
Evaluation of Influencing Factors on Metabolism of Land-Based n-3 Poly Unsaturated Fatty Acids—The KoALA Study


 and
and 
Abstract
:
1. Introduction
2. Materials and Methods
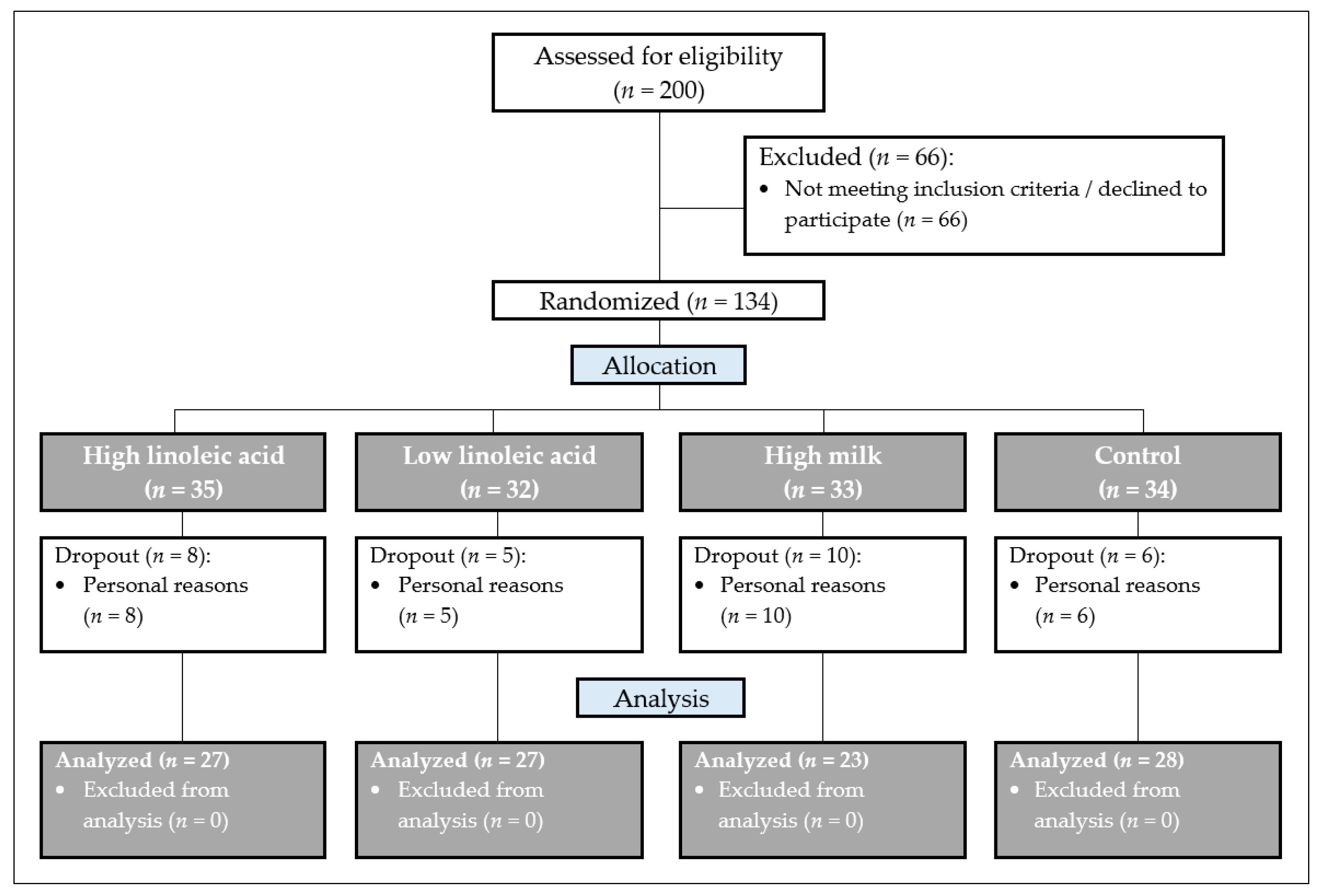
2.1. Study Design (Screening)
- Women (in menopause) and men (50% each), aged between 40 and 65 years, body mass index (BMI) < 30 kg/m2.
- Moderately elevated low-density lipoprotein (LDL) cholesterol (>3 mmol/L).
- Consumption of a traditional “Western diet” composed of meat, sausage, dairy products, cereals, vegetables, fruits, etc.
- Stable eating habits at least one year before enrollment.
- No antihypertensive medication or stable dose for >3 months prior to the start of the study and during the entire study period.
- Acute or chronic diseases which could affect the results of the present study.
- Use of medication, including systemic glucocorticoids or lipid-lowering medication.
- Use of dietary supplements, incl. multivitamins, fish oil capsules, minerals and trace elements (three months before and during the entire study period).
- Weight loss or weight gain (>3 kg) during the last three months before the study began.
- Relevant food allergies (e.g., milk, nuts).
- Pregnancy or lactation.
2.2. Baseline Assessment
2.3. Study Diet—The KoALA Concept
- Absence of other foods providing n-3 PUFA, like fish (oil) and algae (oil).
- Reduced intake of monosaccharides and increased intake of dietary fiber.
- Reduction in salt.
- Increased consumption of fruits and vegetables.
- Reduction in foods which are highly processed, calorie-rich, and low in nutrients, such as fast food and convenience products.
2.4. Sample Collection, Biochemical Analyses, and Further Measurements
2.5. Statistical Methods
3. Results
3.1. Baseline Characteristics of the Study Collective
3.2. Changes in Fatty Acid Distribution in Erythrocyte Lipids within the Diet Groups
3.3. Changes in Fatty Acid Distribution in Erythrocyte Lipids per Diet Group Subdivided into Subgroups by Sex, EPA Status, and LA Change over the Intervention Period
3.4. Influence on Fatty Acid Distribution in Erythrocyte Lipids Depending on Sex, EPA Status, and Total LA Change without the Original Diet Group Split
3.5. Correlation between Micronutrient Status and EPA Percentage Change from Baseline in the Entire Study Population
3.6. Influence on Biochemical Parameters and Anthropometric Measurements per Diet Group
4. Discussion
4.1. Effects on Fatty Acid Distribution in Erythrocytes in Response to Linseed Oil Supplementation
4.2. Discussion of the Observed Effects on Biochemical and Anthropometric Parameters
5. Conclusions
6. Strengths and Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Health Statistics 2016: Monitoring Health for the SDGs; World Health Organization: Geneva, Switzerland, 2016; p. 60. [Google Scholar]
- Statistisches Bundesamt. Causes of Death. Available online: https://www.destatis.de/EN/Themes/Society-Environment/Health/Causes-Death/_node.html (accessed on 29 August 2023).
- Meier, T.; Gräfe, K.; Senn, F.; Sur, P.; Stangl, G.I.; Dawczynski, C.; März, W.; Kleber, M.E.; Lorkowski, S. Cardiovascular mortality attributable to dietary risk factors in 51 countries in the WHO European Region from 1990 to 2016: A systematic analysis of the Global Burden of Disease Study. Eur. J. Epidemiol. 2019, 34, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Max Rubner-Institut. Nationale Verzehrsstudie II, Ergebnisbericht Teil 2. Available online: https://www.bmel.de/SharedDocs/Downloads/DE/_Ernaehrung/NVS_ErgebnisberichtTeil2.pdf?__blob=publicationFile&v=2 (accessed on 27 June 2023).
- Catapano, A.L.; Graham, I.; de Backer, G.; Wiklund, O.; Chapman, M.J.; Drexel, H.; Hoes, A.W.; Jennings, C.S.; Landmesser, U.; Pedersen, T.R.; et al. 2016 ESC/EAS Guidelines for the Management of Dyslipidaemias. Eur. Heart J. 2016, 37, 2999–3058. [Google Scholar] [CrossRef] [PubMed]
- Deutsche Gesellschaft für Ernährung e. V. Evidenzbasierte Leitlinie–Fettzufuhr und Prävention Ernährungsmitbedingter Erkrankungen. Available online: https://www.dge.de/fileadmin/dok/wissenschaft/leitlinien/fette/LL_Fett_Gesamtdokument_Version_2006.pdf (accessed on 27 June 2023).
- Innes, J.K.; Calder, P.C. Marine Omega-3 (N-3) Fatty Acids for Cardiovascular Health: An Update for 2020. Int. J. Mol. Sci. 2020, 21, 1362. [Google Scholar] [CrossRef]
- Wang, C.; Harris, W.S.; Chung, M.; Lichtenstein, A.H.; Balk, E.M.; Kupelnick, B.; Jordan, H.S.; Lau, J. n−3 Fatty acids from fish or fish-oil supplements, but not α-linolenic acid, benefit cardiovascular disease outcomes in primary- and secondary-prevention studies: A systematic review. Am. J. Clin. Nutr. 2006, 84, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Skeaff, C.M.; Miller, J. Dietary Fat and Coronary Heart Disease: Summary of Evidence from Prospective Cohort and Randomised Controlled Trials. Ann. Nutr. Metab. 2009, 55, 173–201. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Mechanisms of Action of (n-3) Fatty Acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef]
- Calder, P.C. Docosahexaenoic Acid. Ann. Nutr. Metab. 2016, 69 (Suppl. 1), 7–21. [Google Scholar] [CrossRef]
- Cambiaggi, L.; Chakravarty, A.; Noureddine, N.; Hersberger, M. The Role of α-Linolenic Acid and Its Oxylipins in Human Cardiovascular Diseases. Int. J. Mol. Sci. 2023, 24, 6110. [Google Scholar] [CrossRef]
- International Society for the Study of Fatty Acids and Lipids. Recommendations for Dietary Intake of Polyunsaturated Fatty Acids in Healthy Adults. Available online: https://www.issfal.org/assets/issfal%2003%20pufaintakereccomdfinalreport.pdf (accessed on 27 June 2023).
- EFSA Panel on Dietetic Products, Nutrition, and Allergies (NDA). Scientific Opinion on Dietary Reference Values for fats, including saturated fatty acids, polyunsaturated fatty acids, monounsaturated fatty acids, trans fatty acids, and cholesterol. EFSA J. 2010, 8, 1461. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Fats and Fatty Acids in Human Nutrition. Available online: https://www.fao.org/3/i1953e/i1953e00.pdf (accessed on 27 June 2023).
- Micha, R.; Khatibzadeh, S.; Shi, P.; Fahimi, S.; Lim, S.; Andrews, K.G.; Engell, R.E.; Powles, J.; Ezzati, M.; Mozaffarian, D.; et al. Global, regional, and national consumption levels of dietary fats and oils in 1990 and 2010: A systematic analysis including 266 country-specific nutrition surveys. BMJ 2014, 348, g2272. [Google Scholar] [CrossRef]
- Sala-Vila, A.; Fleming, J.; Kris-Etherton, P.; Ros, E. Impact of α-Linolenic Acid, the Vegetable ω-3 Fatty Acid, on Cardiovascular Disease and Cognition. Adv. Nutr. Int. Rev. J. 2022, 13, 1584–1602. [Google Scholar] [CrossRef] [PubMed]
- Sala-Vila, A.; Guasch-Ferré, M.; Hu, F.B.; Sánchez-Tainta, A.; Bulló, M.; Serra-Mir, M.; López-Sabater, C.; Sorlí, J.V.; Arós, F.; Fiol, M.; et al. Dietary α-Linolenic Acid, Marine ω-3 Fatty Acids, and Mortality in a Population With High Fish Consumption: Findings From the PREvención con DIeta MEDiterránea (PREDIMED) Study. J. Am. Heart Assoc. 2016, 5, e002543. [Google Scholar] [CrossRef] [PubMed]
- Mente, A.; de Koning, L.; Shannon, H.S.; Anand, S.S. A Systematic Review of the Evidence Supporting a Causal Link Between Dietary Factors and Coronary Heart Disease. JAMA Intern. Med. 2009, 169, 659–669. [Google Scholar] [CrossRef]
- Naghshi, S.; Aune, D.; Beyene, J.; Mobarak, S.; Asadi, M.; Sadeghi, O. Dietary intake and biomarkers of alpha linolenic acid and risk of all cause, cardiovascular, and cancer mortality: Systematic review and dose-response meta-analysis of cohort studies. BMJ 2021, 375, n2213. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Finnegan, Y.E.; Minihane, A.M.; Williams, C.M.; Wootton, S.A. Effect of altered dietaryn-3 fatty acid intake upon plasma lipid fatty acid composition, conversion of [13C]α-linolenic acid to longer-chain fatty acids and partitioning towards β-oxidation in older men. Br. J. Nutr. 2003, 90, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Calder, P.C. Conversion of α-linolenic acid to longer-chain polyunsaturated fatty acids in human adults. Reprod. Nutr. Dev. 2005, 45, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Watkins, B.A. Conjugated linoleic acids alter bone fatty acid composition and reduce ex vivo prostaglandin E2 biosynthesis in rats fed n-6 or n-3 fatty acids. Lipids 1998, 33, 417–425. [Google Scholar] [CrossRef]
- Du, M.; Ahn, D.U.; Sell, J.L. Effects of dietary conjugated linoleic acid and linoleic:linolenic acid ratio on polyunsaturated fatty acid status in laying hens. Poult. Sci. 2000, 79, 1749–1756. [Google Scholar] [CrossRef]
- Drouin, G.; Catheline, D.; Sinquin, A.; Baudry, C.; Le Ruyet, P.; Rioux, V.; Legrand, P. Incorporation of Dairy Lipids in the Diet Increased Long-Chain Omega-3 Fatty Acids Status in Post-weaning Rats. Front. Nutr. 2018, 5, 42. [Google Scholar] [CrossRef]
- Takahashi, Y.; Kushiro, M.; Shinohara, K.; Ide, T. Activity and mRNA levels of enzymes involved in hepatic fatty acid synthe-sis and oxidation in mice fed conjugated linoleic acid. Biochim. Biophys. Acta 2003, 1631, 265–273. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Degen, C.; Ecker, J.; Piegholdt, S.; Liebisch, G.; Schmitz, G.; Jahreis, G. Metabolic and growth inhibitory effects of conjugated fatty acids in the cell line HT-29 with special regard to the conversion of t11,t13-CLA. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2011, 1811, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, M.; Jahreis, G.; Bothor, K.; Drechsel, C.; Kiehntopf, M.; Blüher, M.; Dawczynski, C. Benefits of foods supplemented with vegetable oils rich in α-linolenic, stearidonic or docosahexaenoic acid in hypertriglyceridemic subjects: A double-blind, randomized, controlled trail. Eur. J. Nutr. 2015, 54, 881–893. [Google Scholar] [CrossRef]
- Liou, Y.A.; King, D.J.; Zibrik, D.; Innis, S.M. Decreasing Linoleic Acid with Constant α-Linolenic Acid in Dietary Fats Increases (n-3) Eicosapentaenoic Acid in Plasma Phospholipids in Healthy Men1. J. Nutr. 2007, 137, 945–952. [Google Scholar] [CrossRef]
- MacIntosh, B.A.; Ramsden, C.E.; Faurot, K.R.; Zamora, D.; Mangan, M.; Hibbeln, J.R.; Mann, J.D. Low-n-6 and low-n-6 plus high-n-3 diets for use in clinical research. Br. J. Nutr. 2013, 110, 559–568. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Harris, W.S.; Luo, J.; Pottala, J.V.; Espeland, M.A.; Margolis, K.L.; Manson, J.E.; Wang, L.; Brasky, T.M.; Robinson, J.G. Red blood cell polyunsaturated fatty acids and mortality in the Women’s Health Initiative Memory Study. J. Clin. Lipidol. 2017, 11, 250–259.e5. [Google Scholar] [CrossRef]
- Harris, W.S.; Tintle, N.L.; Etherton, M.R.; Vasan, R.S. Erythrocyte long-chain omega-3 fatty acid levels are inversely associated with mortality and with incident cardiovascular disease: The Framingham Heart Study. J. Clin. Lipidol. 2018, 12, 718–727.e6. [Google Scholar] [CrossRef]
- Kleber, M.E.; Delgado, G.E.; Lorkowski, S.; März, W.; von Schacky, C. Omega-3 fatty acids and mortality in patients referred for coronary angiography. The Ludwigshafen Risk and Cardiovascular Health Study. Atherosclerosis 2016, 252, 175–181. [Google Scholar] [CrossRef]
- Janson, M. Rund 8 Millionen Deutsche essen kein Fleisch. Available online: https://de.statista.com/infografik/24000/anzahl-der-vegetarier-und-veganer-in-deutschland/ (accessed on 27 June 2023).
- Greupner, T.; Kutzner, L.; Nolte, F.; Strangmann, A.; Kohrs, H.; Hahn, A.; Schebb, N.H.; Schuchardt, J.P. Effects of a 12-week high-α-linolenic acid intervention on EPA and DHA concentrations in red blood cells and plasma oxylipin pattern in subjects with a low EPA and DHA status. Food Funct. 2018, 9, 1587–1600. [Google Scholar] [CrossRef]
- Wilkinson, P.; Leach, C.; Ah-Sing, E.E.; Hussain, N.; Miller, G.J.; Millward, D.J.; Griffin, B.A. Influence of α-linolenic acid and fish-oil on markers of cardiovascular risk in subjects with an atherogenic lipoprotein phenotype. Atherosclerosis 2005, 181, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Kuhnt, K.; Weiß, S.; Kiehntopf, M.; Jahreis, G. Consumption of echium oil increases EPA and DPA in blood fractions more efficiently compared to linseed oil in humans. Lipids Health Dis. 2016, 15, 32. [Google Scholar] [CrossRef] [PubMed]
- Barceló-Coblijn, G.; Murphy, E.J.; Othman, R.; Moghadasian, M.H.; Kashour, T.; Friel, J.K. Flaxseed oil and fish-oil capsule consumption alters human red blood cell n–3 fatty acid composition: A multiple-dosing trial comparing 2 sources of n–3 fatty acid. Am. J. Clin. Nutr. 2008, 88, 801–809. [Google Scholar] [CrossRef]
- Baker, E.J.; Miles, E.A.; Burdge, G.C.; Yaqoob, P.; Calder, P.C. Metabolism and functional effects of plant-derived omega-3 fatty acids in humans. Prog. Lipid Res. 2016, 64, 30–56. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T.; Salem, N.; Sinclair, A.J.; Cunnane, S.C. α-Linolenic acid supplementation and conversion to n-3 long-chain polyunsaturated fatty acids in humans. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 85–91. [Google Scholar] [CrossRef]
- Geiger, M.; Mohammed, B.S.; Sankarappa, S.; Sprecher, H. Studies to determine if rat liver contains chain-length-specific acyl-CoA 6-desaturases. Biochim. Biophys. Acta 1993, 1170, 137–142. [Google Scholar] [CrossRef]
- Morise, A.; Combe, N.; Boué, C.; Legrand, P.; Catheline, D.; Delplanque, B.; Fénart, E.; Weill, P.; Hermier, D. Dose effect of α-linolenic acid on PUFA conversion, bioavailability, and storage in the hamster. Lipids 2004, 39, 325–334. [Google Scholar] [CrossRef]
- Dawczynski, C.; Dittrich, M.; Neumann, T.; Goetze, K.; Welzel, A.; Oelzner, P.; Völker, S.; Schaible, A.; Troisi, F.; Thomas, L.; et al. Docosahexaenoic acid in the treatment of rheumatoid arthritis: A double-blind, placebo-controlled, randomized cross-over study with microalgae vs. sunflower oil. Clin. Nutr. 2018, 37, 494–504. [Google Scholar] [CrossRef]
- Escudero, A.; Montilla, J.C.; García, J.M.; Sánchez-Quevedo, M.C.; Periago, J.L.; Hortelano, P.; Suárez, M.D. Effect of dietary (n-9), (n-6) and (n-3) fatty acids on membrane lipid composition and morphology of rat erythrocytes. Biochim. Biophys. Acta 1998, 1394, 65–73. [Google Scholar] [CrossRef]
- Harris, W.S.; Masson, S.; Barlera, S.; Milani, V.; Pileggi, S.; Franzosi, M.G.; Marchioli, R.; Tognoni, G.; Tavazzi, L.; Latini, R. Red blood cell oleic acid levels reflect olive oil intake while omega-3 levels reflect fish intake and the use of omega-3 acid ethyl esters: The Gruppo Italiano per lo Studio della Sopravvivenza nell’Infarto Miocardico–Heart Failure trial. Nutr. Res. 2016, 36, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.D.; Periago, J.L.; Pita, M.L.; Hortelano, P. The n−3 polyunsaturated fatty acid levels in rat tissue lipids increase in response to dietary olive oil relative to sunflower oil. Lipids 1994, 29, 845–849. [Google Scholar] [CrossRef]
- Pfeuffer, M.; Watzl, B. Nutrition and health aspects of milk and dairy products and their ingredients. Ernaehrungs Umsch. 2018, 65, 22–33. [Google Scholar] [CrossRef]
- Fusconi, E.; Pala, V.; Riboli, E.; Vineis, P.; Sacerdote, C.; Del Pezzo, M.; de Magistris, M.S.; Palli, D.; Masala, G.; Sieri, S.; et al. Relationship between plasma fatty acid composition and diet over previous years in the Italian centers of the European Prospective Investigation into Cancer and Nutrition (EPIC). Tumori J. 2003, 89, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Welch, A.A.; Shakya-Shrestha, S.; Lentjes, M.A.; Wareham, N.J.; Khaw, K.-T. Dietary intake and status of n–3 polyunsaturated fatty acids in a population of fish-eating and non-fish-eating meat-eaters, vegetarians, and vegans and the precursor-product ratio of α-linolenic acid to long-chain n–3 polyunsaturated fatty acids: Results from the EPIC-Norfolk cohort. Am. J. Clin. Nutr. 2010, 92, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Stute, P. Veränderungen in der Menopause. Gynäkologische Endokrinol. 2011, 9, 144–150. [Google Scholar] [CrossRef]
- Rahm, J.J.; Holman, R.T. Effect of Linoleic Acid upon the Metabolism of Linolenic Acid. J. Nutr. 1964, 84, 15–19. [Google Scholar] [CrossRef]
- Gibson, R.; Neumann, M.; Lien, E.; Boyd, K.; Tu, W. Docosahexaenoic acid synthesis from alpha-linolenic acid is inhibited by diets high in polyunsaturated fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 139–146. [Google Scholar] [CrossRef]
- Smink, W.; Gerrits, W.J.; Gloaguen, M.; Ruiter, A.; van Baal, J. Linoleic and α-linolenic acid as precursor and inhibitor for the synthesis of long-chain polyunsaturated fatty acids in liver and brain of growing pigs. Animal 2012, 6, 262–270. [Google Scholar] [CrossRef]
- Blanchard, H.; Pédrono, F.; Boulier-Monthéan, N.; Catheline, D.; Rioux, V.; Legrand, P. Comparative effects of well-balanced diets enriched in α-linolenic or linoleic acids on LC-PUFA metabolism in rat tissues. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 383–389. [Google Scholar] [CrossRef]
- Goyens, P.L.L.; Spilker, M.E.; Zock, P.L.; Katan, M.B.; Mensink, R.P. Conversion of α-linolenic acid in humans is influenced by the absolute amounts of α-linolenic acid and linoleic acid in the diet and not by their ratio. Am. J. Clin. Nutr. 2006, 84, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Rosell, M.S.; Lloyd-Wright, Z.; Appleby, P.N.; AB Sanders, T.; E Allen, N.; Key, T.J. Long-chain n–3 polyunsaturated fatty acids in plasma in British meat-eating, vegetarian, and vegan men. Am. J. Clin. Nutr. 2005, 82, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Emken, E.A.; Adlof, R.O.; Gulley, R.M. Dietary linoleic acid influences desaturation and acylation of deuterium-labeled lino-leic and linolenic acids in young adult males. Biochim. Biophys. Acta 1994, 1213, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Jones, A.E.; Wootton, S.A. Eicosapentaenoic and docosapentaenoic acids are the principal products of α-linolenic acid metabolism in young men. Br. J. Nutr. 2002, 88, 355–363. [Google Scholar] [CrossRef]
- Burdge, G.C.; Wootton, S.A. Conversion of α-linolenic acid to eicosapentaenoic, docosapentaenoic and docosahexaenoic acids in young women. Br. J. Nutr. 2002, 88, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Pawlosky, R.J.; Hibbeln, J.R.; Novotny, J.A.; Salem, N. Physiological compartmental analysis of α-linolenic acid metabolism in adult humans. J. Lipid Res. 2001, 42, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Shen, J.; Abecasis, G.R.; Kisialiou, A.; Ordovas, J.M.; Guralnik, J.M.; Singleton, A.; Bandinelli, S.; Cherubini, A.; Arnett, D.; et al. Genome-Wide Association Study of Plasma Polyunsaturated Fatty Acids in the InCHIANTI Study. PLoS Genet. 2009, 5, e1000338. [Google Scholar] [CrossRef]
- Matsuzaka, T.; Shimano, H.; Yahagi, N.; Amemiya-Kudo, M.; Yoshikawa, T.; Hasty, A.H.; Tamura, Y.; Osuga, J.; Okazaki, H.; Iizuka, Y.; et al. Dual regulation of mouse Δ5- and Δ6-desaturase gene expression by SREBP-1 and PPARα. J. Lipid Res. 2002, 43, 107–114. [Google Scholar] [CrossRef]
- Brenner, R.R. Nutritional and hormonal factors influencing desaturation of essential fatty acids. Prog. Lipid Res. 1981, 20, 41–47. [Google Scholar] [CrossRef]
- Gonzalez-Soto, M.; Mutch, D.M. Diet Regulation of Long-Chain PUFA Synthesis: Role of Macronutrients, Micronutrients, and Polyphenols on Δ-5/Δ-6 Desaturases and Elongases 2/5. Adv. Nutr. Int. Rev. J. 2021, 12, 980–994. [Google Scholar] [CrossRef]
- Chiavaroli, L.; Nishi, S.K.; Khan, T.A.; Braunstein, C.R.; Glenn, A.J.; Mejia, S.B.; Rahelić, D.; Kahleová, H.; Salas-Salvadó, J.; Jenkins, D.J.; et al. Portfolio Dietary Pattern and Cardiovascular Disease: A Systematic Review and Meta-analysis of Controlled Trials. Prog. Cardiovasc. Dis. 2018, 61, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Yu-Poth, S.; Zhao, G.; Etherton, T.; Naglak, M.; Jonnalagadda, S.; Kris-Etherton, P.M. Effects of the National Cholesterol Education Program’s Step I and Step II dietary intervention programs on cardiovascular disease risk factors: A meta-analysis. Am. J. Clin. Nutr. 1999, 69, 632–646. [Google Scholar] [CrossRef] [PubMed]
- Stone, N.J.; Nicolosi, R.J.; Kris-Etherton, P.; Ernst, N.D.; Krauss, R.M.; Winston, M. Summary of the Scientific Conference on the Efficacy of Hypocholesterolemic Dietary Interventions. Circulation 1996, 94, 3388–3391. [Google Scholar] [CrossRef] [PubMed]
- Rosa, G.; Avelino, A.P.A.; de Oliveira, G.M.M.; Luiz, R.R.; Ferreira, C.C.D. Additive effect of linseed oil supplementation on the lipid profiles of older adults. Clin. Interv. Aging 2015, 10, 1679–1685. [Google Scholar] [CrossRef]
- Paschos, G.K.; Magkos, F.; Panagiotakos, D.B.; Votteas, V.; Zampelas, A. Dietary supplementation with flaxseed oil lowers blood pressure in dyslipidaemic patients. Eur. J. Clin. Nutr. 2007, 61, 1201–1206. [Google Scholar] [CrossRef]
- Mirmasoumi, G.; Fazilati, M.; Foroozanfard, F.; Vahedpoor, Z.; Mahmoodi, S.; Taghizadeh, M.; Esfeh, N.K.; Mohseni, M.; Karbassizadeh, H.; Asemi, Z. The Effects of Flaxseed Oil Omega-3 Fatty Acids Supplementation on Metabolic Status of Patients with Polycystic Ovary Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial. Exp. Clin. Endocrinol. Diabetes 2018, 126, 222–228. [Google Scholar] [CrossRef]
- Wendland, E.; Farmer, A.; Glasziou, P.; Neil, A. Effect of linolenic acid on cardiovascular risk markers: A systematic review. Heart 2006, 92, 166–169. [Google Scholar] [CrossRef]
- De Abreu, A.M.; Copetti, C.L.K.; Hauschild, D.B.; Di Pietro, P.F.; Wazlawik, E. Effects of supplementation with vegetable sources of alpha-linolenic acid (ALA) on inflammatory markers and lipid profile in individuals with chronic kidney disease: A systematic review and meta-analysis. Clin. Nutr. 2022, 41, 1434–1444. [Google Scholar] [CrossRef]
- Ristic-Medic, D.; Perunicic-Pekovic, G.; Rasic-Milutinovic, Z.; Takic, M.; Popovic, T.; Arsic, A.; Glibetic, M. Effects of Dietary Milled Seed Mixture on Fatty Acid Status and Inflammatory Markers in Patients on Hemodialysis. Sci. World J. 2014, 2014, 563576. [Google Scholar] [CrossRef]
- Soltani, S.K.; Jamaluddin, R.; Tabibi, H.; Yusof, B.N.M.; Atabak, S.; Loh, S.; Rahmani, L. Effects of flaxseed consumption on systemic inflammation and serum lipid profile in hemodialysis patients with lipid abnormalities. Hemodial. Int. 2013, 17, 275–281. [Google Scholar] [CrossRef]
- Weikert, C.; Trefflich, I.; Menzel, J.; Obeid, R.; Longree, A.; Dierkes, J.; Meyer, K.; Herter-Aeberli, I.; Mai, K.; Stangl, G.I.; et al. Vitamin and Mineral Status in a Vegan Diet. Dtsch. Aerzteblatt Int. 2020, 117, 575–582. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid Distribution of Linseed Oil (% FAME) | |||||||
|---|---|---|---|---|---|---|---|
| C-16:0 | C-18:0 | C-18:1c9 | C-18:2c9, c12 | aC-18:3c9, c12, c15 | SFA | MUFA | PUFA |
| 4.6 | 2.9 | 17.5 | 13.7 | 60.2 | 7.8 | 18.3 | 74.0 |
| Characteristics of the Study Collective 69 w (65.7%); 36 m (34.3%) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Age (Years) | BMI (kg/m2) | Total Cholesterol (mmol/L) | LDL Cholesterol (mmol/L) | HDL Cholesterol (mmol/L) | Triglycerides (mmol/L) | Lipoprotein(a) (mg/L) | Systolic Blood Pressure (mmHG) | Diastolic Blood Pressure (mmHG) |
| 57.4 (±12.1) | 25.6 (23.4, 29.1) | 5.8 (±1.2) | 3.6 (±1.0) | 1.6 (±1.0) | 1.1 (0.9, 1.5) | 88.5 (40.3, 264.5) | 140.5 (±21.1) | 86.9 (±11.0) |
| High Linoleic Acid 15 w (55.6%) 12 m (44.4%) | Low Linoleic Acid 19 w (70.4%) 8 m (29.6%) | High Milk 17 w (73.9%) 6 m (26.1%) | Control 18 w (65.7%) 10 m (34.3%) | |||||
|---|---|---|---|---|---|---|---|---|
| Age (years) | 57.7 (±11.5) | a | 56.8 (±11.6) | a | 55.8 (±12.3) | a | 58.9 (±13.4) | a |
| Subgroups | High Linoleic Acid | Low Linoleic Acid | High Milk | Control | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cfb (%) * | ∆ | ◊ | n | Cfb (%) * | ∆ | ◊ | n | Cfb (%) * | ∆ | ◊ | n | Cfb (%) * | ∆ | ◊ | n | |
| aC-18:3c9c12c15 (ALA) | ||||||||||||||||
| Women | 440.07 (±235.72) | a | 15 | 355.28 (±246.75) | a | 19 | 389.05 (±267.22) | a | 17 | 300.40 (±174.32) | a | 17 | ||||
| Men | 496.34 (±349.31) | n.s. | a | 12 | 352.12 (±171.60) | n.s. | a | 8 | 341.87 (±207.12) | n.s. | a | 5 | 288.55 (±132.89) | n.s. | a | 9 |
| EPA (<0.9)Baseline | 463.30 (±264.72) 511.27 (206.41, 700.75) | a | 14 | 310.82 (±217.39) | a | 13 | 407.49 (±245.47) | a | 14 | 295.96 (±136.85) | a | 12 | ||||
| EPA (≥0.9)Baseline | 350.23 (263.60, 557.27) | n.s. | a | 13 | 381.07 (±233.81) 404.86 (179.94, 579.31) | n.s. | a | 14 | 350.20 (±268.10) 395.00 (112.72, 495.26) | n.s. | a | 8 | 278.03 (±173.12) 207.75 (166.42, 364.09) | n.s. | a | 13 |
| LA (<1.5)Change | 512.00 (±271.13) 532.77 (424.35, 620.47) | a | 4 | 301.76 (±198.13) 386.47 (119.93, 464.86) | a | 7 | 354.17 (±227.09) 343.57 (162.18, 524.13) | a | 20 | 261.63 (±164.96) 265.22 (187.96, 324.90) | a | 20 | ||||
| LA (≥1.5)Change | 399.88 (254.99, 699.00) | n.s. | a | 23 | 429.40 (242.02, 562.22) | n.s | a | 20 | 619.88 (456.14, 783.63) | n.s | a | 2 | 446.61 (288.99, 464.50) | 0.037 | a | 6 |
| C-20:4n3 (ETA) | ||||||||||||||||
| Women | 81.12 (±110.66) | a,b | 15 | 60.27 (±87.56) | a | 19 | 156.49 (±134.89) | b | 17 | 92.94 (±61.36) | a,b | 14 | ||||
| Men | 108.16 (±97.61) | n.s. | a | 12 | 113.86 (±68.37) | n.s. | a | 8 | 188.56 (±63.19) | n.s. | a | 5 | 100.29 (±95.39) | n.s. | a | 8 |
| EPA (<0.9)Baseline | 135.20 (±104.60) 138.04 (61.09, 225.49) | a | 14 | 114.61 (±89.01) | a | 13 | 182.42 (±133.39) | a | 14 | 91.13 (±55.75) | a | 8 | ||||
| EPA (≥0.9)Baseline | 30.02 (−6.89, 80.63) | 0.025 | a | 13 | 33.65 (±57.69) 28.81 (−11.25, 65.11) | 0.011 | a | 14 | 131.16 (±97.40) 120.49 (65.12, 205.27) | n.s. | b | 8 | 109.17 (±83.16) 83.58 (53.36, 129.30) | n.s. | b | 12 |
| LA (<1.5)Change | 137.34 (±78.69) 117.32 (78.86, 175.80) | a | 4 | 135.06 (±100.98) 165.04 (43.66, 202.61) | a | 7 | 154.90 (±112.80) 140.21 (92.79, 205.27) | a | 20 | 57.78 (±52.16) 62.71 (27.46, 82.67) | a | 18 | ||||
| LA (≥1.5)Change | 47.17 (7.69, 142.03) | n.s | a | 23 | 37.90 (9.27, 107.54) | 0.030 | a | 20 | 252.59 (171.98, 333.19) | n.s | a | 2 | 115.20 (85.39, 141.34) | n.s | a | 4 |
| C-20:5n3 (EPA) | ||||||||||||||||
| Women | 59.81 (24.86, 76.67) | a | 15 | 51.62 (±49.10) 28.13 (16.47, 84.55) | a | 19 | 68.98 (±75.86) 58.03 (13.06, 108.47) | a | 17 | 32.47 (±37.81) 29.83 (8.61, 42.52) | a | 17 | ||||
| Men | 79.40 (±76.71) 73.35 (29.75, 101.59) | n.s. | a | 12 | 64.33 (±26.56) | n.s. | a | 8 | 92.70 (±25.56) | n.s. | a | 5 | 66.83 (±64.39) | n.s. | a | 10 |
| EPA (<0.9)Baseline | 107.50 (±78.73) | a | 14 | 74.00 (±41.07) | a | 13 | 98.73 (±68.92) | a | 14 | 56.71 (±45.75) 68.44 (23.32, 101.10) | a | 13 | ||||
| EPA (≥0.9)Baseline | 32.10 (±35.28) 28.96 (−3.47, 68.84) | 0.004 | a | 13 | 38.53 (±39.89) 22.22 (14.61, 68.02) | 0.032 | a | 14 | 34.02 (±43.98) 26.22 (8.96, 59.44) | 0.027 | a | 8 | 32.43 (11.06, 39.89) | n.s. | a | 14 |
| LA (<1.5)Change | 117.27 (±105.52) 79.58 (61.35, 135.50) | a | 4 | 67.79 (±32.73) 75.83 (47.03, 87.31) | a | 7 | 65.75 (±59.93) 60.72 (27.77, 106.44) | a | 20 | 30.92 (±44.66) 29.79 (4.67, 53.14) | a | 21 | ||||
| LA (≥1.5)Change | 59.81 (24.86, 79.28) | n.s | a | 23 | 50.25 (15.69, 81.12) | n.s | a | 20 | 160.59 (121.44, 199.75) | n.s | a | 2 | 26.57 (19.64, 35.34) | n.s | a | 6 |
| C-22:5n3 (DPA) | ||||||||||||||||
| Women | −3.02 (±26.18) | a | 15 | −4.82 (±23.00) | a | 19 | 34.26 (±49.65) | b | 17 | 8.89 (±15.43) | a | 17 | ||||
| Men | −1.40 (±19.34) | n.s. | a | 12 | 17.56 (±16.58) | 0.020 | b | 8 | 41.11 (±11.51) | n.s. | c | 5 | 15.31 (±13.64) | n.s. | b | 10 |
| EPA (<0.9)Baseline | 2.68 (±26.35) | a | 14 | 11.77 (±20.35) | a,b | 13 | 40.90 (±51.54) | b | 14 | 13.75 (±16.20) | a,b | 13 | ||||
| EPA (≥0.9)Baseline | −7.66 (±18.21) | n.s. | a | 13 | −7.55 (±22.80) | 0.029 | a | 14 | 28.32 (±29.17) | n.s. | b | 8 | 8.97 (±13.69) | n.s. | a,b | 14 |
| LA (<1.5)Change | 13.94 (±35.11) 17.52 (−4.90, 36.36) | a | 4 | 17.54 (±19.96) 24.32 (12.17, 29.05) | a | 7 | 33.73 (±43.23) 35.27 (8.32, 48.14) | a | 20 | 8.69 (±14.26) 8.74 (−4.03, 20.06) | a | 21 | ||||
| LA (≥1.5)Change | −12.91 (−16.82, 12.83) | n.s | a | 23 | −6.99 (−15.93, 6.69) | 0.036 | a | 20 | 56.72 (35.29, 78.16) | n.s | a,b | 2 | 17.39 (10.91, 33.93) | 0.026 | b | 6 |
| C-22:6n3 (DHA) | ||||||||||||||||
| Women | −27.60 (±19.16) | a | 15 | −32.59 (±14.65) | a | 19 | 2.80 (±32.21) | b | 17 | −20.72 (±13.63) | a | 17 | ||||
| Men | −32.70 (±12.76) | n.s. | a | 12 | −26.88 (±9.38) | n.s. | a,b | 8 | −4.30 (±24.92) | n.s. | c | 5 | −15.31 (±11.67) | n.s. | b,c | 10 |
| EPA (<0.9)Baseline | −25.38 (±19.61) | a | 14 | −24.23 (±13.19) | a | 13 | 11.94 (±31.47) | b | 14 | −20.32 (±13.72) | a | 13 | ||||
| EPA (≥0.9)Baseline | −34.70 (±11.27) | n.s. | a | 13 | −36.77 (±11.06) | 0.013 | a | 14 | −18.63 (±14.67) | 0.018 | b | 8 | −17.23 (±12.58) | n.s. | b | 14 |
| LA (<1.5)Change | −10.49 (−34.11, 8.57) | a | 4 | −17.25 (−23.05, −14.31) | a | 7 | −7.01 (−22.77, 17.87) | a | 20 | −14.00 (−23.35, −11.72) | a | 21 | ||||
| LA (≥1.5)Change | −34.10 (−41.68, −20.77) | n.s | a | 23 | −37.03 (−40.61, −29.90) | 0.007 | a | 20 | 5.88 (1.37, 10.39) | n.s | a | 2 | −21.70 (−35.15, −12.34) | n.s | a | 6 |
| n-3 index | ||||||||||||||||
| Women | −25.95 (−28.58, −9.96) | a | 15 | −20.49 (±15.70) −23.91 (−32.02, −6.83) | a | 19 | 9.81 (±27.95) −0.04 (−9.27, 29.80) | b | 17 | −14.12 (−21.15, −3.20) | a,b | 17 | ||||
| Men | −18.16 (−23.46, −13.09) | n.s. | a | 12 | −12.05 (±8.78) −12.23 (−16.09, −6.14) | n.s. | a,b | 8 | 11.82 (±22.98) 18.58 (−6.18, 18.77) | n.s. | b | 5 | −4.70 (−9.67, −2.17) | n.s. | a,b | 10 |
| EPA (<0.9)Baseline | −9.17 (±26.00) | a | 14 | −9.96 (±13.12) | a | 13 | 21.34 (±25.26) | b | 14 | −9.34 (±16.90) −5.73 (−28.66, 1.94) | a | 13 | ||||
| EPA (≥0.9)Baseline | −23.89 (±9.93) −25.95 (−28.85, −16.77) | n.s. | a | 13 | −25.34 (±11.58) −26.46 (−32.31, −21.42) | 0.003 | a | 14 | −9.38 (±14.49) −11.42 (−15.40, −6.71) | 0.002 | b | 8 | −12.20 (−16.88, −4.22) | n.s. | b | 14 |
| LA (<1.5)Change | 4.48 (±45.02) 3.27 (−25.29, 33.03) | a | 4 | −6.67 (±10.24) −5.72 (−11.24, −1.16) | a | 7 | 8.53 (±26.35) −3.11 (−9.58, 25.31) | a | 20 | −9.58 (±13.30) −9.61 (−13.93, −0.63) | a | 21 | ||||
| LA (≥1.5)Change | −20.12 (−28.56, −13.27) | n.s | a | 23 | −24.01 (−31.72, −13.14) | 0.012 | a | 20 | 27.61 (17.80, 37.41) | n.s | a | 2 | −18.65 (−28.86, −5.49) | n.s | a | 6 |
| C-18:2c9c12 (LA) | ||||||||||||||||
| Women | 28.30 (±11.89) 29.78 (23.63, 35.54) | a | 15 | 20.70 (±13.85) 21.83 (12.02, 31.19) | a | 19 | 4.19 (−3.48, 6.35) | b | 17 | 6.14 (−0.14, 10.64) | b | 17 | ||||
| Men | 26.12 (±8.28) 26.92 (20.88, 31.42) | n.s. | a | 12 | 20.44 (±7.25) 20.29 (16.21, 23.04) | n.s. | a,b | 8 | 9.54 (4.54, 9.81) | n.s. | c | 5 | 9.13 (5.51, 13.36) | n.s. | b,c | 10 |
| EPA (<0.9)Baseline | 23.00 (±10.56) 23.64 (14.28, 28.44) | a | 14 | 15.95 (±12.08) 15.53 (10.80, 21.73) | a,b | 13 | 5.55 (2.56, 8.83) | c | 14 | 9.50 (5.40, 14.12) | b,c | 13 | ||||
| EPA (≥0.9)Baseline | 32.00 (±8.02) 31.91 (27.30, 36.31) | 0.020 | a | 13 | 24.96 (±10.82) 23.87 (18.37, 30.90) | n.s. | a | 14 | 3.39 (−1.00, 6.43) | n.s. | b | 8 | 5.98 (−2.99, 9.48) | n.s. | b | 14 |
| LA (<1.5)Change | 10.16 (8.76, 11.71) | a | 4 | 9.31 (2.39, 10.47) | a | 7 | 4.36 (−2.43, 6.44) | a | 20 | 5.52 (−0.14, 8.76) | a | 21 | ||||
| LA (≥1.5)Change | 29.78 (24.88, 34.85) | <0.001 | a | 23 | 23.87 (18.44, 32.60) | <0.001 | a | 20 | 20.66 (17.36, 23.96) | 0.009 | a | 2 | 21.65 (19.07, 26.39) | <0.001 | a | 6 |
| C-20:4c5c8c11c14 (ARA) | ||||||||||||||||
| Women | −14.69 (±7.57) −16.22 (−21.35, −10.14) | a | 15 | −12.82 (±8.43) −12.92 (−18.09, −6.52) | a | 19 | −7.08 (−14.83, 6.22) | a | 17 | −14.26 (±7.52) −14.16 (−19.13, −7.68) | a | 17 | ||||
| Men | −13.90 (±8.32) | n.s. | a | 12 | −9.43 (±7.14) | n.s. | a | 8 | −1.31 (±4.31) −2.33 (−2.51, 0.50) | n.s. | a | 5 | −13.22 (±19.52) | n.s. | a | 10 |
| EPA (<0.9)Baseline | −16.43 (±7.51) −17.43 (−20.90, −12.03) | a | 14 | −10.85 (±7.71) −10.01 (−16.64, −6.36) | a,b | 13 | −4.64 (−11.00, 5.92) | b | 14 | −13.43 (±5.44) −13.52 (−16.75, −11.18) | a,b | 13 | ||||
| EPA (≥0.9)Baseline | −12.10 (±7.67) | n.s. | a | 13 | −12.67 (±8.62) | n.s. | a | 14 | −7.40 (±9.07) −4.80 (−14.65, −1.76) | n.s. | a | 8 | −14.28 (±17.50) | n.s. | a | 14 |
| LA (<1.5)Change | −16.58 (−18.54, −12.14) | a | 4 | −6.53 (−9.43, −5.95) | a | 7 | −4.80 (−13.84, 4.81) | a | 20 | −13.52 (−16.12, −9.42) | a | 21 | ||||
| LA (≥1.5)Change | −13.99 (−20.20, −9.14) | n.s | a | 23 | −14.49 (−18.03, −9.29) | n.s | a | 20 | 0.17 (−5.15, 5.48) | n.s | a | 2 | −16.06 (−21.75, −8.66) | n.s | a | 6 |
| C-22:4n6 | ||||||||||||||||
| Women | −27.13 (±14.12) | a | 15 | −25.63 (±16.45) | a | 19 | −3.74 (±25.66) | b | 17 | −11.18 (±14.77) | b | 17 | ||||
| Men | −27.72 (±18.55) | n.s. | a | 12 | −12.71 (13.33) | n.s. | a,b | 8 | 0.62 (±7.60) | n.s. | b | 5 | −13.71 (±15.47) | n.s. | a,b | 10 |
| EPA (<0.9)Baseline | −29.03 (±15.90) | a | 14 | −15.83 (±13.95) | a,b | 13 | −4.33 (±23.23) −1.97 (−20.59, 12.07) | b | 14 | −10.07 (±7.66) | b | 13 | ||||
| EPA (≥0.9)Baseline | −25.63 (±16.36) −30.21 (−37.30, −14.59) | n.s. | a | 13 | −27.39 (±17.12) −31.58 (−40.73, −17.35) | n.s. | a | 14 | 2.95 (±30.14) −3.21 (−18.16, 7.33) | n.s. | b | 8 | −14.01 (±19.36) −22.27 (−12.38, 0.63) | n.s. | a,b | 14 |
| LA (<1.5)Change | −38.80 (−70.56, −23.11) | a | 4 | −12.17 (−14.32, −4.42) | a | 7 | −2.00 (−24.30, 12.16) | a | 20 | −13.96 (−22.16, −4.27) | a | 21 | ||||
| LA (≥1.5)Change | −43.28 (−63.97, −15.25) | n.s | a | 23 | −35.16 (−59.88, −20.40) | n.s | a | 20 | 1.55 (−1.35, 4.44) | n.s | a | 2 | −3.30 (−63.92, −1.54) | n.s | a | 6 |
| C-22:5n6 | ||||||||||||||||
| Women | −43.98 (±13.90) −48.41 (−54.91, −37.67) | a | 15 | −42.01 (±17.11) | a | 19 | −16.81 (±29.00) | b | 17 | −29.37 (±21.93) | a,b | 17 | ||||
| Men | −45.43 (−59.05, −37.93) | n.s. | a | 12 | −32.65 (±14.02) −36.72 (−38.57, −28.21) | n.s. | a,b | 8 | −13.14 (±6.21) −10.99 (−14.46, −10.00) | n.s. | b | 5 | −26.06 (±33.16) −30.96 (−46.41, −21.32) | n.s. | a,b | 10 |
| EPA (<0.9)Baseline | −46.52 (±14.72) −49.12 (−59.09, −35.55) | a | 14 | −32.92 (±14.41) | a,b | 13 | −18.83 (±28.62) −31.14 (−14.37, −0.65) | b | 14 | −35.75 (±19.14) | a,b | 13 | ||||
| EPA (≥0.9)Baseline | −43.48 (−54.67, −39.87) | n.s. | a,b | 13 | −44.21 (±17.51) −46.05 (−56.95, −38.57) | n.s. | a | 14 | −16.59 (−24.20, 0.53) | n.s. | c | 8 | −21.09 (±30.15) −32.05 (−40.23, −1.28) | n.s. | b,c | 14 |
| LA (<1.5)Change | −45.96 (−52.19, −35.88) | a | 4 | −28.67 (−34.70, −21.42) | a | 7 | −12.72 (−26.63, 1.94) | a | 20 | −29.23 (−38.88, −15.85) | a | 21 | ||||
| LA (≥1.5)Change | −46.72 (−56.48, −36.62) | n.s | a | 23 | −43.60 (−52.52, −38.01) | n.s. | a | 20 | −23.46 (−28.04, −18.87) | n.s | a | 2 | −59.63 (−65.10, −44.86) | 0.012 | a | 6 |
| n-6/n-3 | ||||||||||||||||
| Women | 5.90 (±23.85) | a,b | 15 | 7.70 (±19.78) | a | 19 | −20.72 (±15.89) | c | 17 | −5.98 (±14.48) −6.19 (−17.84, 3.56) | b | 17 | ||||
| Men | 2.87 (±16.60) 1.79 (−11.20, 15.65) | n.s. | a | 12 | −4.47 (±9.50) −5.35 (−7.04, −1.29) | n.s. | a,b | 8 | −20.38 (±9.71) −23.95 (−27.34, −15.85) | n.s. | b | 5 | −10.87 (−16.16, −7.78) | n.s. | a,b | 10 |
| EPA (<0.9)Baseline | −4.13 (±20.01) | a | 14 | −5.38 (±14.45) | a | 13 | −27.40 (±12.05) | b | 14 | −7.04 (±15.07) | a | 13 | ||||
| EPA (≥0.9)Baseline | 13.90 (±17.46) | 0.020 | a | 13 | 12.88 (±17.00) | 0.006 | a | 14 | −9.30 (±11.52) | 0.003 | b | 8 | −10.18 (±12.51) | n.s. | b | 14 |
| LA (<1.5)Change | −14.27 (±26.68) −13.91 (−31.32, 3.15) | a | 4 | −11.21 (±10.68) −11.00 (−17.77, −4.97) | a | 7 | −20.09 (±14.73) −16.59 (−29.14, 11.63) | a | 20 | −7.77 (±15.27) −12.26 (−17.38, −1.12) | a | 21 | ||||
| LA (≥1.5)Change | 6.63 (−4.99, 18.75) | 0.046 | a | 23 | 8.21 (−3.05, 19.21) | 0.007 | a | 20 | −26.11 (−31.47, −20.75) | n.s | a | 2 | −3.55 (−10.08, 1.28) | n.s | a | 6 |
| Biochemical Parameters and Anthropometric Measurements | Week | High Linoleic Acid | Low Linoleic Acid | High Milk | Control | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Characteristics * | p ∆ | ◊ | Characteristics * | p ∆ | ◊ | Characteristics * | p ∆ | ◊ | Characteristics * | p ∆ | ◊ | |||
| Total cholesterol (mmol/L) (n † = 26, 27, 23, 28) | 0 | 5.70 (±1.36) | a | 6.07 (±0.94) | a | 5.84 (±1.24) | a | 5.76 (±1.07) | a | |||||
| 12 | 5.65 (±0.93) | n.s. | a | 5.65 (±1.06) | 0.001 | a | 5.84 (±1.27) | n.s. | a | 5.65 (±1.14) | n.s. | a | ||
| Cfb (%) | −0.88 (−8.42, 5.37) | a | −6.45 (−13.68, −2.57) | b | 0.99 (−3.97, 6.34) | a | −3.73 (−7.44, 3.24) | a,b | ||||||
| LDL cholesterol (mmol/L) (n † = 26, 27, 23, 28) | 0 | 3.57 (±1.07) | a | 3.77 (±0.73) | a | 3.58 (±1.12) | a | 3.50 (±0.89) | a | |||||
| 12 | 3.65 (±0.76) | n.s. | a | 3.57 (±0.86) | 0.029 | a | 3.67 (±1.15) | n.s. | a | 3.52 (±0.96) | n.s. | a | ||
| Cfb (%) | 1.95 (−7.19, 12.4) | a | −5.18 (−13.62, −0.85) | b | 2.35 (−6.20, 12.82) | a | −1.67 (−7.73, 6.61) | a,b | ||||||
| MDA-LDL cholesterol (U/L) (n † = 27, 27, 23, 28) | 0 | 67.20 (57.60, 92.75) | a | 68.00 (55.60, 106.50) | a | 72.24 (±22.35) 70.20 (60.15. 90.30) | a | 72.75 (±20.82) 70.20 (60.70, 85.28) | a | |||||
| 12 | 80.91 (±30.78) 77.30 (64.25, 97.55) | n.s. | a | 70.78 (±22.26) 67.70 (56.50, 86.60) | n.s. | a | 73.38 (±30.77) | n.s. | a | 76.22 (±32.57) | n.s. | a | ||
| Cfb (%) | 2.09 (−26.68, 39.31) | a | −15.60 (−25.04, 6.29) | a | 9.46 (−14.67, 15.52) | a | 2.66 (−13.30, 23.54) | a | ||||||
| HDL cholesterol (mmol/L) (n † = 26, 27, 23, 28) | 0 | 1.50 (±0.33) 1.52 (1.23, 1.75) | a | 1.62 (±0.35) 1.55 (1.41, 1.88) | a | 1.56 (1.40, 1.70) | a | 1.62 (±0.34) 1.59 (1.42, 1.83) | a | |||||
| 12 | 1.46 (±0.32) | n.s. | a | 1.56 (±0.32) | n.s. | a | 1.65 (±0.43) 1.57 (1.38, 1.73) | n.s. | a | 1.60 (±0.35) | n.s. | a | ||
| Cfb (%) | −4.07 (−5.88, 4.04) | a,b | −5.71 (−10.56, 2.31) | a | 3.73 (−1.67, 7.60) | b | −0.55 (−5.91, 3.22) | a,b | ||||||
| Non-HDL cholesterol (mmol/L) (n † = 26, 27, 23, 28) | 0 | 4.20 (±1.24) | a | 4.45 (±0.92) | a | 4.23 (±1.22) | a | 4.14 (±1.04) | a | |||||
| 12 | 4.18 (±0.86) | n.s. | a | 4.09 (±1.04) | 0.001 | a | 4.19 (1.27) | n.s. | a | 4.05 (±1.09) | n.s. | a | ||
| Cfb (%) | −0.83 (−8.97, 6.89) | a | −7.59 (−14.63, −3.69) | a | 0.32 (−6.70, 6.76) | a | −4.47 (−9.80, 4.83) | a | ||||||
| LDL/HDL (mmol/L) (n † = 26, 27, 23, 28) | 0 | 2.46 (±0.76) 2.57 (1.94, 2.71) | a | 2.30 (1.86, 2.79) | a | 2.40 (±1.02) 2.17 (1.65, 3.27) | a | 2.26 (±0.76) 2.13 (1.72, 2.71) | a | |||||
| 12 | 2.61 (±0.77) 2.46 (1.97, 3.23) | n.s. | a | 2.20 (1.81,2.74) | n.s. | a | 2.39 (±1.03) 2.19 (1.56, 3.21) | n.s. | a | 2.31 (±0.84) 2.34 (1.57, 2.83) | n.s. | a | ||
| Cfb (%) | 3.63 (−4.06, 11.25) | a | −2.66 (−6.38, 5.12) | a | −1.33 (−9.21, 6.98) | a | −0.01 (−7.95, 11.95) | a | ||||||
| Triglycerides (mmol/L) (n † = 26, 27, 23, 28) | 0 | 1.21 (1.01, 1.33) | a | 1.09 (0.92, 1.67) | a | 0.93 (0.77, 1.18) | a | 1.00 (0.90, 1.36) | a | |||||
| 12 | 1.27 (1.07, 1.67) | n.s. | a | 1.01 (0.90, 1.63) | n.s. | a | 1.19 (0.88, 1.35) | n.s. | a | 1.16 (0.79, 1.46) | n.s. | a | ||
| Cfb (%) | 12.71 (−13.13, 34.61) | a | −5.85 (−14.95, 15.83) | a | 14.02 (−5.58, 36.85) | a | 1.63 (−16.68, 42.64) | a | ||||||
| High-sensitivity CRP (mg/L) (n † = 26, 27, 23, 28) | 0 | 1.20 (0.60, 1.90) | a | 1.10 (0.75, 2.70) | a | 1.70 (1.00, 2.90) | a | 0.85 (0.40, 1.93) | a | |||||
| 12 | 1.35 (0.50, 2.58) | n.s. | a | 0.90 (0.40, 2.30) | n.s. | a | 1.30 (0.65, 2.05) | 0.031 | a | 0.85 (0.40, 1.93) | n.s. | a | ||
| Cfb (%) | −1.79 (−24.74, 0.00) | a | −16.67 (−36.04, 2.50) | a | −21.43 (−50.00, 12.04) | a | 0.00 (−25.75, 20.36) | a | ||||||
| Apolipoprotein B/apolipoprotein A1 (n † = 27, 27, 23, 28) | 0 | 0.71 (±0.21) 0.71 (0.58, 0.77) | a | 0.64 (0.55, 0.80) | a | 0.71 (±0.26) 0.63 (0.50, 0.90) | a | 0.66 (±0.18) 0.64 (0.55, 0.76) | a | |||||
| 12 | 0.78 (±0.23) 0.75 (0.63, 1.00) | 0.004 | a | 0.66 (0.53, 0.80) | n.s. | a | 0.72 (±0.27) 0.66 (0.52, 0.91) | n.s. | a | 0.70 (±0.21) 0.68 (0.56, 0.83) | 0.022 | a | ||
| Cfb (%) | 7.61 (2.78, 12.59) | a | 2.00 (−6.64, 9.05) | a | 1.59 (−5.88, 10.40) | a | 1.95 (−1.17, 10.01) | a | ||||||
| Apolipoprotein A1 (g/L) (n † = 27, 27, 23, 28) | 0 | 1.65 (±0.28) | a | 1.72 (±0.26) | a | 1.66 (±0.32) 1.66 (1.49, 1.81) | a | 1.68 (±0.24) | a | |||||
| 12 | 1.53 (±0.29) 1.47 (1.33, 1.76) | <0.001 | a | 1.60 (±0.25) 1.53 (1.47, 1.77) | 0.001 | a | 1.60 (1.38, 1.79) | n.s. | a | 1.60 (±0.25) 1.57 (1.46, 1.68) | 0.001 | a | ||
| Cfb (%) | −7.33 (±6.34) | a | −6.61 (±9.10) | a | −1.84 (±8.37) | a | −4.89 (±6.70) | a | ||||||
| Apolipoprotein B (g/L) (n † = 27, 27, 23, 28) | 0 | 1.04 (0.94, 1.27) | a | 1.18 (±0.27) 1.14 (0.97, 1.35) | a | 1.13 (±0.31) 1.08 (0.86, 1.32) | a | 1.10 (±0.26) 1.14 (0.90, 1.25) | a | |||||
| 12 | 1.15 (±0.25) 1.12 (0.95, 1.37) | n.s. | a | 1.11 (±0.29) | 0.027 | a | 1.13 (±0.31) | n.s. | a | 1.09 (±0.28) | n.s. | a | ||
| Cfb (%) | −0.96 (−7.08, 6.72) | a | −6.11 (−11.80, 0.00) | a | 0.00 (−6.35, 6.40) | a | −0.91 (−8.14, 3.62) | a | ||||||
| Lipoprotein(a) (mg/L) (n † = 26, 27, 23, 28) | 0 | 109.50 (51.00, 304.00) | a | 65.00 (30.50, 263.50) | a | 87.00 (51.00, 265.50) | a | 86.00 (43.00, 206.25) | a | |||||
| 12 | 116.50 (64.00, 351.50) | 0.002 | a | 67.00 (45.50, 325.50) | 0.002 | a | 92.00 (50.00, 288.00) | n.s. | a | 116.50 (55.75, 234.00) | n.s. | a | ||
| Cfb (%) | 16.21 (−1.76, 35.35) | a | 19.05 (5.25, 35.37) | a | 7.48 (−1.52, 23.40) | a | 11.53 (−6.80, 27.04) | a | ||||||
| Homocysteine (µmol/L) (n † = 27, 27, 23, 28) | 0 | 10.70 (8.65, 12.80) | a | 9.45 (±1.99) 9.90 (7.70, 10.40) | a | 8.70 (8.15, 9.90) | a | 9.30 (7.87, 11.93) | a | |||||
| 12 | 9.70 (8.35, 11.30) | n.s. | a | 10.06 (±2.17) 10.00 (8.90, 11.45) | n.s. | a | 9.50 (7.55, 11.50) | n.s. | a | 9.80 (7.85, 11.63) | n.s. | a | ||
| Cfb (%) | −4.04 (−24.30, 8.78) | a | 10.19 (−6.31, 23.77) | a | 6.10 (−19.16, 27.13) | a | 1.61 (−12.42, 13.71) | a | ||||||
| Fasting blood glucose (mmol/L) (n † = 27, 26, 23, 28) | 0 | 5.89 (±0.47) 5.90 (5.60, 6.20) | a | 5.60 (5.33, 6.23) | a | 5.50 (5.20, 5.75) | a | 5.09 (5.40, 6.10) | a | |||||
| 12 | 5.63 (±0.56) 5.60 (5.35, 5.75) | 0.006 | a | 5.25 (5.10, 5.73) | 0.018 | a | 5.40 (5.05, 5,80) | n.s. | a | 5.50 (5.30, 5.90) | 0.027 | a | ||
| Cfb (%) | −3.85 (−9.92, 0.76) | a | −3.44 (−7.99, 0.00) | a | −1.82 (−7.35, 4.12) | a | −3.39 (−7.42, 1.83) | a | ||||||
| HbA1c (%) (n † = 27, 27, 23, 28) | 0 | 5.31 (±0.36) 5.30 (5.15, 5.50) | a | 5.30 (5.00, 5.50) | a | 5.30 (5.10, 5.55) | a | 5.30 (5.08, 5.50) | a | |||||
| 12 | 5.36 (±0.29) 5.40 (5.20, 5.60) | n.s. | a | 5.20 (5.05, 5.55) | n.s. | a | 5.40 (5.20, 5.50) | n.s. | a | 5.40 (5.18, 5.50) | 0.015 | a | ||
| Cfb (%) | 1.82 (−1.84, 3.77) | a | 1.85 (−1.92, 2.63) | a | 0.00 (−1.75, 3.89) | a | 1.62 (0.00, 3.67) | a | ||||||
| Fasting insulin (mU/L) (n † = 26, 27, 23, 27) | 0 | 8.70 (±3.42) 8.10 (6.05, 11.43) | a | 7.50 (4.60, 9.55) | a | 6.30 (5.35, 9.65) | a | 6.20 (4.95, 7.80) | a | |||||
| 12 | 8.67 (±3.85) 7.95 (6.00, 11.25) | n.s. | a | 6.80 (4.35, 8.00) | n.s. | a | 5.70 (4.30, 8.50) | n.s. | a | 6.00 (4.85, 7.60) | n.s. | a | ||
| Cfb (%) | −9.94 (−29.53, 24.51) | a | −12.50 (−32.32, 8.52) | a | −13.83 (−30.59, 5.24) | a | −6.49 (−21.05, 8.72) | a | ||||||
| Vitamin E (mg) (n † = 27, 27, 23, 28) | 0 | 34.13 (±8.37) | a | 36.57 (±6.32) | a | 37.05 (±9.04) | a | 35.59 (±7.34) | a | |||||
| 12 | 33.60 (±6.78) | n.s. | a | 32.91 (±7.35) | <0.001 | a | 33.58 (±9.08) | 0.004 | a | 33.58 (±6.80) | n.s. | a | ||
| Cfb (%) | −5.47 (−16.03, 11.25) | a | −14.33 (−16.85, −5.60) | a | −7.14 (−14.58, −3.59) | a | −3.86 (−14.65, 4.51) | a | ||||||
| Body weight (kg) (n † = 27, 27, 23, 28) | 0 | 77.86 (±12.06) | a | 80.11 (±14.73) | a | 75.57 (±16.41) | a | 73.44 (±14.44) | a | |||||
| 12 | 76.80 (±11.69) | 0.013 | a | 77.83 (±15.04) | <0.001 | a | 74.62 (±16.30) | 0.03 | a | 72.84 (±14.95) | n.s. | a | ||
| Cfb (%) | −0.75 (−2.08, 0.65) | a | −2.74 (−4.54, −1.08) | b | −0.33 (−2.95, 0.73) | a | −0.66 (−2.06, 0.14) | a | ||||||
| Waist circumferences (cm) (n † = 27, 27, 23, 27) | 0 | 96.24 (±9.64) | a | 95.69 (±12.98) | a | 90.39 (±14.06) | a | 90.80 (±12.36) | a | |||||
| 12 | 93.80 (±9.34) | 0.023 | a | 93.44 (±11.75) | 0.019 | a | 90.43 (±12.33) | n.s. | a | 89.48 (±12.85) | n.s. | a | ||
| Cfb (%) | −2.38 (±5.23) | a | −2.12 (±4.85) | a | 0.56 (±6.75) | a | −1.39 (±4.69) | a | ||||||
| Body fat (kg) (n † = 26, 27, 21, 27) | 0 | 30.23 (±8.20) | a | 33.50 (±7.07) | a | 30.65 (±7.43) | a | 28.49 (±7.95) | a | |||||
| 12 | 29.23 (±7.78) | 0.035 | a | 31.96 (±7.50) | <0.001 | a | 29.38 (±7.82) | 0.018 | a | 27.50 (±8.55) | 0.026 | a | ||
| Cfb (%) | −4.10 (−6.63, −0.49) | a | −5.62 (−8.51, −1.33) | a | −2.33 (−7.50, 0.52) | a | −4.32 (−7.16, 1.09) | a | ||||||
| Body water (l) (n † = 26, 27, 21, 27) | 0 | 39.45 (±7.39) 38.50 (33.70, 44.70) | a | 37.40 (33.65, 41) | a | 34.10 (32.20, 42.10) | a | 38.68 (±7.34) 39.10 (32.25, 45.40) | a | |||||
| 12 | 39.43 (±7.09) 37.75 (34.20, 42.48) | n.s. | a | 37.90 (33.75, 40.45) | n.s. | a | 34.30 (32.40, 40.40) | n.s. | a | 38.75 (±7.71) 37.50 (32.20, 43.20) | n.s. | a | ||
| Cfb (%) | 0.18 (±4.09) | a | −0.68 (±3.21) | a | 0.46 (±2.94) | a | 0.09 (±3.60) | a | ||||||
| Lean body mass (kg) (n † = 26, 27, 21, 27) | 0 | 53.92 (±10.09) 52.60 (46.03, 61.10) | a | 51.10 (45.95, 56.00) | a | 46.50 (44.00, 57.50) | a | 52.84 (±10.04) 53.40 (44.10, 62.00) | a | |||||
| 12 | 53.87 (±9.69) 51.60 (46.73, 58.05) | n.s. | a | 51.80 (46.10, 55.25) | n.s. | a | 46.80 (44.30, 55.20) | n.s. | a | 52.93 (±10.52) 51.20 (44.00, 59.05) | n.s. | a | ||
| Cfb (%) | 0.15 (±4.06) | a | −0.72 (±3.24) | a | 0.46 (±2.89) | a | 0.09 (±3.61) | a | ||||||
| BMI (kg/m2) (n † = 27, 27, 21, 28) | 0 | 25.77 (24.44, 28.70) | a | 27.90 (±4.57) 27.29 (24.45, 30.48) | a | 25.55 (±4.54) 24.90 (23.00, 27.72) | a | 23.90 (22.60, 28.23) | a | |||||
| 12 | 25.70 (24.07, 28.80) | 0.008 | a | 27.12 (±4.70) 26.12 (23.78, 29.39) | <0.001 | a | 25.22 (±4.42) 24.40 (22.30, 27.80) | 0.030 | a | 23.95 (22.34, 28.73) | n.s. | a | ||
| Cfb (%) | −1.45 (±2.70) | a | −2.86 (±2.75) | a | −1.24 (±2.47) | a | −0.91 (±3.15) | a | ||||||
| Systolic blood pressure (mmHG) (n † = 27, 27, 23, 28) | 0 | 139.93 (±19.97) 137.00 (127.00, 152.00) | a | 142.30 (±19.60) | a | 138.48 (±21.73) | a | 140.82 (±23.98) | a | |||||
| 12 | 136.00 (122.50, 143.50) | 0.043 | a | 136.96 (±18.85) 135.00 (125.50, 148.50) | n.s. | a | 131.00 (±16.77) 129.00 (119.50, 142.50) | 0.021 | a | 133.14 (±20.62) 130.00 (119.75, 146.00) | 0.005 | a | ||
| Cfb (%) | −2.93 (±9.48) | a | −3.31 (±8.49) | a | −4.55 (±9.95) | a | −4.75 (±9.63) | a | ||||||
| Diastolic blood pressure (mmHG) (n † = 27, 27, 23, 28) | 0 | 85.00 (78.00, 92.50) | a | 88.00 (80.50, 93.00) | a | 86.35 (±11.23) 84.00 (79.00, 95.50) | a | 85.11 (±10.14) 85.00 (76.75, 95.25) | a | |||||
| 12 | 84.00 (80.00, 89.00) | 0.043 | a | 84.00 (77.50, 90.50) | n.s. | a | 81.22 (±7.45) 81.00 (75.00, 87.50) | 0.005 | a | 82.00 (±9.73) 80.00 (74.50, 88.25) | 0.032 | a | ||
| Cfb (%) | −2.67 (±7.36) | a | −2.21 (±11.41) | a | −5.12 (±9.27) | a | −3.26 (±8.15) | a | ||||||
| Heart rate (bpm) (n † = 27, 27, 23, 28) | 0 | 70.44 (±11.31) 70.00 (62.50, 79.00) | a | 66.00 (61.00, 70.50) | a | 67.26 (±10.98) 67.00 (60.00, 73.00) | a | 66.00 (61.00, 69.25) | a | |||||
| 12 | 69.67 (±9.36) | n.s. | a | 68.07 (±10.13) 68.00 (62.00, 75.00) | n.s. | a | 65.39 (±12.15) | n.s. | a | 66.57 (±10.34) 65.50 (60.00, 71.50) | n.s. | a | ||
| Cfb (%) | −0.06 (±11.88) | a | 2.21 (±12.23) | a | −2.50 (±12.22) | a | −0.09 (±14.52) | a | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drobner, T.; Braun, T.S.; Kiehntopf, M.; Schlattmann, P.; Lorkowski, S.; Dawczynski, C. Evaluation of Influencing Factors on Metabolism of Land-Based n-3 Poly Unsaturated Fatty Acids—The KoALA Study. Nutrients 2023, 15, 4461. https://doi.org/10.3390/nu15204461
Drobner T, Braun TS, Kiehntopf M, Schlattmann P, Lorkowski S, Dawczynski C. Evaluation of Influencing Factors on Metabolism of Land-Based n-3 Poly Unsaturated Fatty Acids—The KoALA Study. Nutrients. 2023; 15(20):4461. https://doi.org/10.3390/nu15204461
Chicago/Turabian StyleDrobner, Timo, Theresa S. Braun, Michael Kiehntopf, Peter Schlattmann, Stefan Lorkowski, and Christine Dawczynski. 2023. "Evaluation of Influencing Factors on Metabolism of Land-Based n-3 Poly Unsaturated Fatty Acids—The KoALA Study" Nutrients 15, no. 20: 4461. https://doi.org/10.3390/nu15204461
APA StyleDrobner, T., Braun, T. S., Kiehntopf, M., Schlattmann, P., Lorkowski, S., & Dawczynski, C. (2023). Evaluation of Influencing Factors on Metabolism of Land-Based n-3 Poly Unsaturated Fatty Acids—The KoALA Study. Nutrients, 15(20), 4461. https://doi.org/10.3390/nu15204461