
Association of Fatty Acid Desaturase 1 rs174547 Polymorphism with the Composition of Long-Chain Polyunsaturated Fatty Acids in Serum Glycerophospholipids during Pregnancy


 ,
, 
Abstract
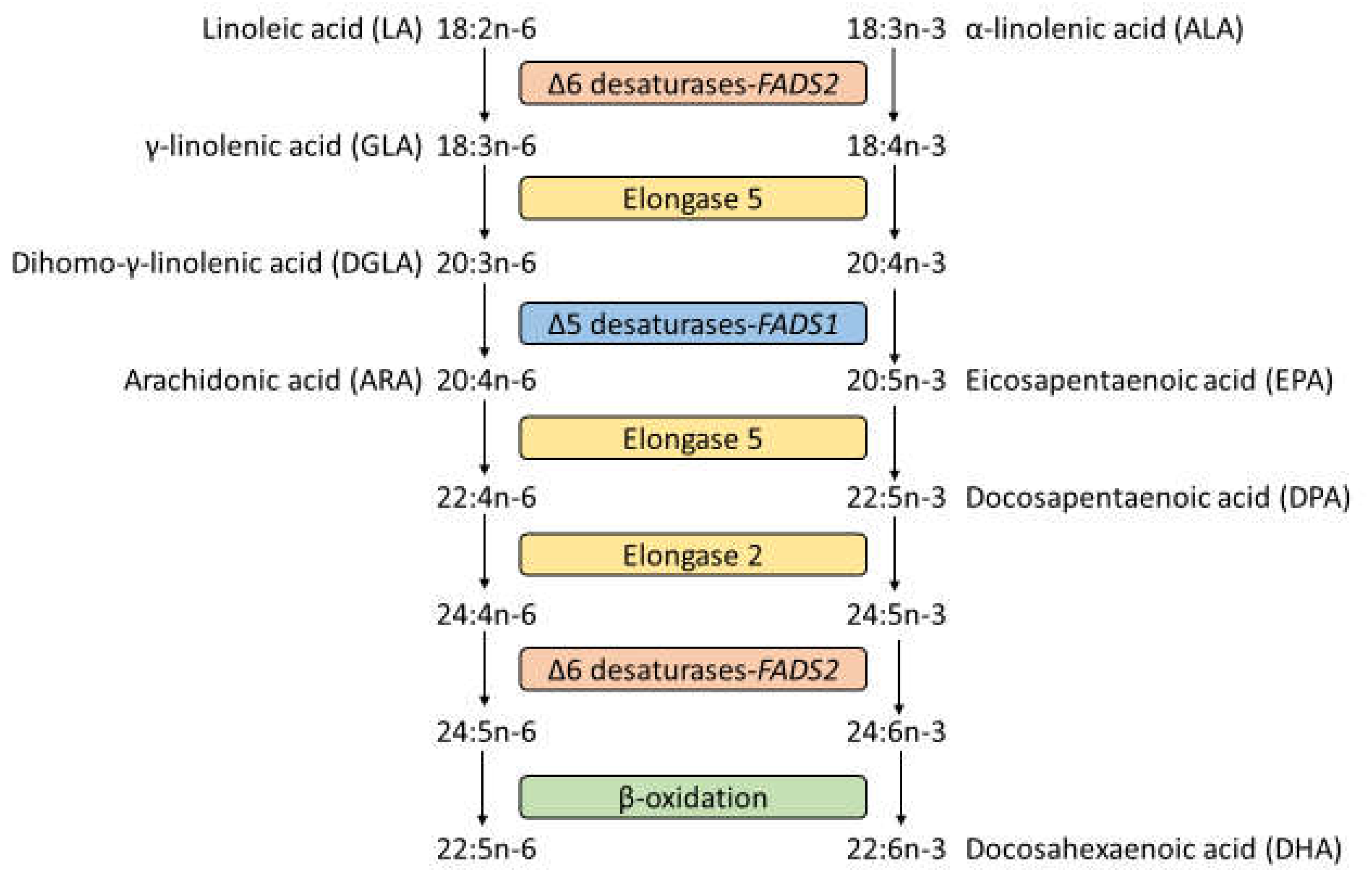
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Study Population
2.3. Maternal and Infant Information
2.4. Dietary Survey
2.5. Fatty Acid Analysis in Serum Glycerophospholipids
2.6. Genotyping
2.7. Statistical Analysis
3. Results
3.1. Participant Characteristics
3.2. Maternal Serum Fatty Acid Changes during Gestation
3.3. Associations between Serum Glycerophospholipid Fatty Acids and rs174547 Polymorphism
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ratnayake, W.M.; Galli, C. Fat and fatty acid terminology, methods of analysis and fat digestion and metabolism: A background review paper. Ann. Nutr. Metab. 2009, 55, 8–43. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, L.; Gohlke, H.; Müller, M.; Heid, I.M.; Palmer, L.J.; Kompauer, I.; Demmelmair, H.; Illig, T.; Koletzko, B.; Heinrich, J. Common genetic variants of the FADS1 FADS2 gene cluster and their reconstructed haplotypes are associated with the fatty acid composition in phospholipids. Hum. Mol. Genet. 2006, 15, 1745–1756. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Reischl, E.; Tanjung, C.; Gonzalez-Casanova, I.; Ramakrishnan, U.; Meldrum, S.; Simmer, K.; Heinrich, J.; Demmelmair, H. FADS1 and FADS2 polymorphisms modulate fatty acid metabolism and dietary impact on health. Annu. Rev. Nutr. 2019, 39, 21–44. [Google Scholar] [CrossRef] [PubMed]
- Alsaleh, A.; Maniou, Z.; Lewis, F.J.; Hall, W.L.; Sanders, T.A.; O’Dell, S.D. ELOVL2 gene polymorphisms are associated with increases in plasma eicosapentaenoic and docosahexaenoic acid proportions after fish oil supplement. Genes Nutr. 2014, 9, 362. [Google Scholar] [CrossRef] [Green Version]
- Lemaitre, R.N.; Tanaka, T.; Tang, W.; Manichaikul, A.; Foy, M.; Kabagambe, E.K.; Nettleton, J.A.; King, I.B.; Weng, L.C.; Bhattacharya, S.; et al. Genetic loci associated with plasma phospholipid n−3 fatty acids: A meta-analysis of genome-wide association studies from the CHARGE Consortium. PLoS Genet. 2011, 7, e1002193. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, K.; Bayasgalan, T.; Tazoe, F.; Yanagisawa, Y.; Gotoh, T.; Yamanaka, K.; Ogawa, A.; Munkhtulga, L.; Chimedregze, U.; Kagawa, Y.; et al. A single nucleotide polymorphism in the FADS1/FADS2 gene is associated with plasma lipid profiles in two genetically similar Asian ethnic groups with distinctive differences in lifestyle. Hum. Genet 2010, 127, 685–690. [Google Scholar] [CrossRef]
- Sasaki, H.; Sueyasu, T.; Tokuda, H.; Ito, M.; Kaneda, Y.; Rogi, T.; Kawashima, H.; Horiguchi, S.; Kawabata, T.; Shibata, H. Aging and FADS1 polymorphisms decrease the biosynthetic capacity of long-chain PUFAs: A human trial using [U-13C]linoleic acid. Prostaglandins Leukot. Essent. Fat Acids 2019, 148, 3. [Google Scholar] [CrossRef]
- Horiguchi, S.; Nakayama, K.; Iwamoto, S.; Ishijima, A.; Minezaki, T.; Baba, M.; Kontai, Y.; Horikawa, C.; Kawashima, H.; Shibata, H.; et al. Associations between a fatty acid desaturase gene polymorphism and blood arachidonic acid compositions in Japanese elderly. Prostaglandins Leukot. Essent. Fat Acids 2016, 105, 9–14. [Google Scholar] [CrossRef]
- Lu, Y.; Vaarhorst, A.; Merry, A.H.; Dollé, M.E.; Hovenier, R.; Imholz, S.; Schouten, L.J.; Heijmans, B.T.; Müller, M.; Slagboom, P.E.; et al. Markers of endogenous desaturase activity and risk of coronary heart disease in the CAREMA cohort study. PLoS ONE 2012, 7, e41681. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Yoo, H.J.; Lee, A.; Jeong, S.; Lee, J.H. Associations among FADS1 rs174547, eicosapentaenoic acid/arachidonic acid ratio, and arterial stiffness in overweight subjects. Prostaglandins Leukot. Essent. Fat Acids 2018, 130, 11–18. [Google Scholar] [CrossRef]
- Cribb, L.; Murphy, J.; Froud, A.; Oliver, G.; Bousman, C.A.; Ng, C.H.; Sarris, J. Erythrocyte polyunsaturated fatty acid composition is associated with depression and FADS genotype in Caucasians. Nutr. Neurosci. 2018, 21, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Middleton, P.; Gomersall, J.C.; Gould, J.F.; Shepherd, E.; Olsen, S.F.; Makrides, M. Omega-3 fatty acid addition during pregnancy. Cochrane Database Syst. Rev. 2018, 11, CD003402. [Google Scholar] [CrossRef]
- Ciesielski, T.H.; Bartlett, J.; Williams, S.M. Omega-3 polyunsaturated fatty acid intake norms and preterm birth rate: A cross-sectional analysis of 184 countries. BMJ Open 2019, 9, e027249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mun, J.G.; Legette, L.L.; Ikonte, C.J.; Mitmesser, S.H. Choline and DHA in Maternal and Infant Nutrition: Synergistic Implications in Brain and Eye Health. Nutrients 2019, 11, 1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nita, R.; Kawabata, T.; Kagawa, Y.; Nakayama, K.; Yanagisawa, Y.; Iwamoto, S.; Kimura, F.; Miyazawa, T.; Tatsuta, N.; Arima, T.; et al. Associations of erythrocyte fatty acid compositions with FADS1 gene polymorphism in Japanese mothers and infants. Prostaglandins Leukot. Essent. Fat Acids 2020, 152, 102031. [Google Scholar] [CrossRef] [Green Version]
- Haggarty, P. Effect of placental function on fatty acid requirements during pregnancy. Eur. J. Clin. Nutr. 2004, 58, 1559–1570. [Google Scholar] [CrossRef] [Green Version]
- Scholtz, S.A.; Kerling, E.H.; Shaddy, D.J.; Li, S.; Thodosoff, J.M.; Colombo, J.; Carlson, S.E. Docosahexaenoic acid (DHA) supplementation in pregnancy differentially modulates arachidonic acid and DHA status across FADS genotypes in pregnancy. Prostaglandins Leukot. Essent Fat. Acids 2015, 94, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Innis, S.M. Genetic variants of the FADS1 FADS2 gene cluster are associated with altered (n−6) and (n−3) essential fatty acids in plasma and erythrocyte phospholipids in women during pregnancy and in breast milk during lactation. J. Nutr. 2008, 138, 2222–2228. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, K.; Miyaso, H.; Eguchi, A.; Matsuno, Y.; Yamamoto, M.; Todaka, E.; Fukuoka, H.; Hata, A.; Mori, C.; On behalf of the Chiba study of Mother and Children’s Health group. Chiba study of Mother and Children’s Health (C-MACH): Cohort study with omics analyses. BMJ Open 2016, 6, e010531. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Honda, S.; Murakami, K.; Sasaki, S.; Okubo, H.; Hirota, N.; Notsu, A.; Fukui, M.; Date, C. Both comprehensive and brief self-administered diet history questionnaires satisfactorily rank nutrient intakes in Japanese adults. J. Epidemiol. 2012, 22, 151–159. [Google Scholar] [CrossRef]
- Glaser, C.; Demmelmair, H.; Koletzko, B. High-throughput analysis of fatty acid composition of plasma glycerophospholipids. J. Lipid Res. 2010, 51, 216–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, A.; Kawabata, T.; Kagawa, Y.; Shoji, K.; Kimura, F.; Miyazawa, T.; Tatsuta, N.; Arima, T.; Yaegashi, N.; Nakai, K. Associations of umbilical cord fatty acid profiles and desaturase enzyme indices with birth weight for gestational age in Japanese infants. Prostaglandins Leukot. Essent. Fat Acids 2021, 165, 102233. [Google Scholar] [CrossRef] [PubMed]
- Niwa, S.; Kawabata, T.; Shoji, K.; Ogata, H.; Kagawa, Y.; Nakayama, K.; Yanagisawa, Y.; Iwamoto, S.; Tatsuta, N.; Asato, K.; et al. Investigation of maternal diet and FADS1 polymorphism associated with long-chain polyunsaturated fatty acid compositions in human milk. Nutrients 2022, 14, 2160. [Google Scholar] [CrossRef] [PubMed]
- de la Garza Puentes, A.; Montes Goyanes, R.; Chisaguano Tonato, A.M.; Torres-Espínola, F.J.; Arias García, M.; de Almeida, L.; Bonilla Aguirre, M.; Guerendiain, M.; Castellote Bargalló, A.I.; Segura Moreno, M.; et al. Association of maternal weight with FADS and ELOVL genetic variants and fatty acid levels- The PREOBE follow-up. PLoS ONE 2017, 12, e0179135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koletzko, B.; Lattka, E.; Zeilinger, S.; Illig, T.; Steer, C. Genetic variants of the fatty acid desaturase gene cluster predict amounts of red blood cell docosahexaenoic and other polyunsaturated fatty acids in pregnant women: Findings from the Avon Longitudinal Study of Parents and Children. Am. J. Clin. Nutr. 2011, 93, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Casanova, I.; Rzehak, P.; Stein, A.D.; Garcia Feregrino, R.; Rivera Dommarco, J.A.; Barraza-Villarreal, A.; Demmelmair, H.; Romieu, I.; Villalpando, S.; Martorell, R.; et al. Maternal single nucleotide polymorphisms in the fatty acid desaturase 1 and 2 coding regions modify the impact of prenatal supplementation with DHA on birth weight. Am. J. Clin. Nutr. 2016, 103, 1171–1178. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, G.Q.; Pereira-Santos, M.; Marcon, L.D.; Louro, I.D.; Peluzio, M.C.G.; Santos, D.B. Maternal polymorphisms in the FADS1 and FADS2 genes modify the association between PUFA ingestion and plasma concentrations of omega-3 polyunsaturated fatty acids. Prostaglandins Leukot. Essent. Fat Acids 2019, 150, 38–46. [Google Scholar] [CrossRef]
- Moltó-Puigmartí, C.; Plat, J.; Mensink, R.P.; Müller, A.; Jansen, E.; Zeegers, M.P.; Thijs, C. FADS1 FADS2 gene variants modify the association between fish intake and the docosahexaenoic acid proportions in human milk. Am. J. Clin. Nutr. 2010, 91, 1368–1376. [Google Scholar] [CrossRef] [Green Version]
- Steer, C.D.; Hibbeln, J.R.; Golding, J.; Davey Smith, G. Polyunsaturated fatty acid levels in blood during pregnancy, at birth and at 7 years: Their associations with two common FADS2 polymorphisms. Hum. Mol. Genet 2012, 21, 1504–1512. [Google Scholar] [CrossRef] [Green Version]
- Santana, J.D.M.; Pereira, M.; Carvalho, G.Q.; Gouveia Peluzio, M.D.C.; Drumond Louro, I.; Santos, D.B.D.; Oliveira, A.M. FADS1 and FADS2 Gene Polymorphisms Modulate the Relationship of Omega-3 and Omega-6 Fatty Acid Plasma Concentrations in Gestational Weight Gain: A NISAMI Cohort Study. Nutrients 2022, 14, 1056. [Google Scholar] [CrossRef]
- Yeates, A.J.; Love, T.M.; Engström, K.; Mulhern, M.S.; McSorley, E.M.; Grzesik, K.; Alhamdow, A.; Wahlberg, K.; Thurston, S.W.; Davidson, P.W.; et al. Genetic variation in FADS genes is associated with maternal long-chain PUFA status but not with cognitive development of infants in a high fish-eating observational study. Prostaglandins Leukot. Essent. Fat Acids 2015, 102–103, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.C.; Chang, W.T.; Chang, H.Y.; Chung, H.F.; Chen, F.P.; Huang, Y.F.; Hsu, C.C.; Hwang, S.J. FADS Gene Polymorphisms, Fatty Acid Desaturase Activities, and HDL-C in Type 2 Diabetes. Int. J. Environ. Res. Public Health 2017, 14, 572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Tang, Y.; Ji, Y.; Xu, W.; Ullah, N.; Yu, H.; Wu, Y.; Xie, L. Association between FADS1 rs174547 and levels of long-chain PUFA: A meta-analysis. Br. J. Nutr. 2021, 126, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Al, M.D.; van Houwelingen, A.C.; Kester, A.D.; Hasaart, T.H.; de Jong, A.E.; Hornstra, G. Maternal essential fatty acid patterns during normal pregnancy and their relationship to the neonatal essential fatty acid status. Br. J. Nutr. 1995, 74, 55–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.B.; Li, H.T.; Trasande, L.; Wang, L.L.; Zhang, Y.L.; Si, K.Y.; Bai, M.X.; Liu, J.M. A Correlation Study of DHA Intake Estimated by a FFQ and Concentrations in Plasma and Erythrocytes in Mid- and Late Pregnancy. Nutrients 2017, 9, 1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, C.; Speake, B.K.; Cameron, A.; Sattar, N.; Weaver, L.T. Maternal docosahexaenoic acid supplementation and fetal accretion. Br. J. Nutr. 2003, 90, 135–145. [Google Scholar] [CrossRef]
- Vlaardingerbroek, H.; Hornstra, G. Essential fatty acids in erythrocyte phospholipids during pregnancy and at delivery in mothers and their neonates: Comparison with plasma phospholipids. Prostaglandins Leukot. Essent. Fat Acids 2004, 71, 363–374. [Google Scholar] [CrossRef]
- Kawabata, T.; Kagawa, Y.; Kimura, F.; Miyazawa, T.; Saito, S.; Arima, T.; Nakai, K.; Yaegashi, N. Polyunsaturated Fatty Acid Levels in Maternal Erythrocytes of Japanese Women during Pregnancy and after Childbirth. Nutrients 2017, 9, 245. [Google Scholar] [CrossRef] [Green Version]
- Kuipers, R.S.; Luxwolda, M.F.; Offringa, P.J.; Boersma, E.R.; Dijck-Brouwer, D.A.; Muskiet, F.A. Fetal intrauterine whole body linoleic, arachidonic and docosahexaenoic acid contents and accretion rates. Prostaglandins Leukot. Essent. Fat Acids 2012, 86, 13–20. [Google Scholar] [CrossRef]
- Henderson, R.J.; Burkow, I.C.; Buzzi, M.; Bayer, A. Effects of docosahexaenoic (22:6n−3), tetracosapentaenoic (24:5n−3) and tetracosahexaenoic (24:6n−3) acids on the desaturation and elongation of n−3 polyunsaturated fatty acids in trout liver microsomes. Biochim. Biophys Acta 1998, 1392, 309–319. [Google Scholar] [CrossRef]
- Bakewell, L.; Burdge, G.C.; Calder, P.C. Polyunsaturated fatty acid concentrations in young men and women consuming their habitual diets. Br. J. Nutr. 2006, 96, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdge, G.C.; Wootton, S.A. Conversion of alpha-linolenic acid to eicosapentaenoic, docosapentaenoic and docosahexaenoic acids in young women. Br. J. Nutr. 2002, 88, 411–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdge, G.C.; Jones, A.E.; Wootton, S.A. Eicosapentaenoic and docosapentaenoic acids are the principal products of alpha-linolenic acid metabolism in young men. Br. J. Nutr. 2002, 88, 355–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giltay, E.J.; Gooren, L.J.; Toorians, A.W.; Katan, M.B.; Zock, P.L. Docosahexaenoic acid concentrations are higher in women than in men because of estrogenic effects. Am. J. Clin. Nutr. 2004, 80, 1167–1174. [Google Scholar] [CrossRef] [Green Version]
- Stark, K.D.; Beblo, S.; Murthy, M.; Buda-Abela, M.; Janisse, J.; Rockett, H.; Whitty, J.E.; Martier, S.S.; Sokol, R.J.; Hannigan, J.H.; et al. Comparison of bloodstream fatty acid composition from African-American women at gestation, delivery, and postpartum. J. Lipid. Res. 2005, 46, 516–525. [Google Scholar] [CrossRef] [Green Version]
- De Haas, S.; Ghossein-Doha, C.; van Kuijk, S.M.; van Drongelen, J.; Spaanderman, M.E. Physiological adaptation of maternal plasma volume during pregnancy: A systematic review and meta-analysis. Ultrasound Obstet. Gynecol. 2017, 49, 177–187. [Google Scholar] [CrossRef]


{kind=link}
| Variables | n | (%) | Median | (25th–75th) |
|---|---|---|---|---|
| Mothers | ||||
| Age at participation | ||||
| <30 years old | 63 | (27.9%) | ||
| 30–39 years old | 148 | (65.5%) | ||
| ≧40 years old | 15 | (6.6%) | ||
| Missing | 16 | - | ||
| Height, cm | 226 | 159 | (155–163) | |
| Prepregnancy weight, kg | 226 | 52 | (48–58) | |
| Prepregnancy body mass index (kg/cm2) | 226 | 20.7 | (19.1–22.9) | |
| Parity | ||||
| 0 | 91 | (40.8%) | ||
| 1 | 96 | (43.1%) | ||
| ≧2 | 36 | (16.1%) | ||
| Missing | 19 | - | ||
| Smoking status in the first trimester of pregnancy | ||||
| Smoker or quit | 48 | (21.3%) | ||
| Never | 177 | (78.7%) | ||
| Missing | 17 | - | ||
| Smoking status in the third trimester of pregnancy | ||||
| Smoker or quit | 52 | (22.7%) | ||
| Never | 177 | (77.3%) | ||
| Missing | 13 | - | ||
| FADS1; rs174547 genotype | ||||
| TT | 97 | (40.1%) | ||
| TC | 108 | (44.6%) | ||
| CC | 37 | (15.3%) | ||
| Infants | ||||
| Gestational period, day | 226 | 278 | (271–284) | |
| Sex | ||||
| Male | 117 | (52.2%) | ||
| Female | 107 | (47.8%) | ||
| Missing | 18 | |||
| FADS1; rs174547 genotype | ||||
| TT | 80 | (35.2%) | ||
| TC | 113 | (49.8%) | ||
| CC | 34 | (15.0%) | ||
| Missing | 15 | - | ||
| The 1st Trimester | The 3rd Trimester | At Delivery | p2 | ||||
|---|---|---|---|---|---|---|---|
| n = 242 | n = 237 | n = 213 | |||||
| SFA (%) | 40.14 | (39.01–41.20) a | 41.78 | (40.28–42.76) b | 41.89 | (40.88–42.82) b | <0.001 |
| MUFA (%) | 18.18 | (16.55–19.50) a | 18.48 | (17.17–19.82) b | 18.60 | (17.51–19.96) b | 0.001 |
| PUFA (%) | 41.90 | (40.6–43.32) a | 40.02 | (38.75–41.06) b | 39.46 | (38.33–40.80) b | <0.001 |
| n−6 (%) | |||||||
| 18:2n−6 (LA) | 22.91 | (21.01–24.74) | 22.99 | (21.16–24.60) | 22.93 | (21.26–24.85) | 0.943 |
| 18:3n−6 (GLA) 3 | 0.07 | (0.05–0.11) a | 0.05 | (0.03–0.07) b | 0.05 | (0.04–0.08) c | <0.001 |
| 20:3n−6 (DGLA) | 2.13 | (1.74–2.55) a | 2.40 | (2.12–2.75) b | 2.44 | (2.08–2.74) b | <0.001 |
| 20:4n−6 (ARA) | 8.76 | (7.73–9.72) a | 7.07 | (6.32–7.85) b | 6.92 | (6.14–7.84) b | <0.001 |
| n−3 (%) | |||||||
| 18:3n−3 (ALA) 3 | 0.51 | (0.40–0.64) a | 0.58 | (0.47–0.70) b | 0.55 | (0.47–0.64) b | <0.001 |
| 20:5n−3 (EPA) 3 | 0.72 | (0.52–1.08) a | 0.58 | (0.38–0.95) b | 0.65 | (0.39–1.00) b | <0.001 |
| 22:5n−3 (DPA) | 0.63 | (0.54–0.76) a | 0.52 | (0.42–0.62) b | 0.47 | (0.37–0.56) c | <0.001 |
| 22:6n−3 (DHA) | 4.84 | (4.16–5.30) a | 4.47 | (3.75–5.23) b | 4.28 | (3.50–5.01) c | <0.001 |
| Ratios | |||||||
| ARA/LA ratio | 0.37 | (0.32–0.45) a | 0.31 | (0.27–0.37) b | 0.30 | (0.26–0.36) b | <0.001 |
| ARA/DGLA ratio 3 | 4.10 | (3.31–4.93) a | 2.91 | (2.56–3.38) b | 2.91 | (2.57–3.30) b | <0.001 |
| Objective Variable 2 | The 1st Trimester | The 3rd Trimester | At Delivery | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n = 185 | n = 180 | n = 165 | |||||||
| B | Std β | p | B | Std β | p | B | Std β | p | |
| SFA | −0.216 | −0.090 | 0.233 | −0.050 | −0.019 | 0.801 | −0.342 | −0.165 | 0.036 |
| MUFA | 0.369 | 0.122 | 0.091 | 0.333 | 0.132 | 0.079 | 0.566 | 0.215 | 0.005 |
| PUFA | −0.159 | −0.052 | 0.469 | −0.249 | −0.094 | 0.207 | −0.186 | −0.064 | 0.410 |
| n−6 | |||||||||
| 18:2n−6 (LA) | 1.316 | 0.351 | <0.001 | 1.179 | 0.348 | <0.001 | 1.289 | 0.395 | <0.001 |
| 18:3n−6 (GLA) | −0.449 | −0.466 | <0.001 | −0.232 | −0.271 | <0.001 | −0.326 | −0.360 | <0.001 |
| 20:3n−6 (DGLA) | −0.176 | −0.216 | 0.003 | −0.144 | −0.212 | 0.004 | −0.098 | −0.154 | 0.051 |
| 20:4n−6 (ARA) | −1.089 | −0.491 | <0.001 | −0.953 | −0.636 | <0.001 | −0.926 | −0.562 | <0.001 |
| n−3 | |||||||||
| 18:3n−3 (ALA) | 0.116 | 0.223 | 0.003 | 0.098 | 0.231 | 0.002 | 0.036 | 0.157 | 0.047 |
| 20:5n−3 (EPA) | −0.159 | −0.222 | 0.002 | −0.127 | −0.142 | 0.052 | −0.133 | −0.141 | 0.063 |
| 22:5n−3 (DPA) | −0.015 | −0.061 | 0.414 | −0.001 | −0.003 | 0.970 | 0.005 | 0.025 | 0.745 |
| 22:6n−3 (DHA) | −0.108 | −0.074 | 0.321 | −0.278 | −0.193 | 0.008 | −0.307 | −0.208 | 0.006 |
| Ratios | |||||||||
| ARA/LA ratio | −0.069 | −0.498 | <0.001 | −0.059 | −0.626 | <0.001 | −0.057 | −0.581 | <0.001 |
| ARA/DGLA ratio | −0.116 | −0.063 | 0.374 | −0.204 | −0.219 | 0.003 | −0.228 | −0.254 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawabata, T.; Fukuoka, H.; Harada, M.; Shoji, K.; Kubo, Y.; Mori, C.; Sakurai, K.; Ohkubo, T.; Oshida, K.; Yamashiro, Y. Association of Fatty Acid Desaturase 1 rs174547 Polymorphism with the Composition of Long-Chain Polyunsaturated Fatty Acids in Serum Glycerophospholipids during Pregnancy. Nutrients 2023, 15, 722. https://doi.org/10.3390/nu15030722
Kawabata T, Fukuoka H, Harada M, Shoji K, Kubo Y, Mori C, Sakurai K, Ohkubo T, Oshida K, Yamashiro Y. Association of Fatty Acid Desaturase 1 rs174547 Polymorphism with the Composition of Long-Chain Polyunsaturated Fatty Acids in Serum Glycerophospholipids during Pregnancy. Nutrients. 2023; 15(3):722. https://doi.org/10.3390/nu15030722
Chicago/Turabian StyleKawabata, Terue, Hideoki Fukuoka, Michiru Harada, Kumiko Shoji, Yoshinori Kubo, Chisato Mori, Kenichi Sakurai, Takeshi Ohkubo, Kyoichi Oshida, and Yuichiro Yamashiro. 2023. "Association of Fatty Acid Desaturase 1 rs174547 Polymorphism with the Composition of Long-Chain Polyunsaturated Fatty Acids in Serum Glycerophospholipids during Pregnancy" Nutrients 15, no. 3: 722. https://doi.org/10.3390/nu15030722
APA StyleKawabata, T., Fukuoka, H., Harada, M., Shoji, K., Kubo, Y., Mori, C., Sakurai, K., Ohkubo, T., Oshida, K., & Yamashiro, Y. (2023). Association of Fatty Acid Desaturase 1 rs174547 Polymorphism with the Composition of Long-Chain Polyunsaturated Fatty Acids in Serum Glycerophospholipids during Pregnancy. Nutrients, 15(3), 722. https://doi.org/10.3390/nu15030722