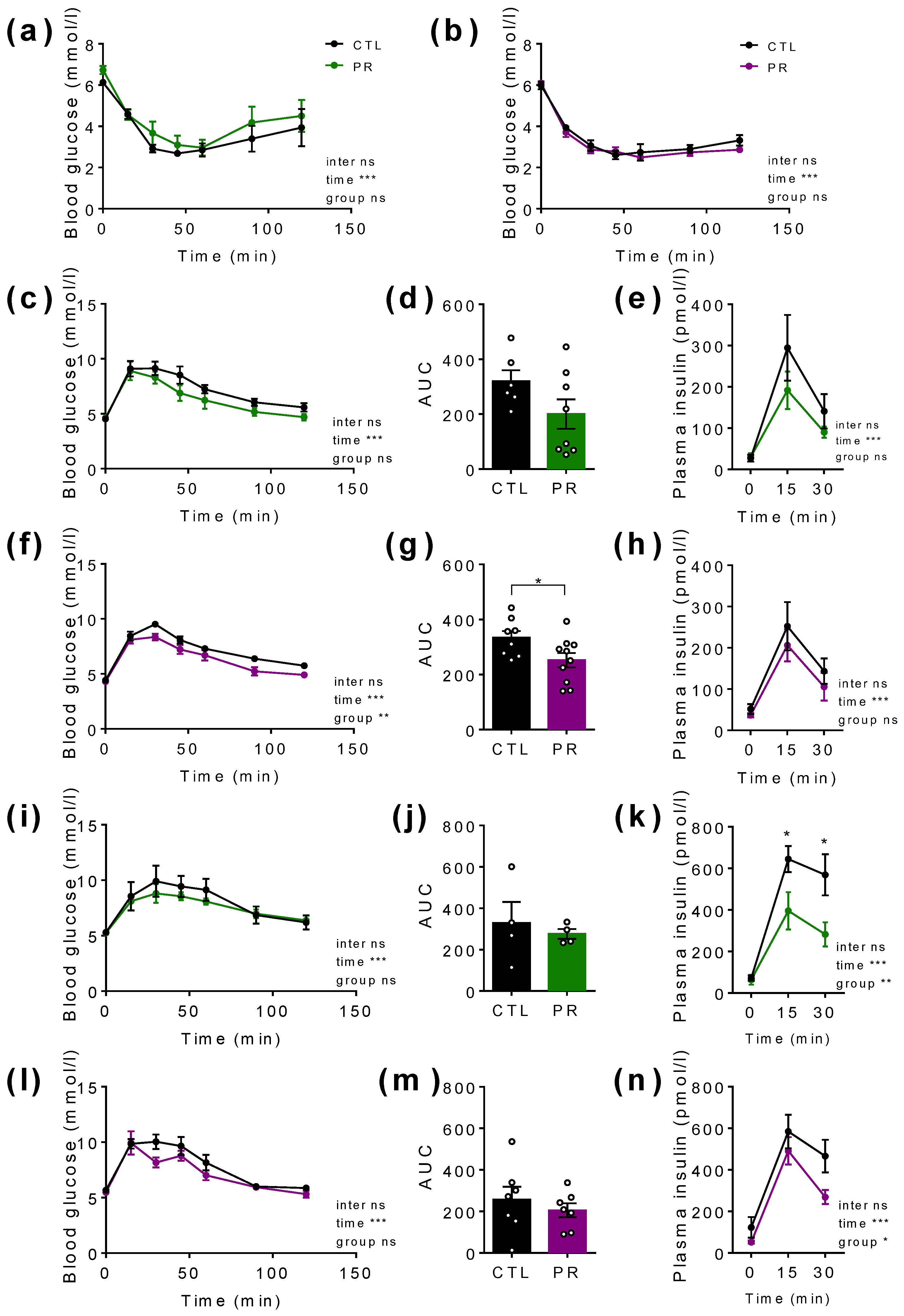
3.3. Maternal PR Has a Small but Significant Impact on Glucose Tolerance in Young (PND30) Male and Female Offspring without Significantly Affecting Insulin Secretion
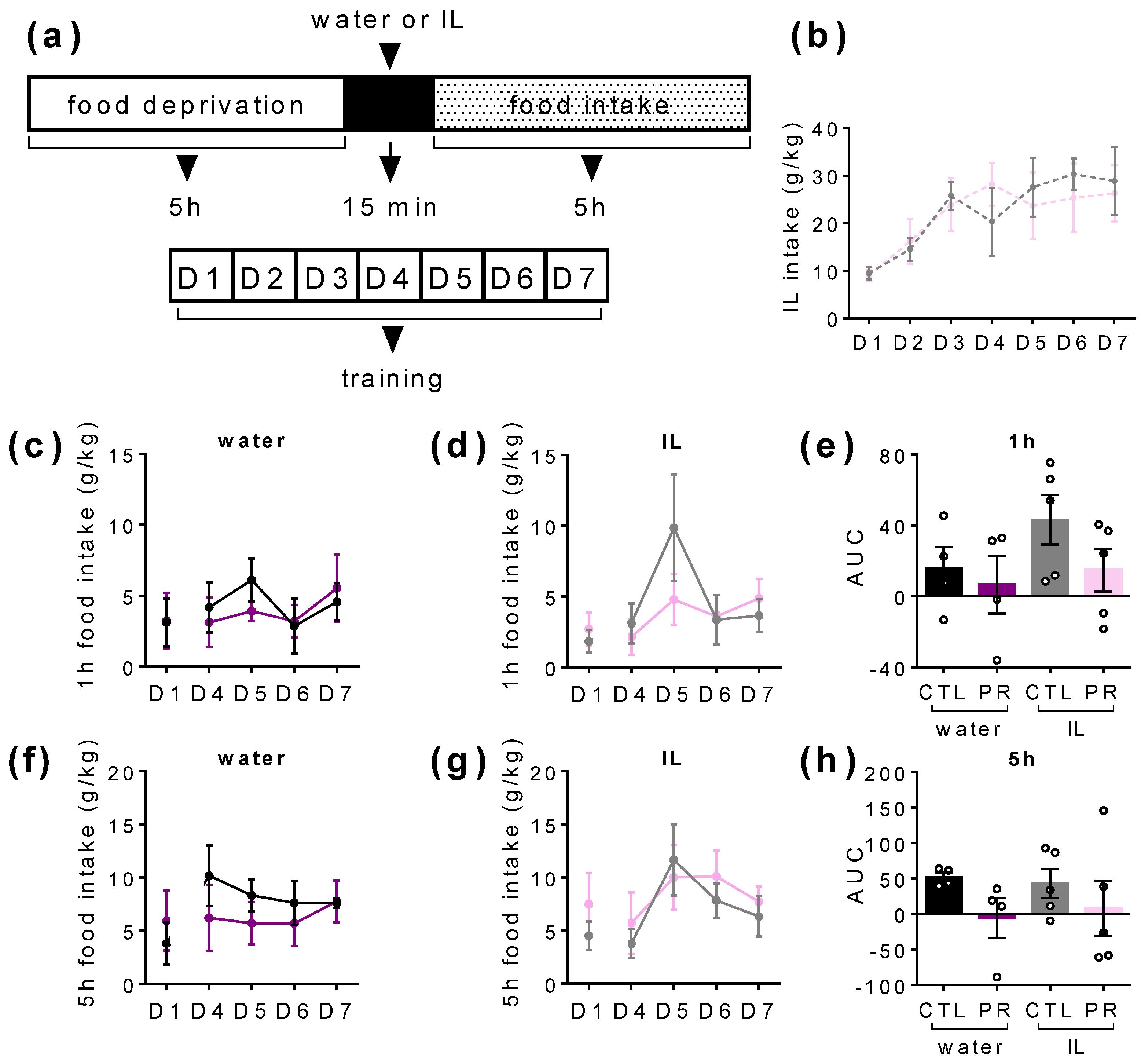
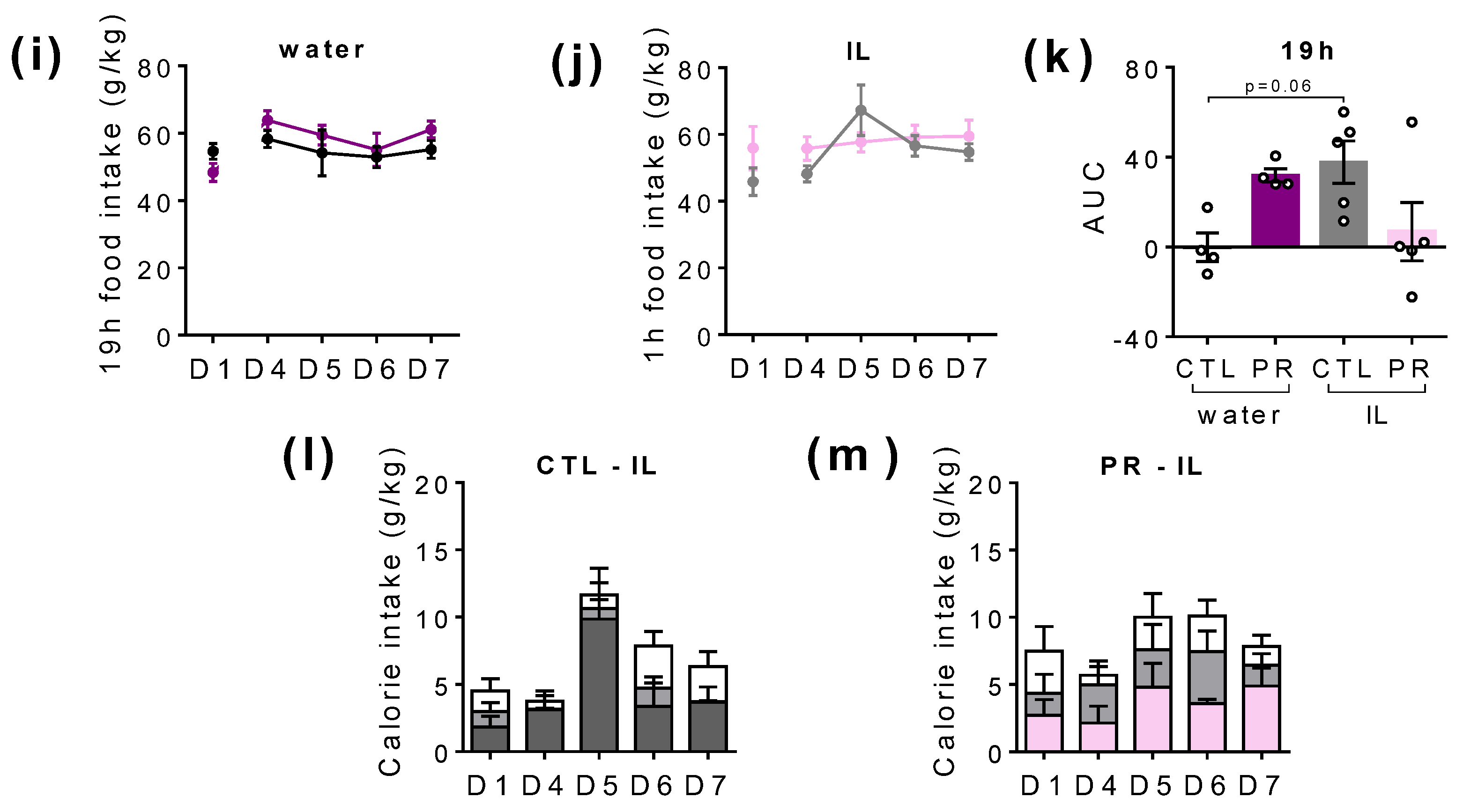
3.4. Maternal PR Modifies Lipid Sensing in Female Offspring at PND100
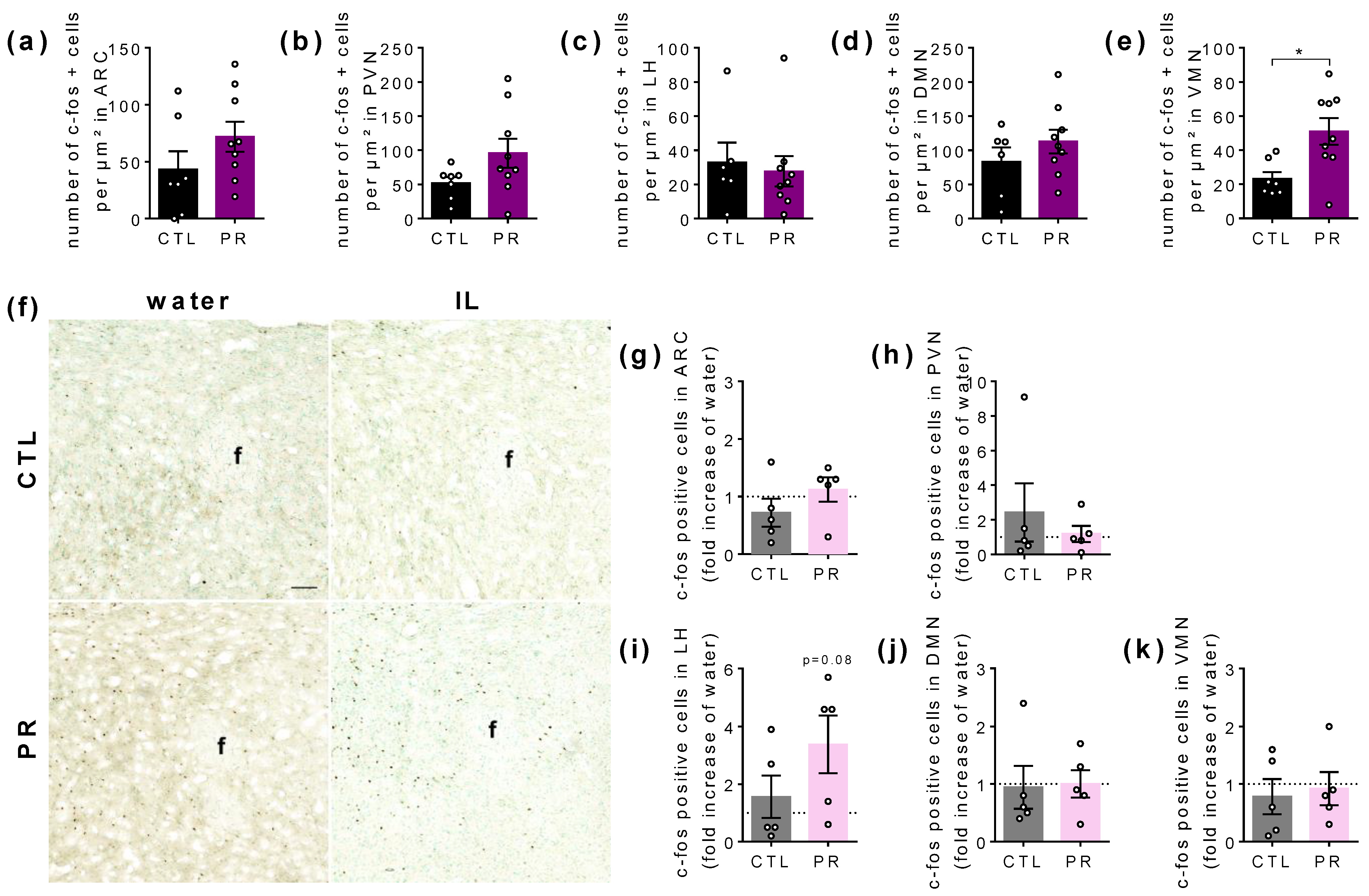
3.5. Maternal PR Changes the Number of c-fos Immunoreactive Cells in Hypothalamus in Basal State
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Lancet Global Health; UNICEF. WHO 1 in 7 Babies Worldwide Born with a Low Birthweight—The Lancet Global Health, UNICEF, WHO. Available online: https://www.unicef.org/press-releases/1-7-babies-worldwide-born-low-birthweight-lancet-global-health-unicef-who (accessed on 1 October 2021).
- Barker, D.J.; Hales, C.N.; Fall, C.H.; Osmond, C.; Phipps, K.; Clark, P.M. Type 2 (Non-Insulin-Dependent) Diabetes Mellitus, Hypertension and Hyperlipidaemia (Syndrome X): Relation to Reduced Fetal Growth. Diabetologia 1993, 36, 62–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gluckman, P.D.; Hanson, M.A. The Developmental Origins of the Metabolic Syndrome. Trends Endocrinol. Metab. 2004, 15, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.M.; Barker, D.J. Fetal Nutrition and Adult Disease. Am. J. Clin. Nutr. 2000, 71, 1344S–1352S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Liu, J.; Qi, L.; Adachi, J.D.; Wu, J.; Li, Z.; Meng, Q.; Li, G.; Lip, G.Y.H. Birth Weight and the Risk of Cardiovascular Outcomes: A Report From the Large Population-Based UK Biobank Cohort Study. Front. Cardiovasc. Med. 2022, 9, 595. [Google Scholar] [CrossRef] [PubMed]
- Santana-Meneses, J.F.; Suano-Souza, F.I.; Franco, M.d.C.; Fonseca, F.L.; Strufaldi, M.W.L. Leptin and Adiponectin Concentrations in Infants with Low Birth Weight: Relationship with Maternal Health and Postnatal Growth. J. Dev. Orig. Health Dis. 2022, 13, 338–344. [Google Scholar] [CrossRef]
- Stefaniak, M.; Dmoch-Gajzlerska, E. Maternal Serum and Cord Blood Leptin Concentrations at Delivery in Normal Pregnancies and in Pregnancies Complicated by Intrauterine Growth Restriction. Obes. Facts 2021, 15, 62–69. [Google Scholar] [CrossRef]
- Schneider-Worthington, C.R.; Bahorski, J.S.; Fields, D.A.; Gower, B.A.; Fernández, J.R.; Chandler-Laney, P.C. Associations Among Maternal Adiposity, Insulin, and Adipokines in Circulation and Human Milk. J. Hum. Lact. 2021, 37, 714–722. [Google Scholar] [CrossRef]
- Briana, D.D.; Malamitsi-Puchner, A. Intrauterine Growth Restriction and Adult Disease: The Role of Adipocytokines. Eur. J. Endocrinol. 2009, 160, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Ayres, C.; Agranonik, M.; Portella, A.K.; Filion, F.; Johnston, C.C.; Silveira, P.P. Intrauterine Growth Restriction and the Fetal Programming of the Hedonic Response to Sweet Taste in Newborn Infants. Int. J. Pediatr. 2012, 2012, 657379. [Google Scholar] [CrossRef]
- Silveira, P.P.; Agranonik, M.; Faras, H.; Portella, A.K.; Meaney, M.J.; Levitan, R.D. Preliminary Evidence for an Impulsivity-Based Thrifty Eating Phenotype. Pediatr. Res. 2012, 71, 293–298. [Google Scholar] [CrossRef]
- Barbieri, M.A.; Portella, A.K.; Silveira, P.P.; Bettiol, H.; Agranonik, M.; Silva, A.A.; Goldani, M.Z. Severe Intrauterine Growth Restriction Is Associated With Higher Spontaneous Carbohydrate Intake in Young Women. Pediatr. Res. 2009, 65, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, A.R.; Portella, A.K.; Paquet, C.; Molle, R.D.; Faber, A.; Arora, N.; Levitan, R.D.; Silveira, P.P.; Dube, L. Low Birth Weight Is Associated with Increased Fat Intake in School-Aged Boys. Br. J. Nutr. 2018, 119, 1295–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blouet, C.; Schwartz, G.J. Hypothalamic Nutrient Sensing in the Control of Energy Homeostasis. Behav. Brain Res. 2010, 209, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Briffa, J.F.; O’Dowd, R.; Moritz, K.M.; Romano, T.; Jedwab, L.R.; McAinch, A.J.; Hryciw, D.H.; Wlodek, M.E. Uteroplacental Insufficiency Reduces Rat Plasma Leptin Concentrations and Alters Placental Leptin Transporters: Ameliorated with Enhanced Milk Intake and Nutrition. J. Physiol. 2017, 595, 3389–3407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coupe, B.; Grit, I.; Hulin, P.; Randuineau, G.; Parnet, P. Postnatal Growth after Intrauterine Growth Restriction Alters Central Leptin Signal and Energy Homeostasis. PLoS ONE 2012, 7, e30616. [Google Scholar] [CrossRef] [Green Version]
- Delahaye, F.; Breton, C.; Risold, P.Y.; Enache, M.; Dutriez-Casteloot, I.; Laborie, C.; Lesage, J.; Vieau, D. Maternal Perinatal Undernutrition Drastically Reduces Postnatal Leptin Surge and Affects the Development of Arcuate Nucleus Proopiomelanocortin Neurons in Neonatal Male Rat Pups. Endocrinology 2008, 149, 470–475. [Google Scholar] [CrossRef]
- Puglianiello, A.; Germani, D.; Antignani, S.; Tomba, G.S.; Cianfarani, S. Changes in the Expression of Hypothalamic Lipid Sensing Genes in Rat Model of Intrauterine Growth Retardation (IUGR). Pediatr. Res. 2007, 61, 433–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coupe, B.; Grit, I.; Darmaun, D.; Parnet, P. The Timing of “Catch-up Growth” Affects Metabolism and Appetite Regulation in Male Rats Born with Intrauterine Growth Restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R813–R824. [Google Scholar] [CrossRef] [Green Version]
- Coupe, B.; Amarger, V.; Grit, I.; Benani, A.; Parnet, P. Nutritional Programming Affects Hypothalamic Organization and Early Response to Leptin. Endocrinology 2010, 151, 702–713. [Google Scholar] [CrossRef]
- Orozco-Solis, R.; Matos, R.J.; Guzman-Quevedo, O.; Lopes de Souza, S.; Bihouee, A.; Houlgatte, R.; Manhaes de Castro, R.; Bolanos-Jimenez, F. Nutritional Programming in the Rat Is Linked to Long-Lasting Changes in Nutrient Sensing and Energy Homeostasis in the Hypothalamus. PLoS ONE 2010, 5, e13537. [Google Scholar] [CrossRef]
- Martin Agnoux, A.; Alexandre-Gouabau, M.C.; Le Drean, G.; Antignac, J.P.; Parnet, P. Relative Contribution of Foetal and Post-Natal Nutritional Periods on Feeding Regulation in Adult Rats. Acta Physiol. 2014, 210, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Dalle Molle, R.; Laureano, D.P.; Alves, M.B.; Reis, T.M.; Desai, M.; Ross, M.G.; Silveira, P.P. Intrauterine Growth Restriction Increases the Preference for Palatable Foods and Affects Sensitivity to Food Rewards in Male and Female Adult Rats. Brain Res. 2015, 1618, 41–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luquet, S.; Magnan, C. The Central Nervous System at the Core of the Regulation of Energy Homeostasis. Front. Biosci. 2009, 1, 448–465. [Google Scholar] [CrossRef]
- Oomura, Y.; Nakamura, T.; Sugimori, M.; Yamada, Y. Effect of Free Fatty Acid on the Rat Lateral Hypothalamic Neurons. Physiol. Behav. 1975, 14, 483–486. [Google Scholar] [CrossRef]
- Moullé, V.S.; Le Foll, C.; Philippe, E.; Kassis, N.; Rouch, C.; Marsollier, N.; Bui, L.C.; Guissard, C.; Dairou, J.; Lorsignol, A.; et al. Fatty Acid Transporter CD36 Mediates Hypothalamic Effect of Fatty Acids on Food Intake in Rats. PLoS ONE 2013, 8, e74021. [Google Scholar] [CrossRef] [PubMed]
- Obici, S.; Feng, Z.; Morgan, K.; Stein, D.; Karkanias, G.; Rossetti, L. Central Administration of Oleic Acid Inhibits Glucose Production and Food Intake. Diabetes 2002, 51, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Cruciani-Guglielmacci, C.; Migrenne, S.; Magnan, C.; Cotero, V.E.; Routh, V.H. Effects of Oleic Acid on Distinct Populations of Neurons in the Hypothalamic Arcuate Nucleus Are Dependent on Extracellular Glucose Levels. J. Neurophysiol. 2006, 95, 1491–1498. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.; Li, T.; Ross, M.G. Hypothalamic Neurosphere Progenitor Cells in Low Birth-Weight Rat Newborns: Neurotrophic Effects of Leptin and Insulin. Brain Res. 2011, 1378, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.; Li, T.; Ross, M.G. Fetal Hypothalamic Neuroprogenitor Cell Culture: Preferential Differentiation Paths Induced by Leptin and Insulin. Endocrinology 2011, 152, 3192–3201. [Google Scholar] [CrossRef] [PubMed]
- Frapin, M.; Guignard, S.; Meistermann, D.; Grit, I.; Moullé, V.S.; Paillé, V.; Parnet, P.; Amarger, V. Maternal Protein Restriction in Rats Alters the Expression of Genes Involved in Mitochondrial Metabolism and Epitranscriptomics in Fetal Hypothalamus. Nutrients 2020, 12, 1464. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates—The New Coronal Set, 5th ed.; Academic Press: Orlando, FL, USA, 2004; ISBN 978-0-08-047412-0. [Google Scholar]
- Ceolin, P.; França, S.a.D.; Froelich, M.; Santos, M.P.D.; Pereira, M.P.; Queiroz, T.S.; Silva, F.H.S.D.; Lisboa, P.C.; Andrade, C.M.B.; Baviera, A.M.; et al. A Low-Protein, High Carbohydrate Diet Induces Increase in Serum Adiponectin and Preserves Glucose Homeostasis in Rats. An. Acad. Bras. Ciênc. 2019, 91, e20180452. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Higginbotham, D.A.; White, B.D. Food Intake, Energy Balance and Serum Leptin Concentrations in Rats Fed Low-Protein Diets. J. Nutr. 2000, 130, 514–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehring, J.; Gaudichon, C.; Even, P.C. Food Intake Control and Body Weight Regulation by Dietary Protein. Cah. De Nutr. Et De Diététique 2020, 55, e1–e8. [Google Scholar] [CrossRef]
- Sevrin, T.; Alexandre-Gouabau, M.-C.; Castellano, B.; Aguesse, A.; Ouguerram, K.; Ngyuen, P.; Darmaun, D.; Boquien, C.-Y. Impact of Fenugreek on Milk Production in Rodent Models of Lactation Challenge. Nutrients 2019, 11, 2571. [Google Scholar] [CrossRef] [Green Version]
- Martin Agnoux, A.; Antignac, J.P.; Boquien, C.Y.; David, A.; Desnots, E.; Ferchaud-Roucher, V.; Darmaun, D.; Parnet, P.; Alexandre-Gouabau, M.C. Perinatal Protein Restriction Affects Milk Free Amino Acid and Fatty Acid Profile in Lactating Rats: Potential Role on Pup Growth and Metabolic Status. J. Nutr. Biochem. 2015, 26, 784–795. [Google Scholar] [CrossRef]
- Sevrin, T.; Alexandre-Gouabau, M.-C.; Darmaun, D.; Palvadeau, A.; André, A.; Nguyen, P.; Ouguerram, K.; Boquien, C.-Y. Use of Water Turnover Method to Measure Mother’s Milk Flow in a Rat Model: Application to Dams Receiving a Low Protein Diet during Gestation and Lactation. PLoS ONE 2017, 12, e0180550. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.F.; Armstrong, L.E.; Rodriguez, N.R. Dietary Protein Intake and Renal Function. Nutr. Metab. 2005, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, A.; Henkel, H.; Verstegen, M.W.A.; Philipczyk, I. The Influence of Protein Intake on Water Balance, Flow Rate and Apparent Digestibilty of Nutrients at the Distal Ileum in Growing Pigs. Livest. Prod. Sci. 1995, 44, 179–187. [Google Scholar] [CrossRef]
- Yagi, T.; Toyoshima, Y.; Tokita, R.; Taguchi, Y.; Okamoto, Y.; Takahashi, S.-I.; Kato, H.; Minami, S. Low-Protein Diet Enhances Adiponectin Secretion in Rats. Biosci. Biotechnol. Biochem. 2019, 83, 1774–1781. [Google Scholar] [CrossRef]
- Tessier, D.R.; Ferraro, Z.M.; Gruslin, A. Role of Leptin in Pregnancy: Consequences of Maternal Obesity. Placenta 2013, 34, 205–211. [Google Scholar] [CrossRef]
- Mazaki-Tovi, S.; Kanety, H.; Pariente, C.; Hemi, R.; Wiser, A.; Schiff, E.; Sivan, E. Maternal Serum Adiponectin Levels during Human Pregnancy. J. Perinatol. 2007, 27, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Bouret, S.G. Nutritional Programming of Hypothalamic Development: Critical Periods and Windows of Opportunity. Int. J. Obes. Suppl. 2012, 2, S19–S24. [Google Scholar] [CrossRef]
- Yura, S.; Itoh, H.; Sagawa, N.; Yamamoto, H.; Masuzaki, H.; Nakao, K.; Kawamura, M.; Takemura, M.; Kakui, K.; Ogawa, Y.; et al. Role of Premature Leptin Surge in Obesity Resulting from Intrauterine Undernutrition. Cell Metab. 2005, 1, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aye, I.L.M.H.; Jansson, T.; Powell, T.L. TNF-α Stimulates System A Amino Acid Transport in Primary Human Trophoblast Cells Mediated by P38 MAPK Signaling. Physiol. Rep. 2015, 3, e12594. [Google Scholar] [CrossRef] [PubMed]
- Nozhenko, Y.; Asnani-Kishnani, M.; Rodríguez, A.M.; Palou, A. Milk Leptin Surge and Biological Rhythms of Leptin and Other Regulatory Proteins in Breastmilk. PLoS ONE 2015, 10, e0145376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agnoux, A.M.; Antignac, J.P.; Simard, G.; Poupeau, G.; Darmaun, D.; Parnet, P.; Alexandre-Gouabau, M.C. Time Window-Dependent Effect of Perinatal Maternal Protein Restriction on Insulin Sensitivity and Energy Substrate Oxidation in Adult Male Offspring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R184–R197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breton, C.; Lukaszewski, M.A.; Risold, P.Y.; Enache, M.; Guillemot, J.; Riviere, G.; Delahaye, F.; Lesage, J.; Dutriez-Casteloot, I.; Laborie, C.; et al. Maternal Prenatal Undernutrition Alters the Response of POMC Neurons to Energy Status Variation in Adult Male Rat Offspring. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E462–E472. [Google Scholar] [CrossRef] [Green Version]
- Dellschaft, N.S.; Alexandre-Gouabau, M.C.; Gardner, D.S.; Antignac, J.P.; Keisler, D.H.; Budge, H.; Symonds, M.E.; Sebert, S.P. Effect of Pre- and Postnatal Growth and Post-Weaning Activity on Glucose Metabolism in the Offspring. J. Endocrinol. 2015, 224, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Tarry-Adkins, J.L.; Fernandez-Twinn, D.S.; Madsen, R.; Chen, J.H.; Carpenter, A.; Hargreaves, I.P.; McConnell, J.M.; Ozanne, S.E. Coenzyme Q10 Prevents Insulin Signaling Dysregulation and Inflammation Prior to Development of Insulin Resistance in Male Offspring of a Rat Model of Poor Maternal Nutrition and Accelerated Postnatal Growth. Endocrinology 2015, 156, 3528–3537. [Google Scholar] [CrossRef] [Green Version]
- Alves, M.B.; Laureano, D.P.; Dalle Molle, R.; Machado, T.D.; Salvador, A.P.d.A.; Miguel, P.M.; Lupinsky, D.; Dalmaz, C.; Silveira, P.P. Intrauterine Growth Restriction Increases Impulsive Behavior and Is Associated with Altered Dopamine Transmission in Both Medial Prefrontal and Orbitofrontal Cortex in Female Rats. Physiol. Behav. 2019, 204, 336–346. [Google Scholar] [CrossRef]
- Laureano, D.P.; Alves, M.B.; Miguel, P.M.; Machado, T.D.; Reis, A.R.; Mucellini, A.B.; Cunha, F.S.; Lampert, C.; Salvador, A.P.A.; Dalle Molle, R.; et al. Intrauterine Growth Restriction Modifies the Accumbal Dopaminergic Response to Palatable Food Intake. Neuroscience 2019, 400, 184–195. [Google Scholar] [CrossRef] [PubMed]
- French, S.J.; Conlon, C.A.; Mutuma, S.T.; Arnold, M.; Read, N.W.; Meijer, G.; Francis, J. The Effects of Intestinal Infusion of Long-Chain Fatty Acids on Food Intake in Humans. Gastroenterology 2000, 119, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, D.; Smith, G.P.; Gibbs, J. Intraduodenal Infusions of Fats Elicit Satiety in Sham-Feeding Rats. Am. J. Physiol. 1990, 259, R110–R118. [Google Scholar] [CrossRef] [PubMed]
- Gregory, P.C.; Rayner, D.V. The Influence of Gastrointestinal Infusion of Fats on Regulation of Food Intake in Pigs. J. Physiol. 1987, 385, 471–481. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| g/100 g | Control Diet | Low Protein Diet |
|---|---|---|
| Casein | 20 | 8 |
| L-Cystine | 0.3 | 0.3 |
| Corn starch | 39.75 | 47.32 |
| Maltodextrine | 13.20 | 15.72 |
| Sucrose | 10 | 11.91 |
| Cellulose | 5 | 5 |
| Corn oil | 7 | 7 |
| t-butylhydroquinone | 0.0014 | 0.0014 |
| Mineral mix AIN 93 G a | 3.5 | 3.5 |
| Vitamin mix AIN 93 Vx b | 1 | 1 |
| Choline bitartrate | 0.25 | 0.25 |
| Total calories | 3948 | 3959 |
| At Birth | CTL | PR | |
|---|---|---|---|
| Pup number | Total | 11.5 ± 0.3 | 11.3 ± 0.5 |
| Male | 5.7 ± 0.5 | 6.4 ± 0.4 | |
| Female | 5.8 ± 0.6 | 4.9 ± 0.5 | |
| Mortality at birth (%) | 1.2 ± 1.2 | 0.0 ± 0.0 | |
| Litter weight (g) | 79.4 ± 2.3 | 81.9 ± 3.4 | |
| Pup weight (g) | Male | 7.5 ± 0.1 | 7.4 ± 0.1 |
| Female | 7.1 ± 0.1 | 7.0 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moullé, V.S.; Frapin, M.; Amarger, V.; Parnet, P. Maternal Protein Restriction in Rats Alters Postnatal Growth and Brain Lipid Sensing in Female Offspring. Nutrients 2023, 15, 463. https://doi.org/10.3390/nu15020463
Moullé VS, Frapin M, Amarger V, Parnet P. Maternal Protein Restriction in Rats Alters Postnatal Growth and Brain Lipid Sensing in Female Offspring. Nutrients. 2023; 15(2):463. https://doi.org/10.3390/nu15020463
Chicago/Turabian StyleMoullé, Valentine S., Morgane Frapin, Valérie Amarger, and Patricia Parnet. 2023. "Maternal Protein Restriction in Rats Alters Postnatal Growth and Brain Lipid Sensing in Female Offspring" Nutrients 15, no. 2: 463. https://doi.org/10.3390/nu15020463
APA StyleMoullé, V. S., Frapin, M., Amarger, V., & Parnet, P. (2023). Maternal Protein Restriction in Rats Alters Postnatal Growth and Brain Lipid Sensing in Female Offspring. Nutrients, 15(2), 463. https://doi.org/10.3390/nu15020463