1. Introduction
With advancements in economic growth and changes in lifestyles, the number of people suffering from hyperuricemia is increasing annually worldwide. Approximately 20.2% and 20.0% of American males and females, respectively, are estimated to suffer from hyperuricemia [
1]. As the largest developing country, the incidence of hyperuricemia in China has been reported to be up to 17.4%, with the age of onset gradually decreasing [
2]. Regardless of the presence of symptoms, hyperuricemia has tremendous harmful effects on the body, posing a challenge to patients and clinicians. In addition to being the main hazard for gout, hyperuricemia also increases the risk of other diseases [
3], such as diabetes and chronic kidney disease. Furthermore, a recent Irish study illustrated that compared to the normal population, blood uric acid of males and females above 535 μM/L and 416 μM/L, respectively, decreased their median survival years by 11.7 and 6 years, respectively [
4]. Therefore, there is an urgent need to identify effective measures to prevent and treat hyperuricemia.
As an end product of purine metabolism, uric acid plays an important antioxidative role similar to Vitamin C [
5]. Hyperuricemia occurs as a result of purine metabolism dysregulation and has a complex pathogenesis. The risk factors of hyperuricemia include genetics, nutritional status, sleep and stress, which consequently induce inflammation, oxidative stress and insulin resistance in the body. Moreover, it is accompanied by liver and kidney damage and toxic epidermal necrolysis [
6]. Under these circumstances, the body compensates by increasing uric acid levels to counteract the damage caused by the metabolic disorder.
Current treatments for hyperuricemia include xanthine oxidase inhibitors (allopurinol or febuxostat), which decrease uric acid production, and uricosurics (probenecid), which increase uric acid excretion. Additionally, a low-purine diet has been recommended for lowering blood uric acid levels. Although this diet is safe, the effects of dietary interventions on hyperuricemia are limited. A recent meta-analysis on the effects of diet and genetics on blood uric acid concentrations in Caucasians in New Zealand demonstrated that 63 types of food combinations accounted for only 4.29% of the variation in blood uric acid concentrations. Contrastingly, 23.8% and 40.3% variations in blood uric acid concentrations in males and females, respectively, were due to genetic factors [
7]. Therefore, simply reducing uric acid concentrations does not deter the development of hyperuricemia. Furthermore, there is a need to elucidate the pathogenesis of hyperuricemia and identify an efficient dietary intervention using natural products to improve overall body metabolism.
Overproduced uric acid or reduced uric acid excretion are the two primary causes of hyperuricemia. The kidneys excrete two-thirds of the uric acid, while the intestines excrete a third [
8]. Excretory disorders related to the kidney are usually associated with the regulation of molecular signals, such as insulin resistance, inflammation, oxidative stress and cell damage [
5]. Low-grade chronic systemic inflammation can directly lead to kidney damage and then affect the kidney’s uric acid-related transporters expression, ultimately affecting the excretion of uric acid [
9]. Additionally, studies show that high uric acid impairs mitochondrial function and produces reactive oxygen species, which activates the inflammatory bodies of the NOD-like receptor (NLR) family containing pyrimidine domain 3 (NLRP3). This cascade further aggravates kidney injury by secreting interleukin-1β (IL-1β) [
10]. Moreover, excess uric acid enters the cells and becomes pro-oxidative, subsequently causing oxidative stress, aggravating kidney dysfunction and improving insulin resistance, thus forming a vicious circle. Therefore, reducing oxidative stress and low-grade chronic inflammation in the kidney is vital for the excretion of uric acid via the kidney. Various metabolites produced during this pathophysiological process are ultimately excreted by the kidneys through urine. Therefore, exploring urinary metabolites in hyperuricemia could provide a more comprehensive elucidation of its pathogenesis.
Recently, the gut microbiota has been reported to influence hyperuricemia, especially via the gut–kidney axis [
11]. Then, the kidneys are damaged due to hyperuricemia, and uric acid and uremic toxins accumulate in the blood [
12]. Moreover, low-grade inflammation produces proinflammatory cytokines. Elevated uric acid levels, uremic toxins and inflammatory cytokines negatively affect gut microbiota homeostasis. Dysbiosis of the gut microbiota increases intestinal permeability, which allows bacteria and intestinal metabolites, such as lipopolysaccharide (LPS), to be transported out of the intestine. LPS forms a complex with its CD14 receptor and is detected by Toll-like receptor 4 (TLR4), which induces chronic low-grade inflammation and thereby aggravates kidney injury [
13]. The entire process forms a vicious cycle and increases the risk of hyperuricemia. Furthermore, gut dysbacteriosis increases the abundance of the xanthine oxidase gene-related microbiota and decreases the abundance of allantoinase gene-related flora, resulting in elevated uric acid levels in the intestinal tract. Therefore, this study aims to explore intervening substances that act on the gut–kidney axis.
The ocean is a uniquely rich source of bioactive peptides. In studies on peptides associated with hyperuricemia, pelagic fishes, such as tuna and bonito, have been found to migrate tens of thousands of kilometres at high speeds without causing an acid build-up in the body. This phenomenon was attributed to the presence of an important dipeptide, anserine, in the body, which has currently become a new target for dietary intervention in hyperuricemia [
14]. Anserine is a multifunctional and highly stable histidine carnosine-like dipeptide found in fish skeletal muscles. In human clinical trials, anserine has been shown to reduce blood glucose and inflammation and elevate kidney functions [
15]. Additionally, it improved gut microbiota disorders caused by hyperuricemia in mice [
16]. However, the effect of anserine on host metabolism remains unclear and the alleviation of hyperuricemia via the gut–kidney axis remains to be investigated.
Thus, this study establishes a rat model for hyperuricemia using potassium oxycyanate and yeast and verifies the ameliorating effect of anserine on hyperuricemia. It also explores the mechanism of anserine-related amelioration of hyperuricemia through gut microbiota and metabolites using a combined ultraperformance liquid chromatography–tandem mass spectrometry (UPLC–MS) and macrogenomic analysis. Thus, this study aims to explore novel ideas for preventing and treating hyperuricemia.
2. Materials and Methods
2.1. Materials and Reagents
A rodent regular diet was obtained from Beijing Weitong Lihua Laboratory Animal Technology Co., Housed in a clean-grade animal house at the Peking University Health Science Center and the provided nutrients met the needs for rodent growth and development per GB 14924.3-2010. The diet for hyperuricemic rats comprised a normal diet combined with 4% potassium oxonate and 20% yeast, which was obtained from Beijing BotaiHongda Biotechnology Co., Ltd. Anserine was obtained from Sinopharm Holding Starshark Pharmaceutical (Xiamen) Co., Ltd. Allopurinol was obtained from Beijing Balinwei Technology Co., Ltd.
Primary antibodies against TLR4, myeloid differentiation factor88 (MyD88), nuclear factor E2-related factor 2 (Nrf2) antibody, NLRP3 inflammation, ATP-binding cassette, subfamily G, member 2 (ABCG2), matrix metallopeptidase 2 (MMP2), matrix metallopeptidase 2 (MMP9), β-actin and secondary antibodies were obtained from Abcam Co., Ltd. The primary antibody nuclear factor-κB (NF-kB) was obtained from CST Co., Ltd., whereas that against urate transporter 1 (URAT1) was obtained from Proteintech Biotech Co., Ltd. The primary antibodies against glucose transporter 9 (GLUT9) and the tissue inhibitor of metalloproteinase1 (TIMP-1) were obtained from Novus Biologicals Co., Ltd. and BOSTER Co., Ltd., respectively.
2.2. Animal Treatment
A total of 60 male Sprague–Dawley rats (180–220 g) were obtained from the Beijing Vital River Laboratory Animal Technology Co., Ltd. The animals were housed in a climate-controlled room at 25 ± 1 °C with a 12/12 h dark/light cycle and free access to sterile water and standard feed. All laboratory procedures were reviewed and approved by the Institutional Animal Care and Use Committee of Peking University (Approved No. 2019PHE017) on the 5th May 2019 and conformed to the Guidelines for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, Revised 1996).
After four days of acclimatization, the rats were randomly divided into six groups (n = 10): normal control group (NC group), hyperuricemia group (HUA group), allopurinol group (Allo group, 10 mg/kg·bw allopurinol), three anserine groups (Ans1, Ans10 and Ans100 groups were treated with 1 mg/kg·bw, 10 mg/kg·bw and 100 mg/kg·bw anserine, respectively). During the six weeks of intervention, the general conditions of experimental animals, including health and behaviour, were observed daily, and food intake, water intake, urine volume and weight gain were monitored weekly.
2.3. Plasma and Urine Biochemical Analysis
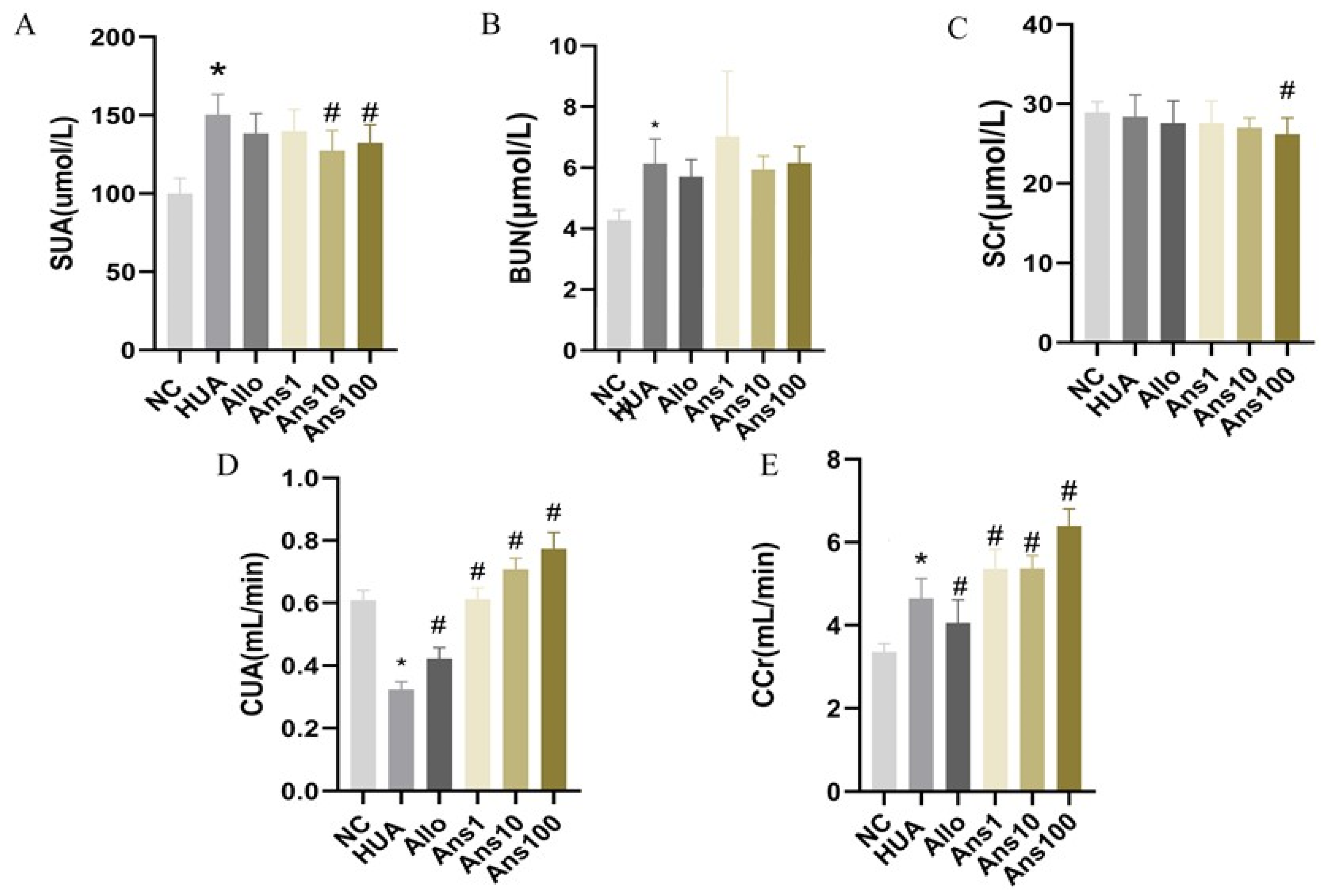
Urine was collected for 24 h after six weeks of intervention and the volume was measured. Following centrifugation at 3000 r/min for 10 min, the supernatant was collected and stored at −80 °C. Subsequently, blood samples were collected and centrifuged at 3000 r/min for 10 min at 4 °C after the rats were sacrificed. The concentrations of serum uric acid (SUA), serum creatinine (SCr), blood urea nitrogen (BUN), urinary uric acid (UUA), urinary creatinine (UCr), alanine aminotransferase (ALT), aspartate aminotransferase (AST), albumin (ALB), globulin (GLB), albumin/globulin (A/G) and total protein (TP) were measured using an automatic biochemical instrument (AU480, Japan Olympus Corporation). Uric acid clearance (CUA) and creatinine clearance (CCr) were calculated using the following formulas:
2.4. Serum and Liver Xanthine Oxidase (XOD) and Adenosine Deaminase (ADA) Assay
Serum XOD activity was measured using the colourimetric method. After the liver was excised, 1 g of liver tissue was added to 9 mL of 0.9% normal saline to prepare 10% liver homogenate using mechanical homogenizers. The homogenate was centrifuged at 3000 r/min for 10 min and then 100 μL of the 10% liver homogenate supernatant was collected to evaluate liver XOD activity using colourimetry. A total of 50 μL of the liver homogenate supernatant was used to determine ADA activity. ADA generates ammonia by hydrolysing adenosine, which is then used to calculate ADA activity.
2.5. Histopathological Evaluation
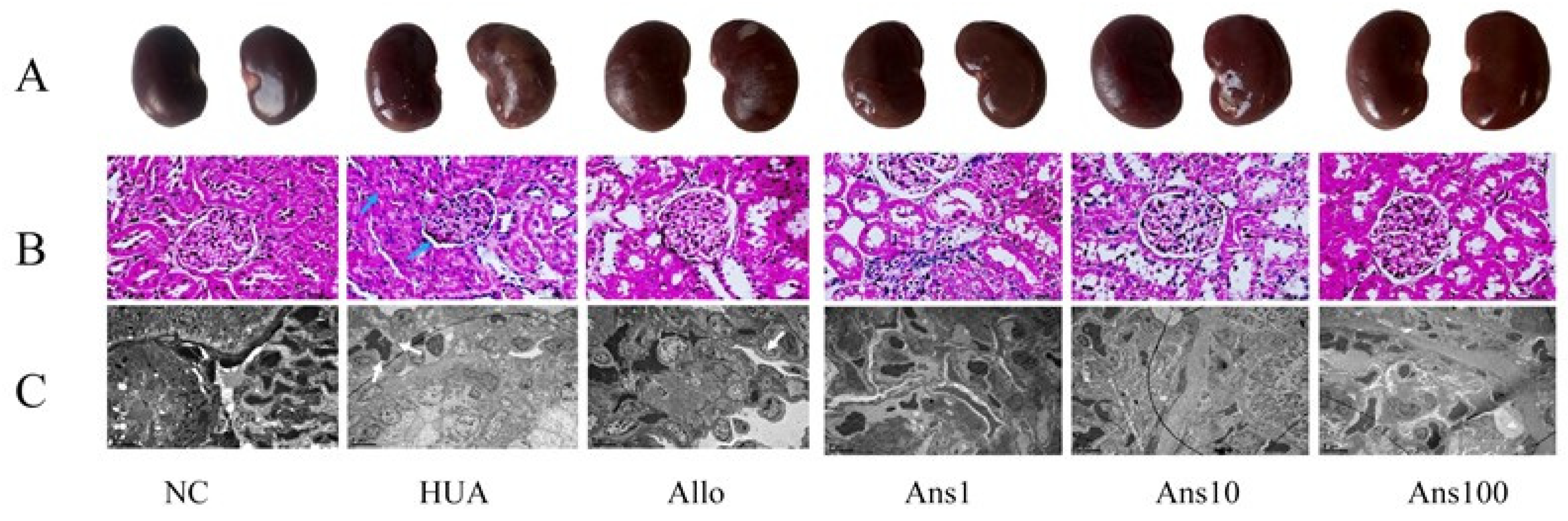
The rats were sacrificed after the experiment. Then, their right kidneys were fixed with 4% paraformaldehyde, and paraffin sections were made by a series of techniques such as dewaxing, hematoxylin staining, eosin staining, dehydration and sealing. Finally, the tissue was examined under an optical microscope (Olympus BX43) and the image was recorded. The remaining half of the right kidney was fixed with 4% glutaraldehyde at a size of 1 mm × 1 mm × 1 mm, and then underwent a series of processes such as dehydration, transparency, wax immersion, embedding and sectioning to obtain electron microscopic sections for observation under an electron microscope (JEM1400).
2.6. Western Blotting
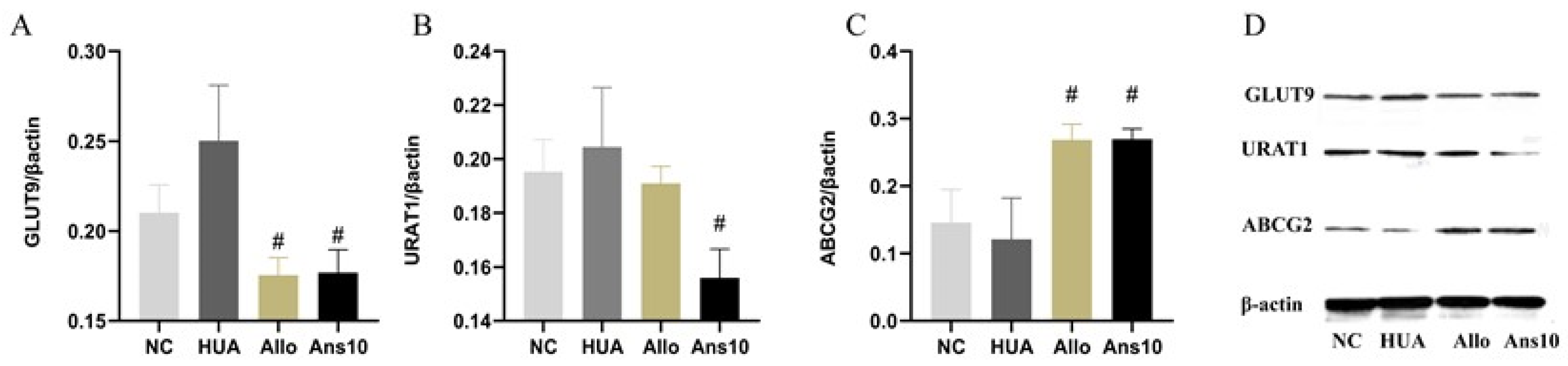
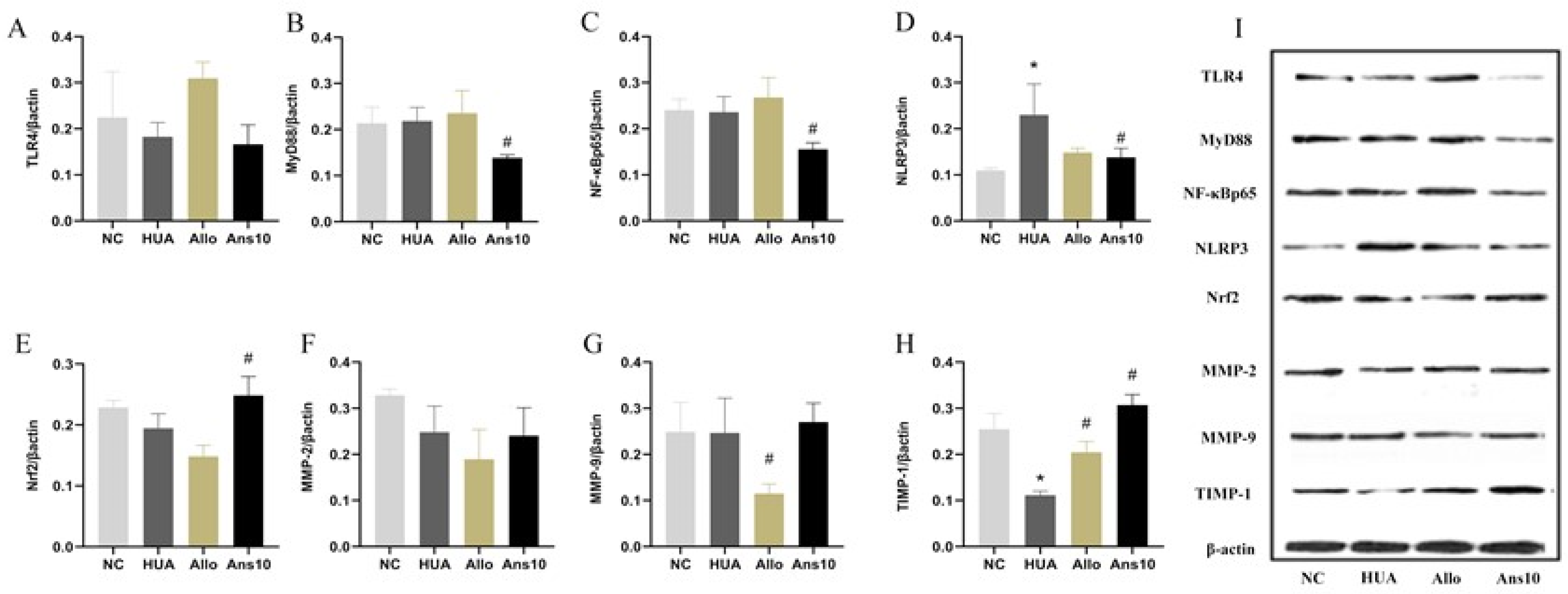
The expressions of GLUT9, URAT1, ABCG2, TLR4, MyD88, NF-κB, NLRP3, Nrf2, MMP-1, MMP-2 and TIMP-1 proteins in the renal tissue were detected using Western blotting. Commercial kits (Beijing Prily Gene Technology Co., Ltd. C1053) were used to extract total protein from kidney tissue, and BCA protein detection kits (Beijing Prily Gene Technology Co., Ltd. P1511) were used to quantify protein concentrations. After protein separation using 12% sodium dodecyl-sulphate polyacrylamide gel electrophoresis, an Immobilon-P transfer membrane (Millipore, Germany, IPVH00010) was used for transferring the protein. Following this, the membrane was sealed with 5% nonfat milk, incubated overnight at 4 °C with primary antibodies, and then incubated with secondary antibodies conjugated with horseradish peroxidase. Colour development using ECL Chemiluminescence Chromogenic solution (PerkinElmer, America, NEL105001EA) was performed. The grayscale was calculated and normalised to the values of β-actin.
2.7. Untargeted Urine Metabolomic Analysis
To 100 μL of urine sample in an Eppendorf tube, 400 μL of 80% methanol aqueous solution (Thermo Fisher, Waltham, MA, USA, 67-56-1) was added. The sample was vortexed, placed in an ice bath for 5 min and centrifuged at 15,000 r/min for 20 min at 4 °C. A quantity of the supernatant was diluted with MS-grade water to obtain a methanol content of 53%. This sample was then centrifuged at 15,000 r/min for 20 min at 4 °C and the supernatant was analysed using a UPLC–MS. Metabolomics workflow was based on the R language MetaboAnalystR package. First, the sample data were subjected to quality control and batch correction, then the sample data were standardized, and finally, the metabolite content was counted. Metabolites that differed between groups were identified using an orthogonal partial least squares discriminant method.
2.8. Metagenomic Sequencing of Gut Microbiota
DNA was extracted from stool samples using the CTAB method. After the DNA samples were qualified, the library was constructed. Qubit 2.0 was used for preliminary quantification, and then, an Agilent 2100 was used for the detection of inserted fragments from the library. Using real-time polymerase chain reaction, the effective concentration of the library could be accurately quantified after the insert size reached the expected size. Following library detection, different libraries were pooled into a flow-through pool based on effective concentration and target data volume. After cBOT clustering, the Illumina PE150 (2 × 150) high-throughput sequencing platform was used for sequencing. KneadData software was used for raw data quality control (Trimmom-based) and de-hosting (Bowtie2-2-based). Before and after KneadData, FastQC was used to verify the justification and effectiveness of the quality control. Furthermore, Kraken2 was used to analyse the species composition and diversity information of samples, whereas Bracken was used for predicting species relative abundances. The identification of microbial genera and species that differed between the six groups was performed using LEfSe biomarker mining analysis (microorganisms with LDA > 2 by default). To functionally annotate genes, we used HUMAnN2 software to compare the sequences after quality control (based on DIAMOND) and host removal with the protein database (UniRef90), further filtering out reads that failed to be compared. Based on the mapping between the ID of UniRef90 and the ID of the KEGG and EC functional databases, the relative abundance of the corresponding functions of each functional database was calculated.
2.9. Statistical Analysis
2.9.1. Statistical Analysis of Non-Omics Data
One-way analysis of variance (ANOVA) was calculated using SPSS 25.0 software; variables were transformed if they did not meet the requirements of normality for variance. If the transformed variables still did not meet the requirements, a nonparametric test was used for statistical analysis. The experimental and control groups were compared using the least significant difference (LSD) method, and a difference of p < 0.05 was considered to be statistically significant.
2.9.2. Statistical Analysis of Omics Data
The omics data were analysed using R software (version 4.1.1). Principal coordinates analysis (PcoA) and principal components analysis (PCA) were used for dimension reduction analysis to show the degree of variation among the samples. Binary unpaired samples were tested for significance using the rank-sum test, and multiple groups were compared using the Kruskal–Wallis rank-sum test.
2.9.3. Multiomics Association Analysis
The correlation between gut microbiota and metabolites was analysed using the Pearson correlation coefficient method, with a significance threshold of p < 0.05.
4. Discussion
This study used yeast combined with potassium oxalate to establish a rat model with hyperuricemia. Anserine was found to significantly reduce blood uric acid levels and improve liver and kidney damage, indicating that anserine had a preventative effect on hyperuricemia. Additionally, the changes in the gut microbiota structure and function as well as host metabolism, which were induced by hyperuricemia, were partially reversed by anserine. Moreover, the underlying mechanism of anserine ameliorating hyperuricemia was explored using integrated macrogenomics and metabolomics for the first time, to the best of our knowledge, in this study.
It is reported that, in general, bonito, bigeye tuna, and southern tuna contain about 1070 mg, 1260 mg and 636 mg of anserine per 100 g of fish flesh, respectively [
17]. We learned that the doses of anserine used in previous animal experiments ranged from 2 mg/kg·bw to 80 mg/kg·bw [
18,
19], so we chose the 1 mg/10 mg/100 mg kg·bw as the low/medium/high doses of anserine intervention in our experiment. Our results did show that medium doses of anserine were the most effective in improving hyperuricemia. Consequently, we speculate that the effect of anserine in improving hyperuricemia is dose-dependent. Therefore, in subsequent experiments using Western blotting to detect the expression of uric acid-related transporter proteins and proteins associated with renal inflammation, oxidative stress, and cellular damage in the kidney, we only examined kidney samples from the mid-dose group of the goose-peptide intervention.
Initially, the effect of anserine on uric acid production and excretion was investigated. Anserine showed no obvious effects on ADA and XOD enzymes, which are closely related to uric acid production. However, anserine ameliorated liver damage induced by hyperuricemia. Then, the effect of anserine on kidney uric acid excretion was investigated. URAT1, GLUT9 and ABCG2 expression changes indicated that anserine reduced serum uric acid levels by inhibiting uric acid reabsorption and promoting uric acid excretion. The changes in CUA and CCr, two indicators of kidney function, also implied that anserine improved kidney function, which is consistent with the morphological changes observed in the kidneys. It is noteworthy that, in our experiments, the levels of CCr were elevated in the HUA group. We assume this is very likely because when the blood creatinine in hyperuricemic rats started to rise, the body compensated by increasing the renal creatinine clearance to keep the body’s creatinine level at a steady state. Additionally, the timing of the measurement also has some influence on the results. In short, this phenomenon is very interesting and deserves further in-depth study.
We further investigated the effect of anserine on kidney injury caused by hyperuricemia by evaluating the effect of anserine on kidney inflammation, oxidative stress and cellular injuries. The TLR4/MyD88/NF-κB and NLRP3 proteins are reported to be the major signalling pathways closely associated with renal inflammation caused by hyperuricemia [
20]. Contrarily, Nrf-2 neutralises the activation of cellular oxidative stress and ameliorates kidney injury by inhibiting NF-κB expression [
21]. Moreover, MMP2 and MMP9 are reported to cleave collagen IV in the basement membrane of cell bands and have activity in kidney tissue [
22]. Furthermore, TIMP-1 inhibits the activities of most MMPs, thus improving cell damage [
23]. Given that anserine decreased the expression of TLR4/MyD88/NF-κB and NLRP3 and elevated Nrf2 and TIMP1 in hyperuricemic rats, we speculate that anserine improves overall kidney function by decreasing inflammation, oxidative stress and cellular damage in hyperuricemia. Anserine also regulates the expressions of uric acid-related transporters, thereby reducing serum uric acid levels.
Increasing evidence suggests that gut microbiota balance is closely related to metabolic disorders, and patients with hyperuricemia have a different intestinal microbiota structure from normal individuals [
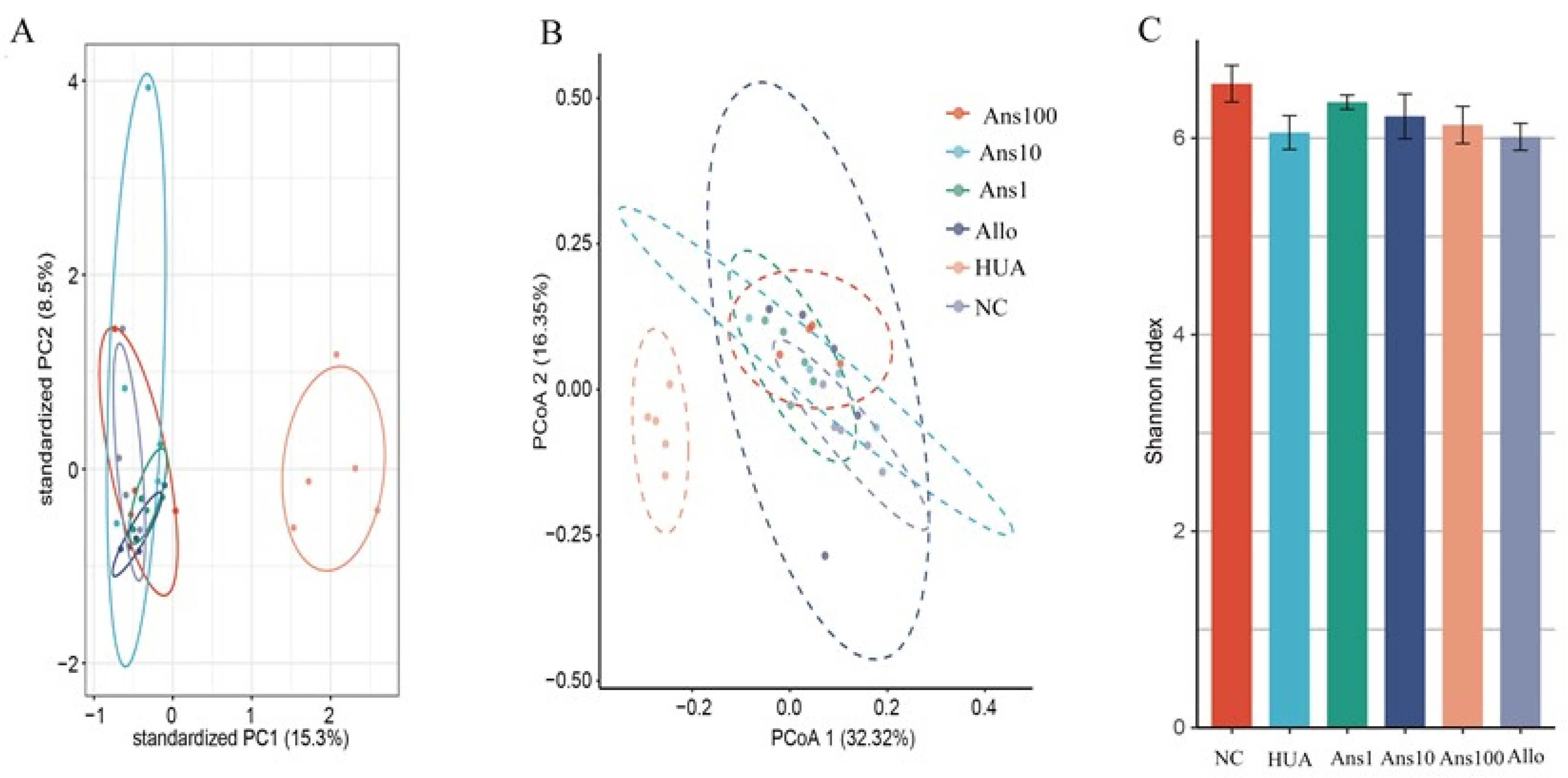
24]. Abnormally high levels of uric acid in the blood enter the intestine and affect the steady state of the intestinal flora, thereby affecting the intestinal metabolism of uric acid and consequently aggravating hyperuricemia. Therefore, we analysed the effect of anserine on the changes in intestinal flora structure caused by hyperuricemia. Our study showed a decrease in the intestinal microbial diversity of hyperuricemic rats, which is consistent with previous studies [
24]; however, this was reversed on anserine intervention.
Uric acid is the end product of purine metabolism; moreover, intestinal flora has also been shown to play an essential role in purine oxidative metabolism. For example,
Escherichia coli in the human gut produces XOD to influence the production of uric acid [
25]. The
Lactobacillaceae family inhibits the growth of
E. coli by secreting reuterin [
26], indirectly inhibiting uric acid accumulation. Additionally,
Lactobacillus can synthesize various UA metabolic enzymes, such as uricase, allantoinase and allantoicase, which can decompose uric acid into 5-hydroxyisothreonate, allantoin, allantoate and finally degrade it into urea [
27]. Similarly, the
Clostridiaceae family also degrades uric acid [
28].
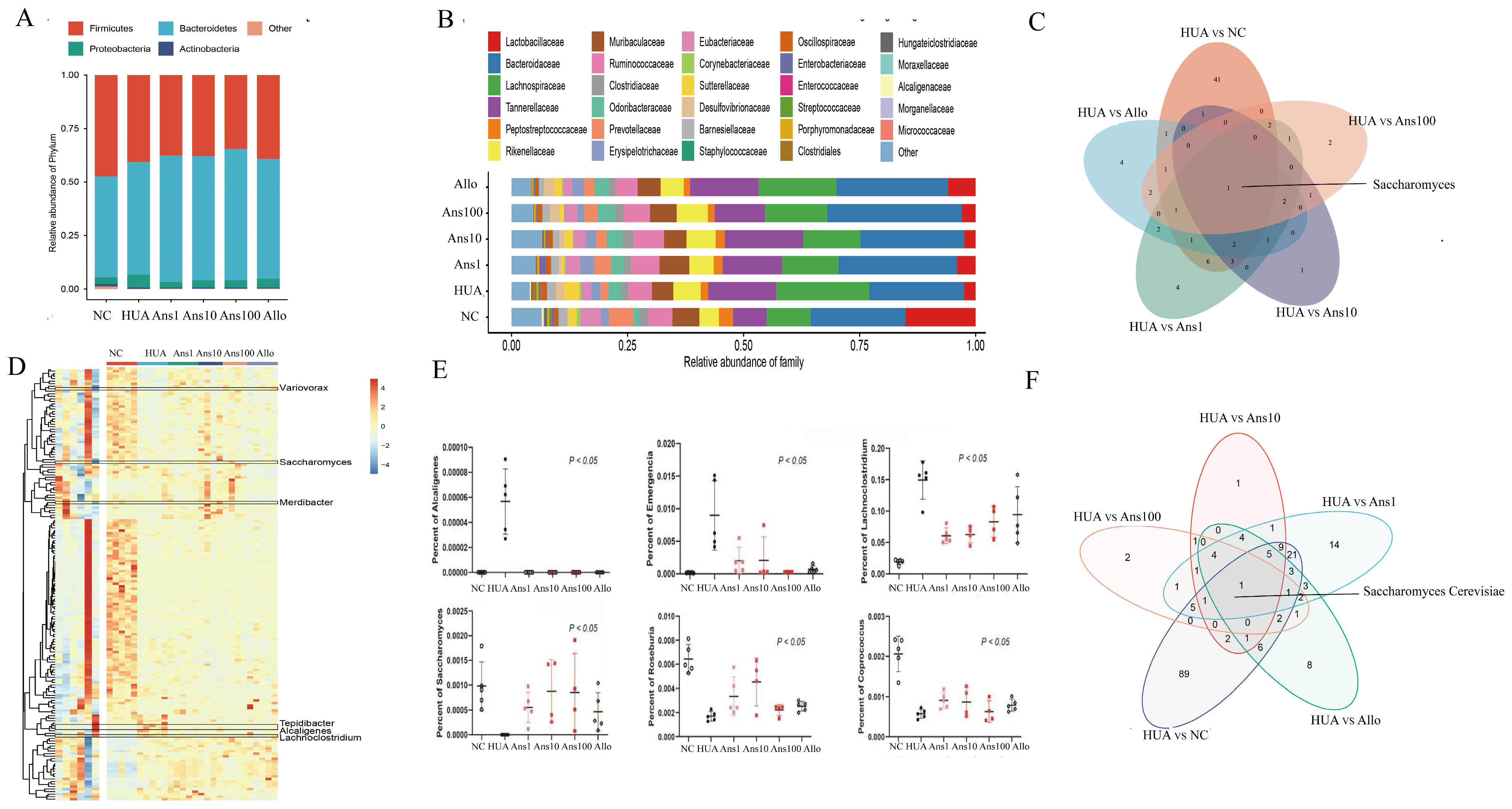
Saccharomyces cerevisiae is a fungus that secretes urate oxidase, which can catalyse uric acid oxidation and plays an essential role in the purine degradation pathway, thereby preventing uric acid accumulation [
29]. This study showed that the abundance of
Lactobacillaceae,
Clostridiaceae family and
Saccharomyces cerevisiae was reduced in hyperuricemic rats but elevated after the anserine intervention, suggesting the preventive effect of anserine was partially due to the changes in some specific microbiota.
A dysregulated gut microbiota is accompanied by imbalanced intestinal metabolites, such as trimethylamine, short-chain fatty acids (SCFAs) and LPS, which are considered mediators between the intestinal microbiome and their human hosts [
30]. Anserine increases the abundance of
Roseburia and
Coprococcus in hyperuricemic rats, which are crucial in SCFA generation. SCFAs regulate gut microbiota homeostasis, repair intestinal permeability and are beneficial to kidney function [
31]. Moreover, butyrate, a major SCFA in the intestine, is reported to be increased in a
Lactobacillaceae-enriched environment [
32]. Therefore,
Lactobacillaceae is speculated to not only participate in purine metabolism but also play a role in increasing butyrate levels in the intestinal tract. Additionally, gut microbiome dysbiosis can cause the excretion of LPS from the cell walls of Gram-negative bacteria, and the inflammation in the liver and kidney is further activated by the excreted LPS entering the bloodstream through a disrupted gut barrier [
33]. Furthermore, members of the Gram-negative
Proteobacteria phylum,
Alcaligenes genus and
Lachnoclostridium genus were increased in the HUA group but were reduced on anserine intervention. Additionally, a Proteobacterial strain has also been shown to enhance intestinal nitrogen fixation [
34], wherein, nitrogen is converted to ammonia. Notably, excess ammonia entering the host’s circulatory system through the intestinal barrier can aggravate kidney damage. Additionally, we observed the
Emergencia timonensis genus was more enriched in the HUA group than in other groups. Furthermore,
Emergencia timonensis, a potential key bacterium for the conversion of carnitine to trimethylamine N-oxide (TMAO), is also a toxin that can aggravate kidney damage [
35]. This study indicated that anserine alleviated hyperuricemia owing to its ability to maintain the balance in the composition of the intestinal microbiota (the increase in beneficial bacteria and the decrease in pathogenic bacteria). Moreover, it also promotes purines and uric acid catabolism, regulates intestinal epithelial cell proliferation, reduces chronic inflammation and improves uric acid excretion.
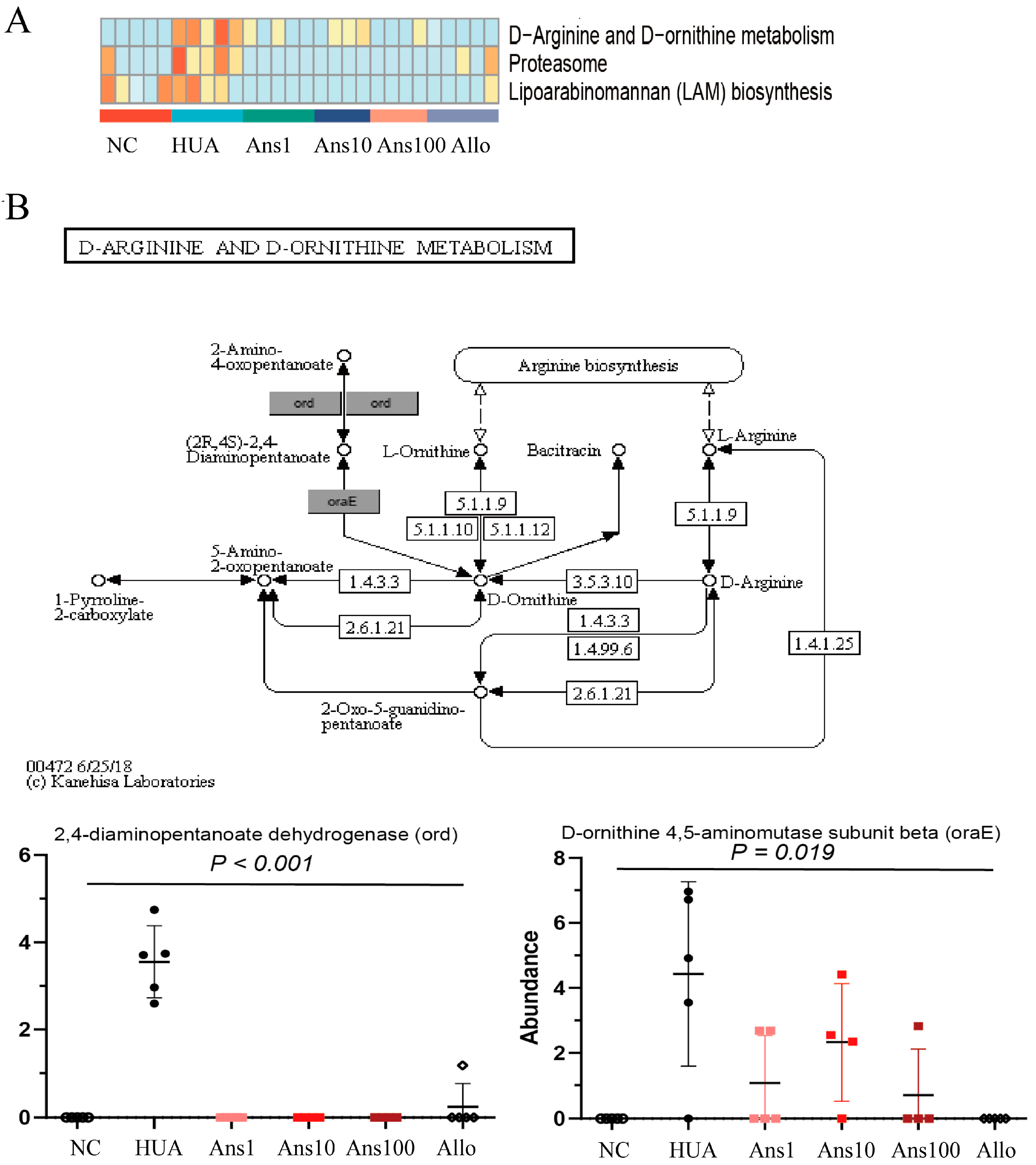
The intestinal microflora greatly affects the health of the host by regulating its metabolic function. In this study, six metabolic pathways were altered in the HUA group compared to the NC group, three of which were elevated and the other three were decreased; however, anserine intervention reversed these changes. For example, D-Arginine and D-ornithine metabolism pathways were significantly enriched by hyperuricemia but anserine supplementation reversed the change, which was verified by the changes in two key proteins in this pathway, namely 4-diaminopentanoate dehydrogenase and D-ornithine 4,5-aminomutase subunit beta. The D-Arginine and D-ornithine metabolism are related to the urea cycle, indicating that more urea is metabolized in the intestine to produce ammonia in hyperuricemia. The disturbance of the gut microbiota combined with the reduction in beneficial metabolites such as SCFAs increases the permeability of the intestine, which increases the ammonia levels entering the circulation system, thereby aggravating liver and kidney function.
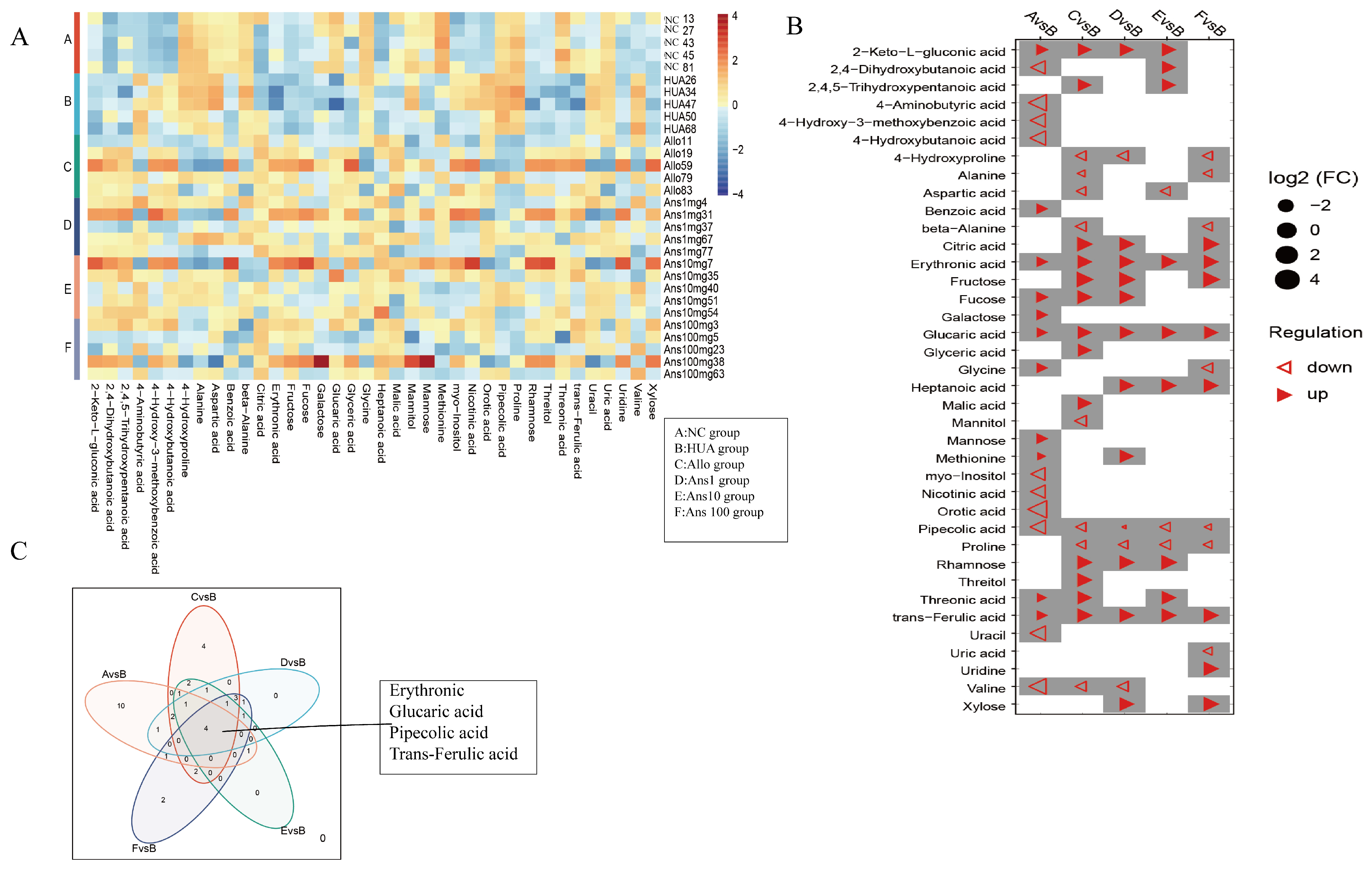
Metabolic profiling of host biofluids provides profound insights into the gut microbiota’s impact on host health/disease status, therefore exploring differential urinary metabolites aids in identifying the causative agent rather than the presence of the metabolite [
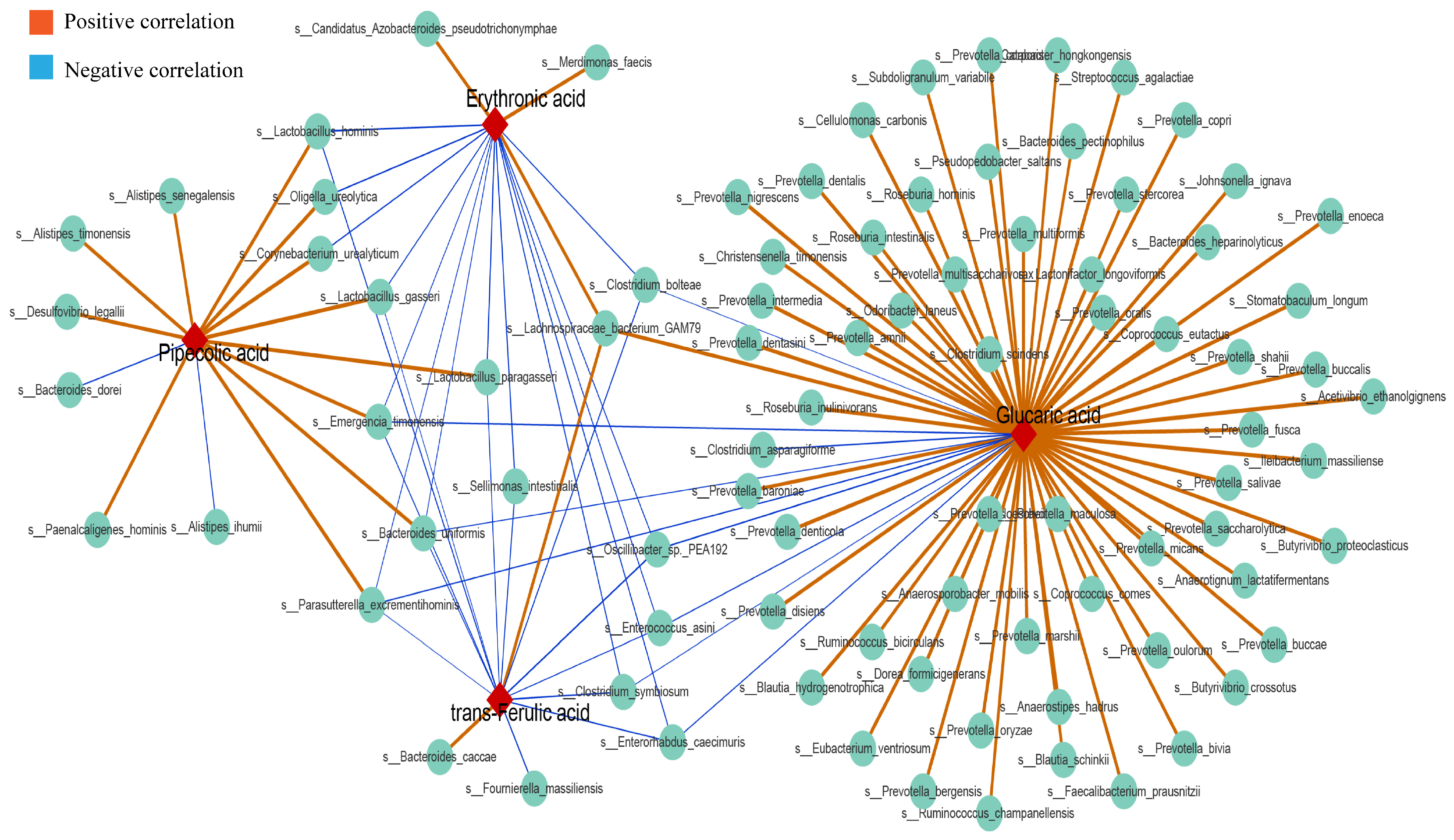
36]. By comparing urinary metabolites in the HUA group with the different doses of the anserine group, Allo and NC groups, we identified erythronic acid, glucaric acid, pipecolic acid and trans-ferulic acid as the four common differential metabolites. This suggests that these four metabolites and their associated metabolic pathways play a critical role in the pathogenesis and amelioration of hyperuricemia. Erythronic acid is related to mitochondrial dysfunction in transaldolase deficiency [
37], highlighting its role in mediating energy metabolism in humans. D-gluconic acid has toxin-reducing and antioxidant abilities, wherein it can improve diabetic kidney tubular damage by inhibiting inositol oxygenase, preventing mitochondrial damage and apoptosis and reducing oxidative stress through the ascorbic acid and aldehyde metabolic pathway, thereby improving kidney function [
38]. Moreover, ferulic acid has been shown to lighten oxidative stress through the activation of the AMPK signalling pathway in vitro [
39]. Pipecolic acid is an intermediate in the lysine degradation pathway, with an enhanced lysine degradation pathway indicating enhanced levels of oxidative stress in the host [
40]. Thus, by enhancing the levels of erythronic acid, glucaric acid and trans−ferulic acid and decreasing the levels of pipecolic acid in hyperuricemic rats, anserine exerts an anti-hyperuricemia effect by improving energy metabolism and reducing oxidative stress and inflammation. Notably,
Parasutterella excrementihominis,
Emergencia timonensis and
Bacteroides uniformis were associated with these four metabolites. As
Parasutterella excrementihominis and
Emergencia timonensis are positively associated with pipecolic acid but negatively associated with erythronic acid, glucaric acid and trans-ferulic acid, we speculated that anserine primarily reduced the abundance of
Parasutterella excrementihominis and
Emergencia timonensis to exert an ameliorating effect on kidney injuries.
Notably, methionine was associated with the highest number of differential genera and species; however, the
Saccharomyces genus was only correlated with methionine. Methionine produces strong antioxidative metabolites such as glutathione, cysteine and sulphate through the trans-sulphuration pathway. Previous studies have demonstrated the ameliorative effect of methyl and S-adenosylmethionine produced by the methionine cycle on systemic inflammation and liver damage [
41]. The methionine cycle is widely active in
Saccharomyces cerevisiae [
42]. This suggests that
Saccharomyces cerevisiae could be a target probiotic for anserine to improve hyperuricemia. Additionally, the anserine group exhibited significantly higher levels of two differential metabolites (fructose and xylose) in the starch and sucrose metabolism pathway than the HUA group. Starch and sucrose metabolic pathways are directly related to the development of diseases involved in energy metabolism and insulin resistance, such as obesity, diabetes and kidney tubular dysfunction [
43].



 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








