
Cinnamomum cassia and Rosa laevigata Mixture Improves Benign Prostatic Hyperplasia in Rats by Regulating Androgen Receptor Signaling and Apoptosis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of HT080
2.2. High-Performance Liquid Chromatography (HPLC) of HT080 Extract
2.3. Animal Experimental Design
2.4. Histology
2.5. Determination of Testosterone and DHT Levels in Serum
2.6. Measurement of Serum Alanine Aminotransferase (ALT) and Aspartate Aminotransferase (AST) Levels
2.7. Western Blot Analysis
2.8. Hershberger Bioassay
2.9. Statistical Analysis
3. Results
3.1. HPLC Analysis of Cinnamic Acid and Ellagic Acid in the HT080 Extract
3.2. Effects of HT080 on Prostate Weight and PI
3.3. Effects of HT080 on Prostate Tissue Histology
3.4. Serum Concentration of Testosterone and DHT
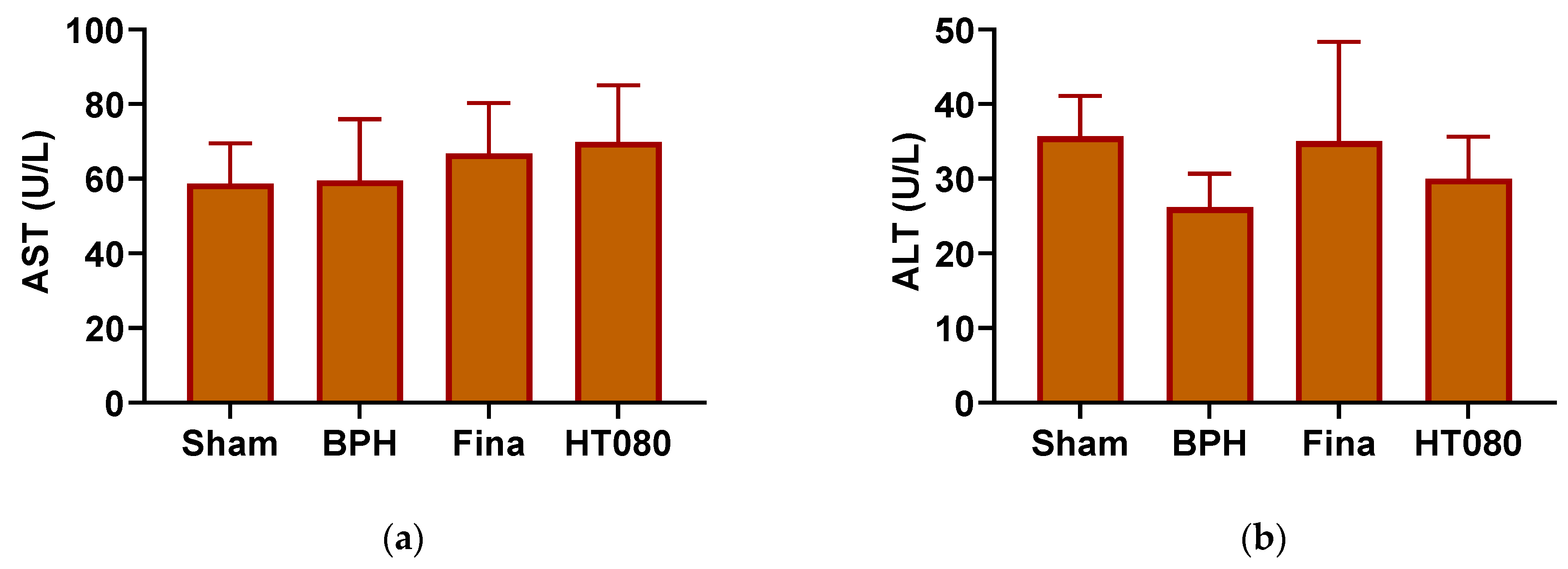
3.5. Serum ALT and AST Levels
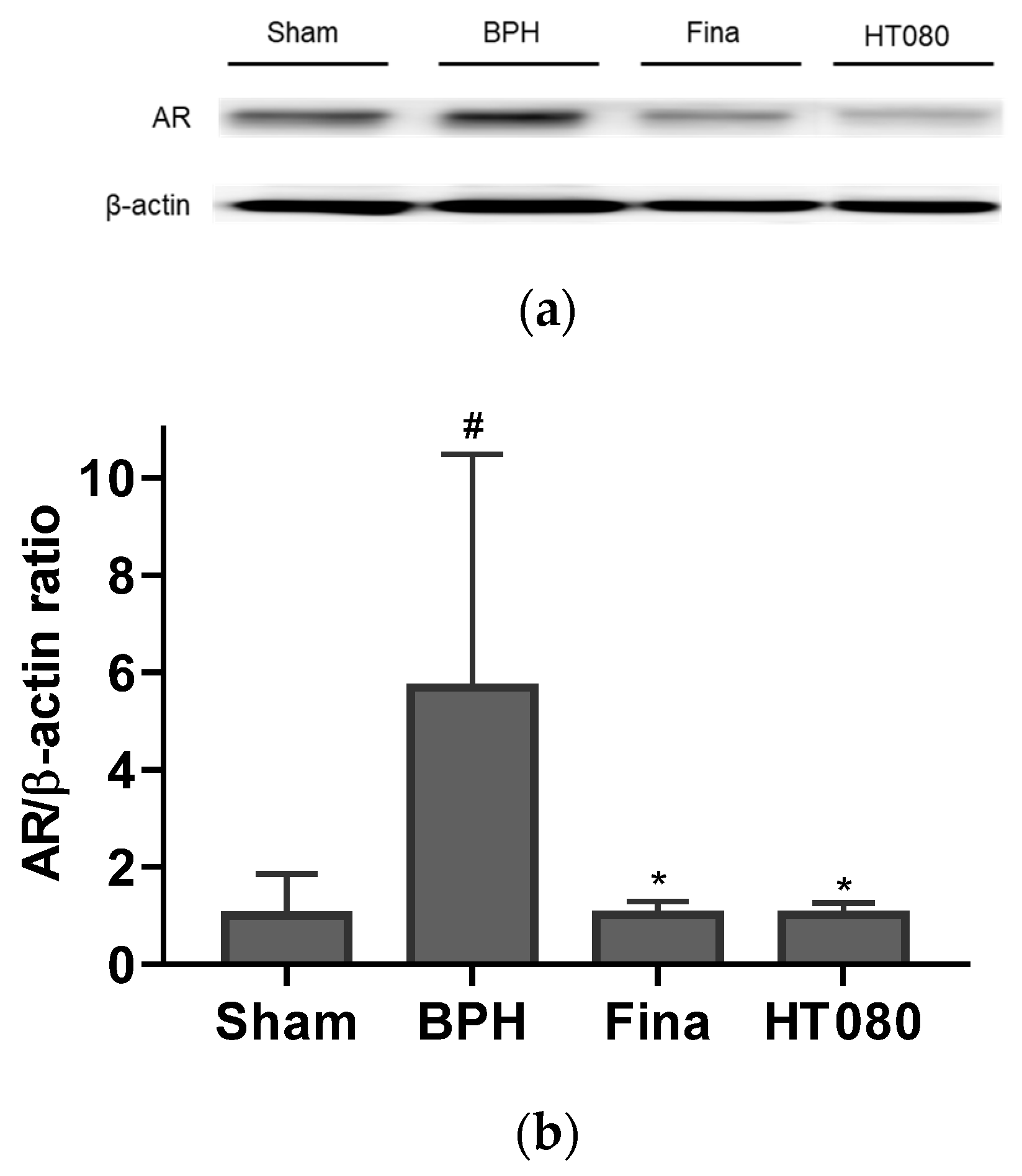
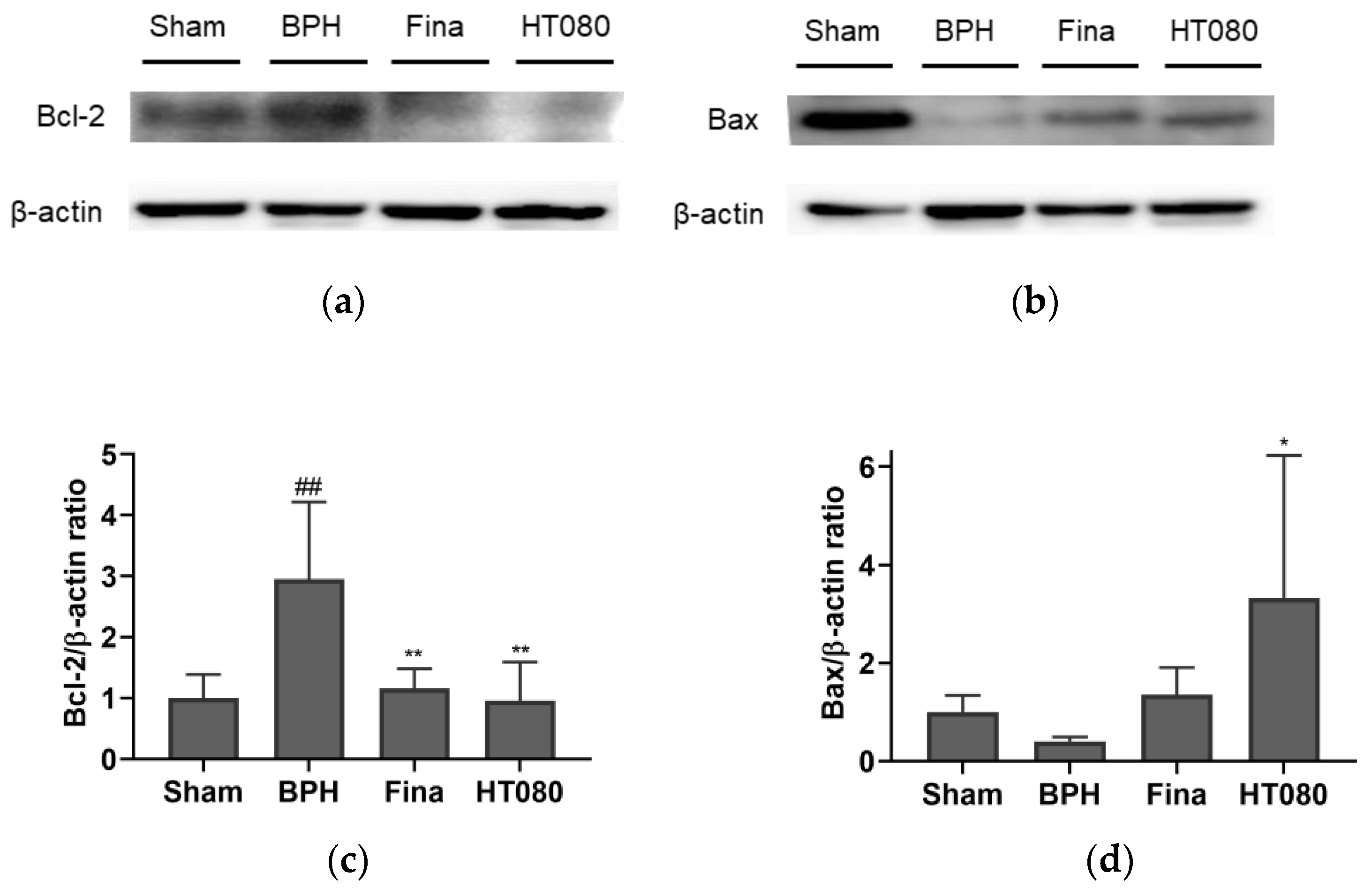
3.6. Effects of HT080 on AR and Apoptosis in BPH-Induced Rats
3.7. Effects of HT080 on Antiandrogenic and Androgenic Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- William, F.P.; Russa, D.A. Prunus africanus Herbal Extracts Reverse and Ameliorate the Histological and Histomorphometric Changes in Testosterone-Induced Benign Prostate Hyperplasia Rat Models. Tanzan. J. Sci. 2021, 47, 1184–1194. [Google Scholar] [CrossRef]
- Barkin, J.; Roehrborn, C.G.; Siami, P.; Haillot, O.; Morrill, B.; Black, L.; Montorsi, F. Effect of Dutasteride, Tamsulosin and the Combination on Patient-Reported Quality of Life and Treatment Satisfaction in Men with Moderate-to-Severe Benign Prostatic Hyperplasia: 2-Year Data from the CombAT Trial. BJU Int. 2009, 103, 919–926. [Google Scholar] [CrossRef]
- Barkin, J. Benign Prostatic Hyperplasia and Lower Urinary Tract Symptoms: Evidence and Approaches for Best Case Management. Can. J. Urol. 2011, 18 (Suppl. S1), 14–19. [Google Scholar]
- Barry, M.J.; Fowler, F.J.; O’Leary, M.P.; Bruskewitz, R.C.; Holtgrewe, H.L.; Mebust, W.K.; Cockett, A.T.K. The American Urological Association Symptom Index for Benign Prostatic Hyperplasia. J. Urol. 1992, 148, 1549–1557. [Google Scholar] [CrossRef]
- Bortnick, E.; Brown, C.; Simma-Chiang, V.; Kaplan, S.A. Modern Best Practice in the Management of Benign Prostatic Hyperplasia in the Elderly. Ther. Adv. Urol. 2020, 12, 175628722092948. [Google Scholar] [CrossRef]
- Kevin, T.; McVary, M.D.F. Epidemiology and Pathophysiology of Benign Prostatic Hyperplasia—UpToDate. Available online: https://www.uptodate.com/contents/epidemiology-and-pathophysiology-of-benign-prostatic-hyperplasia (accessed on 22 November 2022).
- Parsons, J.K. Benign Prostatic Hyperplasia and Male Lower Urinary Tract Symptoms: Epidemiology and Risk Factors. Curr. Bladder Dysfunct. Rep. 2010, 5, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Kopp, W. Diet-Induced Hyperinsulinemia as a Key Factor in the Etiology of Both Benign Prostatic Hyperplasia and Essential Hypertension? Nutr. Metab. Insights 2018, 11, 117863881877307. [Google Scholar] [CrossRef] [PubMed]
- Castro-Diaz, D. Exercise, Diet and Weight Loss before Therapy for Lower Urinary Tract Symptoms/Benign Prostatic Hyperplasia? BJU Int. 2015, 116, 168–169. [Google Scholar] [CrossRef] [PubMed]
- Krušlin, B.; Tomas, D.; Džombeta, T.; Milković-Periša, M.; Ulamec, M. Inflammation in Prostatic Hyperplasia and Carcinoma—Basic Scientific Approach. Front. Oncol. 2017, 7, 77. [Google Scholar] [CrossRef]
- Tjahjodjati, I.S.; Noegroho, B.S.; Sihombing, A.T. Urinary Tract Stones Risk Factors in Patients with Benign Prostatic Hyperplasia in West Java, Indonesia. Althea Med. J. 2021, 8, 93–98. [Google Scholar] [CrossRef]
- Wu, Y.; Ding, Y.; Cao, Q.F.; Qian, S.B.; Wang, C.; Duan, H.Q.; Gu, J.; Shen, H.B. The Relationship between Glucose Homeostasis Status and Prostate Size in Aging Chinese Males with Benign Prostatic Hyperplasia. World J. Urol. 2020, 38, 2923–2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, T.M.; Ricke, W.A. Androgens and Estrogens in Benign Prostatic Hyperplasia: Past, Present and Future. Differentiation 2011, 82, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Buncharoen, W.; Saenphet, K.; Saenphet, S.; Thitaram, C. Uvaria Rufa Blume Attenuates Benign Prostatic Hyperplasia via Inhibiting 5α-Reductase and Enhancing Antioxidant Status. J. Ethnopharmacol. 2016, 194, 483–494. [Google Scholar] [CrossRef]
- Bartsch, G.; Rittmaster, R.S.; Klocker, H. Dihydrotestosterone and the Concept of 5alpha-Reductase Inhibition in Human Benign Prostatic Hyperplasia. Eur. Urol. 2000, 37, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Sciarra, A.; Mariotti, G.; Salciccia, S.; Gomez, A.A.; Monti, S.; Toscano, V.; Di Silverio, F. Prostate Growth and Inflammation. J. Steroid Biochem. Mol. Biol. 2008, 108, 254–260. [Google Scholar] [CrossRef]
- Miano, R.; De Nunzio, C.; Asimakopoulos, A.D.; Germani, S.; Tubaro, A. Treatment Options for Benign Prostatic Hyperplasia in Older Men. Med. Sci. Monit. 2008, 14, RA94–RA102. [Google Scholar]
- Foley, C.L.; Kirby, R.S. 5 Alpha-Reductase Inhibitors: What’s New? Curr. Opin. Urol. 2003, 13, 31–37. [Google Scholar] [CrossRef]
- Gravas, S.; Oelke, M. Current Status of 5α-Reductase Inhibitors in the Management of Lower Urinary Tract Symptoms and BPH. World J. Urol. 2010, 28, 9–15. [Google Scholar] [CrossRef]
- Mysore, V. Finasteride and Sexual Side Effects. Indian Dermatol. Online J. 2012, 3, 62. [Google Scholar] [CrossRef]
- Moinpour, C.M.; Darke, A.K.; Donaldson, G.W.; Thompson, I.M.; Langley, C.; Ankerst, D.P.; Patrick, D.L.; Ware, J.E.; Ganz, P.A.; Shumaker, S.A.; et al. Longitudinal Analysis of Sexual Function Reported by Men in the Prostate Cancer Prevention Trial. JNCI J. Natl. Cancer Inst. 2007, 99, 1025–1035. [Google Scholar] [CrossRef]
- Traish, A.M.; Hassani, J.; Guay, A.T.; Zitzmann, M.; Hansen, M.L. Adverse Side Effects of 5α-reductase Inhibitors Therapy: Persistent Diminished Libido and Erectile Dysfunction and Depression in a Subset of Patients. J. Sex. Med. 2011, 8, 872–884. [Google Scholar] [CrossRef] [PubMed]
- Hirshburg, J.M.; Kelsey, P.A.; Therrien, C.A.; Gavino, A.C.; Reichenberg, J.S. Adverse Effects and Safety of 5-Alpha Reductase Inhibitors (Finasteride, Dutasteride): A Systematic Review. J. Clin. Aesthet. Dermatol. 2016, 9, 56–62. [Google Scholar]
- Wallner, L.P.; DiBello, J.R.; Li, B.H.; Van Den Eeden, S.K.; Weinmann, S.; Ritzwoller, D.P.; Abell, J.E.; D’Agostino, R.; Loo, R.K.; Aaronson, D.S.; et al. The Use of 5-Alpha Reductase Inhibitors to Manage Benign Prostatic Hyperplasia and the Risk of All-Cause Mortality. Urology 2018, 119, 70–78. [Google Scholar] [CrossRef]
- Deng, T.; Lin, X.; Duan, X.; He, Z.; Zhao, Z.; Zeng, G. Prostate Cancer Patients Can Benefit from 5-Alpha-Reductase Inhibitor Treatment: A Meta-Analysis. PeerJ 2020, 8, e9282. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Grady, J.J.; Albertsen, P.C.; Helen Wu, Z. Tamsulosin and the Risk of Dementia in Older Men with Benign Prostatic Hyperplasia. Pharmacoepidemiol. Drug Saf. 2018, 27, 340–348. [Google Scholar] [CrossRef]
- Frankel, J.K.; Duan, Y.; Albertsen, P.C. Is Tamsulosin Linked to Dementia in the Elderly? Curr. Urol. Rep. 2018, 19, 69. [Google Scholar] [CrossRef] [PubMed]
- Bullock, T.L.; Andriole, G.L., Jr. Emerging Drug Therapies for Benign Prostatic Hyperplasia. Expert Opin. Emerg. Drugs 2006, 11, 111–123. [Google Scholar] [CrossRef]
- Karunasagara, S.; Hong, G.-L.; Jung, D.-Y.; Kim, K.-H.; Cho, K.; Jung, J.-Y. Protective Effects of Combination of Stauntonia hexaphylla and Cornus officinalis on Testosterone-Induced Benign Prostatic Hyperplasia through Inhibition of 5α—Reductase Type 2 and Induced Cell Apoptosis. PLoS ONE 2020, 15, e0236879. [Google Scholar] [CrossRef]
- Elsherbini, D.M.A.; Almohaimeed, H.M.; El-Sherbiny, M.; Mohammedsaleh, Z.M.; Elsherbiny, N.M.; Gabr, S.A.; Ebrahim, H.A. Origanum majorana L. Extract Attenuated Benign Prostatic Hyperplasia in Rat Model: Effect on Oxidative Stress, Apoptosis, and Proliferation. Antioxidants 2022, 11, 1149. [Google Scholar] [CrossRef]
- Choi, H.-M.; Jung, Y.; Park, J.; Kim, H.-L.; Youn, D.-H.; Kang, J.; Jeong, M.-Y.; Lee, J.-H.; Yang, W.M.; Lee, S.-G.; et al. Cinnamomi Cortex (Cinnamomum verum) Suppresses Testosterone-Induced Benign Prostatic Hyperplasia by Regulating 5α-Reductase. Sci. Rep. 2016, 6, 31906. [Google Scholar] [CrossRef]
- Park, W.Y.; Song, G.; Park, J.Y.; Ahn, K.S.; Kwak, H.J.; Park, J.; Lee, J.H.; Um, J.-Y. Ellagic Acid Improves Benign Prostate Hyperplasia by Regulating Androgen Signaling and STAT3. Cell Death Dis. 2022, 13, 554. [Google Scholar] [CrossRef]
- Chang, C.-T.; Chang, W.-L.; Hsu, J.-C.; Shih, Y.; Chou, S.-T. Chemical Composition and Tyrosinase Inhibitory Activity of Cinnamomum cassia Essential Oil. Bot. Stud. 2013, 54, 10. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Wu, H. Analysis and Evaluation of Essential Oil Components of Cinnamon Barks Using GC–MS and FTIR Spectroscopy. Ind. Crops Prod. 2013, 41, 269–278. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Yu, J.-W.; Lu, F.-L.; Lin, M.-C.; Cheng, H.-F. Differentiating Parts of Cinnamomum cassia Using LC-QTOF-MS in Conjunction with Principal Component Analysis. Biomed. Chromatogr. 2016, 30, 1449–1457. [Google Scholar] [CrossRef]
- Zhang, C.; Fan, L.; Fan, S.; Wang, J.; Luo, T.; Tang, Y.; Chen, Z.; Yu, L. Cinnamomum cassia Presl: A Review of Its Traditional Uses, Phytochemistry, Pharmacology and Toxicology. Molecules 2019, 24, 3473. [Google Scholar] [CrossRef] [PubMed]
- Li, B.-L.; Yuan, J.; Wu, J.-W. A Review on the Phytochemical and Pharmacological Properties of Rosa laevigata: A Medicinal and Edible Plant. Chem. Pharm. Bull. 2021, 69, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Sun, Z.; Xu, H.; Zhang, S. Effect of Aqueous Extract of Rosa laevigata on Urinary System. J. Electron. Publ. House 1995, 26, 529–531. [Google Scholar]
- Chang, H.-K.; Hsu, F.-L.; Liu, I.-M.; Cheng, J.-T. Stimulatory Effect of Cinnamic Acid Analogues on A1A-Adrenoceptors in-Vitro. J. Pharm. Pharmacol. 2010, 55, 833–837. [Google Scholar] [CrossRef]
- Mueller-Harvey, I. Analysis of Hydrolysable Tannins. Anim. Feed Sci. Technol. 2001, 91, 3–20. [Google Scholar] [CrossRef]
- Brown, C. Intra-Abdominal Castration in the Rat. Lab. Anim. 2008, 37, 73–74. [Google Scholar] [CrossRef]
- Veeresh Babu, S.V.; Veeresh, B.; Patil, A.A.; Warke, Y.B. Lauric Acid and Myristic Acid Prevent Testosterone Induced Prostatic Hyperplasia in Rats. Eur. J. Pharmacol. 2010, 626, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Arruzazabala, M.L.; Mas, R.; Molina, V.; Noa, M.; Carbajal, D.; Mendoza, N. Effect of D-004, a Lipid Extract from the Cuban Royal Palm Fruit, on Atypical Prostate Hyperplasia Induced by Phenylephrine in Rats. Drugs R D 2006, 7, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fang, T.; Li, M.; Song, Y.; Li, J.; Xue, Z.; Li, J.; Bu, D.; Liu, W.; Zeng, Q.; et al. Pao Pereira Extract Attenuates Testosterone-Induced Benign Prostatic Hyperplasia in Rats by Inhibiting 5α-Reductase. Sci. Rep. 2019, 9, 19703. [Google Scholar] [CrossRef]
- Izumi, K.; Mizokami, A.; Lin, W.-J.; Lai, K.-P.; Chang, C. Androgen Receptor Roles in the Development of Benign Prostate Hyperplasia. Am. J. Pathol. 2013, 182, 1942–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DePrimo, S.E.; Diehn, M.; Nelson, J.B.; Reiter, R.E.; Matese, J.; Fero, M.; Tibshirani, R.; Brown, P.O.; Brooks, J.D. Transcriptional Programs Activated by Exposure of Human Prostate Cancer Cells to Androgen. Genome Biol. 2002, 3, RESEARCH0032. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, T.M.; Sehgal, P.D.; Drew, S.A.; Huang, W.; Ricke, W.A. Sex Steroid Receptor Expression and Localization in Benign Prostatic Hyperplasia Varies with Tissue Compartment. Differentiation 2013, 85, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Kyprianou, N.; Tu, H.; Jacobs, S.C. Apoptotic versus Proliferative Activities in Human Benign Prostatic Hyperplasia. Hum. Pathol. 1996, 27, 668–675. [Google Scholar] [CrossRef]
- Li, C.; Hu, W.-L.; Lu, M.-X.; Xiao, G.-F. Resveratrol Induces Apoptosis of Benign Prostatic Hyperplasia Epithelial Cell Line (BPH-1) through P38 MAPK-FOXO3a Pathway. BMC Complement. Altern. Med. 2019, 19, 233. [Google Scholar] [CrossRef]
- Liu, J.; Yin, J.; Chen, P.; Liu, D.; He, W.; Li, Y.; Li, M.; Fu, X.; Zeng, G.; Guo, Y.; et al. Smoothened Inhibition Leads to Decreased Cell Proliferation and Suppressed Tissue Fibrosis in the Development of Benign Prostatic Hyperplasia. Cell Death Discov. 2021, 7, 115. [Google Scholar] [CrossRef]
- Li, F.; Pascal, L.E.; Zhou, J.; Zhou, Y.; Wang, K.; Parwani, A.V.; Dhir, R.; Guo, P.; He, D.; Nelson, J.B.; et al. BCL-2 and BCL-XL Expression Are down-Regulated in Benign Prostate Hyperplasia Nodules and Not Affected by Finasteride and/or Celecoxib. Am. J. Clin. Exp. Urol. 2018, 6, 1–10. [Google Scholar]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of Apoptosis by the Bcl-2 Protein Family: Implications for Physiology and Therapy. Nat. Rev. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Linehan, W.M.; Walther, M.M.; Zbar, B. The Genetic Basis of Cancer of the Kidney. J. Urol. 2003, 170, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Krajewska, M.; Krajewski, S.; Epstein, J.I.; Shabaik, A.; Sauvageot, J.; Song, K.; Kitada, S.; Reed, J.C. Immunohistochemical Analysis of Bcl-2, Bax, Bcl-X, and Mcl-1 Expression in Prostate Cancers. Am. J. Pathol. 1996, 148, 1567–1576. [Google Scholar]
- Wang, Q.; Zhang, L.; Yuan, X.; Ou, Y.; Zhu, X.; Cheng, Z.; Zhang, P.; Wu, X.; Meng, Y.; Zhang, L. The Relationship between the Bcl-2/Bax Proteins and the Mitochondria-Mediated Apoptosis Pathway in the Differentiation of Adipose-Derived Stromal Cells into Neurons. PLoS ONE 2016, 11, e0163327. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Shin, J.; Choi, H.; Jo, A.; Pan, S.; Bae, D.; Lee, Y.; Choi, C. Cynanchum Wilfordii Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia by Regulating 5α-Reductase and Androgen Receptor Activities in a Rat Model. Nutrients 2017, 9, 1070. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Androgen-Dependent Tissue Weight/Body Weight (%) | |||||
|---|---|---|---|---|---|
| Treatment | Ventral Prostate | Seminal Vesicles | Glans Penis | LABC | Cowper’s Gland |
| Control | 0.041 ± 0.005 | 0.121 ± 0.031 | 0.027 ± 0.004 | 0.183 ± 0.014 | 0.010 ± 0.001 |
| Flutamide 3 mg/kg | 0.011 ± 0.001 *** | 0.017 ± 0.006 *** | 0.013 ± 0.003 *** | 0.099 ± 0.009 *** | 0.004 ± 0.001 *** |
| HT080 100 mg/kg | 0.030 ± 0.009 | 0.089 ± 0.016 | 0.026 ± 0.004 | 0.170 ± 0.011 | 0.009 ± 0.001 |
| HT080 300 mg/kg | 0.035 ± 0.014 | 0.091 ± 0.038 | 0.030 ± 0.003 | 0.165 ± 0.013 | 0.010 ± 0.003 |
| HT080 1000 mg/kg | 0.039 ± 0.038 | 0.125 ± 0.031 | 0.031 ± 0.004 | 0.173 ± 0.034 | 0.010 ± 0.001 |
| HT080 3000 mg/kg | 0.039 ± 0.008 | 0.127 ± 0.042 | 0.031 ± 0.003 | 0.186 ± 0.035 | 0.010 ± 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.; Tran, P.; Yin, J.; Song, J.; Kim, H. Cinnamomum cassia and Rosa laevigata Mixture Improves Benign Prostatic Hyperplasia in Rats by Regulating Androgen Receptor Signaling and Apoptosis. Nutrients 2023, 15, 818. https://doi.org/10.3390/nu15040818
Kim M, Tran P, Yin J, Song J, Kim H. Cinnamomum cassia and Rosa laevigata Mixture Improves Benign Prostatic Hyperplasia in Rats by Regulating Androgen Receptor Signaling and Apoptosis. Nutrients. 2023; 15(4):818. https://doi.org/10.3390/nu15040818
Chicago/Turabian StyleKim, Myunghee, Phuong Tran, Jun Yin, Jungbin Song, and Hocheol Kim. 2023. "Cinnamomum cassia and Rosa laevigata Mixture Improves Benign Prostatic Hyperplasia in Rats by Regulating Androgen Receptor Signaling and Apoptosis" Nutrients 15, no. 4: 818. https://doi.org/10.3390/nu15040818
APA StyleKim, M., Tran, P., Yin, J., Song, J., & Kim, H. (2023). Cinnamomum cassia and Rosa laevigata Mixture Improves Benign Prostatic Hyperplasia in Rats by Regulating Androgen Receptor Signaling and Apoptosis. Nutrients, 15(4), 818. https://doi.org/10.3390/nu15040818