
The Associations between Multiple Essential Metal(loid)s and Gut Microbiota in Chinese Community-Dwelling Older Adults

Abstract
:1. Introduction
2. Material and Methods
2.1. Data Source and Population
2.2. Measurement of Urinary EMs
2.3. 16S rRNA Gene Sequencing and Data Analyses
2.4. Outcome Assessments
2.5. Covariates
2.6. Statistical Analysis
3. Results
3.1. Population Characteristics
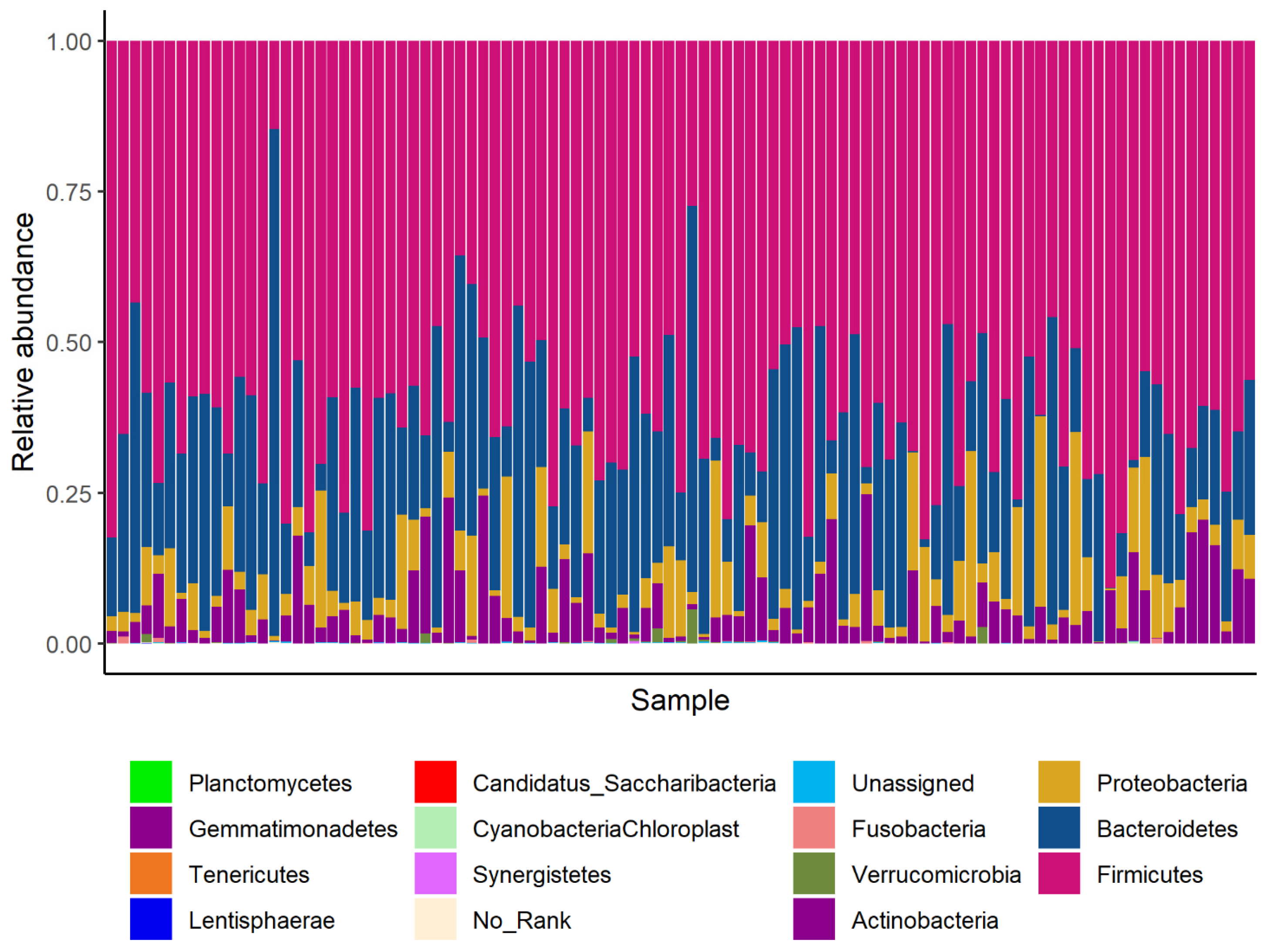
3.2. Gut Microbiota Compositions
3.3. Distributions of Urinary EM Concentrations
3.4. Associations of Single EMs with α-Diversity and β-Diversity
3.5. Associations between Single EMs and Specific Taxons
3.6. The Cumulative Effects of EMs on the Compositions of the Gut Microbiota
3.7. The Interaction Effects of EMs on the Composition of the Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cell. Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Xing, Y.; Song, X.; Qian, Y. The impact of lung microbiota dysbiosis on inflammation. Immunology 2020, 159, 156–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, P.W.; Claesson, M.J. Gut microbiota: Changes throughout the lifespan from infancy to elderly. Int. Dairy, J. 2010, 20, 281–291. [Google Scholar] [CrossRef]
- Biagi, E.; Franceschi, C.; Rampelli, S.; Severgnini, M.; Ostan, R.; Turroni, S.; Consolandi, C.; Quercia, S.; Scurti, M.; Monti, D.; et al. Gut Microbiota and Extreme Longevity. Curr. Biol. 2016, 26, 1480–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Jazwinski, S.M. The Gut Microbiota and Healthy Aging: A Mini-Review. Gerontology 2018, 64, 513–520. [Google Scholar] [CrossRef]
- Kong, F.; Hua, Y.; Zeng, B.; Ning, R.; Li, Y.; Zhao, J. Gut microbiota signatures of longevity. Curr. Biol. 2016, 26, R832–R833. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, P.W.; Jeffery, I.B. Gut microbiota and aging. Science 2015, 350, 1214–1215. [Google Scholar] [CrossRef]
- Haran, J.P.; McCormick, B.A. Aging, frailty, and the microbiome—How dysbiosis influences human aging and disease. Gastroenterology 2021, 160, 507–523. [Google Scholar] [CrossRef]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef] [Green Version]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The Impact of the Gut Microbiota on Human Health: An Integrative View. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, D.; Jeffery, I.; Cusack, S.; O’Connor, E.; O’Toole, P. Diet-Microbiota-Health Interactions in Older Subjects: Implications for Healthy Aging. Aging Health-A Syst. Biol. Perspect. 2015, 40, 141–154. [Google Scholar] [CrossRef]
- Calderón Guzmán, D.; Juárez Olguín, H.; Osnaya Brizuela, N.; Hernández Garcia, E.; Lindoro Silva, M. The use of trace and essential elements in common clinical disorders: Roles in assessment of health and oxidative stress status. Nutr.Cancer 2019, 71, 13–20. [Google Scholar] [CrossRef]
- Vural, Z.; Avery, A.; Kalogiros, D.I.; Coneyworth, L.J.; Welham, S.J.M. Trace Mineral Intake and Deficiencies in Older Adults Living in the Community and Institutions: A Systematic Review. Nutrients 2020, 12, 1072. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Liang, Q.; Balakrishnan, B.; Belobrajdic, D.P.; Feng, Q.-J.; Zhang, W. Role of Dietary Nutrients in the Modulation of Gut Microbiota: A Narrative Review. Nutrients 2020, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Chaplin, A.; Parra, P.; Laraichi, S.; Serra, F.; Palou, A. Calcium supplementation modulates gut microbiota in a prebiotic manner in dietary obese mice. Mol. Nutr. Food Res. 2016, 60, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Cha, K.H.; Yang, J.-S.; Kim, K.-A.; Yoon, K.-Y.; Song, D.-G.; Erdene-Ochir, E.; Kang, K.; Pan, C.-H.; Ko, G. Improvement in host metabolic homeostasis and alteration in gut microbiota in mice on the high-fat diet: A comparison of calcium supplements. Food Res. Int. 2020, 136, 109495. [Google Scholar] [CrossRef] [PubMed]
- Kasaikina, M.V.; Kravtsova, M.A.; Lee, B.C.; Seravalli, J.; Peterson, D.A.; Walter, J.; Legge, R.; Benson, A.K.; Hatfield, D.L.; Gladyshev, V.N. Dietary selenium affects host selenoproteome expression by influencing the gut microbiota. Faseb J. 2011, 25, 2492–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, G.; Yu, M.; Li, K.; Hu, Y.; Wang, Y.; Xu, X.; Qu, J. Seleno-lentinan prevents chronic pancreatitis development and modulates gut microbiota in mice. J. Funct. Foods 2016, 22, 177–188. [Google Scholar] [CrossRef]
- Trautvetter, U.; Camarinha-Silva, A.; Jahreis, G.; Lorkowski, S.; Glei, M. High phosphorus intake and gut-related parameters—Results of a randomized placebo-controlled human intervention study. Nutr. J. 2018, 17, 23. [Google Scholar] [CrossRef]
- Yoon, L.; Michels, K. Characterizing the Effects of Calcium and Prebiotic Fiber on Human Gut Microbiota Composition and Function Using a Randomized Crossover Design—A Feasibility Study. Nutrients 2021, 13, 1937. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.J.; Wang, J.; Meng, X.L.; Sun, L.; Hu, B.; Li, H.B.; Sheng, J.; Chen, G.-M.; Tao, F.-B.; Sun, Y.-H.; et al. The association between essential trace element mixture and cognitive function in chinese community-dwelling older adults. Ecotoxicol. Environ. Saf. 2022, 231, 113182. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Eggers, S.; Safdar, N.; Sethi, A.K.; Suen, G.; Peppard, P.E.; Kates, A.E.; Skarlupka, J.H.; Kanarek, M.; Malecki, K.M. Urinary lead concentration and composition of the adult gut microbiota in a cross-sectional population-based sample. Environ. Int. 2019, 133, 105122. [Google Scholar] [CrossRef]
- Zeng, Y.; Li, J.; Wei, C.; Zhao, H.; Tao, W. mbDenoise: Microbiome data denoising using zero-inflated probabilistic principal components analysis. Genome Biol. 2022, 23, 94. [Google Scholar] [CrossRef]
- Bobb, J.F.; Valeri, L.; Henn, B.C.; Christiani, D.C.; Wright, R.O.; Mazumdar, M.; Godleski, J.J.; Coull, B.A. Bayesian kernel machine regression for estimating the health effects of multi-pollutant mixtures. Biostatistics 2015, 16, 493–508. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xiao, P.; Wang, R.; Luo, C.; Zhang, Z.; Yu, S.; Wu, Q.; Li, Y.; Zhang, Y.; Zhang, H.; et al. Relationships between urinary metals concentrations and cognitive performance among U.S. older people in NHANES 2011–2014. Front. Public Health 2022, 10, 985127. [Google Scholar] [CrossRef]
- Jiang, S.; Zhou, S.; Liu, H.; Peng, C.; Zhang, X.; Zhou, H.; Wang, Z.; Lu, Q. Concentrations of vanadium in urine with hypertension prevalence and blood pressure levels. Ecotoxicol. Environ. Saf. 2021, 213, 112028. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, T.; Zhang, Y.; Hu, Q.; Wang, X.; Chang, H.; Mao, J.-H.; Snijders, A.M.; Xia, Y. Contribution of trace element exposure to gestational diabetes mellitus through disturbing the gut microbiome. Environ. Int. 2021, 153, 106520. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.-J.; Yang, X.-L.; Zhang, H.; Xu, Q.; Wei, X.-T.; Feng, G.-J.; Zhao, M.; Pei, Y.-F.; Zhang, L. Assessing causal relationship from gut microbiota to heel bone mineral density. Bone 2021, 143, 115652. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, Z.; Celi, P.; Yan, L.; Ding, X.; Bai, S.; Zeng, Q.; Mao, X.; Feng, B.; Xu, S.; et al. Alteration of the Antioxidant Capacity and Gut Microbiota under High Levels of Molybdenum and Green Tea Polyphenols in Laying Hens. Antioxidants 2019, 8, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeem Aslam, M.; Bassis, C.M.; Zhang, L.; Zaidi, S.; Varani, J.; Bergin, I.L. Calcium reduces liver injury in mice on a high-fat diet: Alterations in microbial and bile acid profiles. PLoS ONE 2016, 11, e0166178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, J.; Costa, J.; Alfenas, R. Could the beneficial effects of dietary calcium on obesity and diabetes control be mediated by changes in intestinal microbiota and integrity? Br. J. Nutr. 2015, 114, 1756–1765. [Google Scholar] [CrossRef]
- Skrypnik, K.; Suliburska, J. Association between the gut microbiota and mineral metabolism. J. Sci. Food Agric. 2018, 98, 2449–2460. [Google Scholar] [CrossRef]
- Zeb, F.; Wu, X.; Chen, L.; Fatima, S.; Haq, I.-U.; Chen, A.; Xu, C.; Jianglei, R.; Feng, Q.; Li, M. Time-restricted feeding is associated with changes in human gut microbiota related to nutrient intake. Nutrition 2020, 78, 110797. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Cani, P.D. Diabetes, obesity and gut microbiota. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human Gut Microbes Associated with Obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Li, P.; Tang, T.; Chang, X.; Fan, X.; Chen, X.; Wang, R.; Fan, C.; Qi, K. Abnormality in Maternal Dietary Calcium Intake During Pregnancy and Lactation Promotes Body Weight Gain by Affecting the Gut Microbiota in Mouse Offspring. Mol. Nutr. Food Res. 2019, 63, e1800399. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basic Characteristics | Levels | n (%) |
|---|---|---|
| Categorical variable | ||
| Gender | Male | 140(51.9) |
| Female | 130(48.1) | |
| Education | Illiteracy | 128(47.4) |
| Primary school | 76(28.1) | |
| Junior school or above | 66(24.4) | |
| Family income | Low income | 200(74.1) |
| High income | 70(25.9) | |
| Region | Rural | 203(75.2) |
| Urban | 67(24.8) | |
| Smoking | Yes | 58(21.5) |
| No | 212(78.5) | |
| Drinking | Never | 192(71.1) |
| Often | 36(13.3) | |
| Always | 42(15.6) | |
| Physical activities | Low level | 95(35.2) |
| Moderate level | 78(28.9) | |
| Severe level | 97(35.9) | |
| Antibiotic used | Yes | 81(30.0) |
| No | 189(70.0) | |
| Diabetes | Yes | 92(34.1) |
| No | 178(65.9) | |
| CKD | Yes | 37(13.7) |
| No | 233(86.3) | |
| Hypertension | Yes | 171(63.3) |
| No | 99(36.7) | |
| Continuous variables | Mean ± SD/Range(median) | |
| Age (years) | - | 71.422 ± 4.911 |
| BMI (kg/m2) | - | 24.675 ± 3.739 |
| Dietary pattern1 | - | −2.703–10.792(−0.262) |
| Dietary pattern2 | - | −1.382–9.073(−0.255) |
| Dietary pattern3 | - | −2.005–4.694(−0.227) |
| Dietary pattern4 | - | −1.517–6.367(−0.282) |
| Dietary pattern5 | - | −8.173–2.648(0.043) |
| Elements | LOD | DR% | GM | Unadjusted (Creatinine Adjusted) a | ||||
|---|---|---|---|---|---|---|---|---|
| 5th | 25th | 50th | 75th | 95th | ||||
| V | 0.005 | 100% | 1.804 (2.225) | 0.746 (0.532) | 1.238 (1.088) | 1.730 (1.773) | 2.258 (2.720) | 3.150 (5.673) |
| Co | 0.001 | 100% | 0.369 (0.405) | 0.010 (0.117) | 0.182 (0.201) | 0.289 (0.300) | 0.443 (0.462) | 0.941 (1.238) |
| Se | 0.274 | 100% | 16.795 (17.374) | 3.892 (4.678) | 8.855 (10.657) | 13.867 (15.179) | 20.518 (20.641) | 37.616 (38.424) |
| Sr | 0.015 | 100% | 215.848 (233.08) | 51.942 (56.235) | 100.366 (112.627) | 178.797 (192.590) | 282.117 (331.390) | 540.303 (557.702) |
| Mg | 0.0022 | 100% | 121.837 (124.291) | 21.142 (34.060) | 60.933 (69.872) | 103.073 (105.487) | 149.911 (150.942) | 293.133 (321.088) |
| Ca | 0.0127 | 100% | 149.939 (159.150) | 23.140 (30.732) | 59.810 (63.997) | 113.075 (128.613) | 202.794 (225.834) | 391.535 (405.582) |
| Mo | 0.002 | 100% | 116.160 (115.540) | 18.992 (24.983) | 44.287 (56.391) | 87.116 (89.089) | 159.657 (138.747) | 340.495 (315.389) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Wang, Y.; Chen, G.; Wang, H.; Sun, L.; Zhang, D.; Tao, F.; Zhang, Z.; Yang, L. The Associations between Multiple Essential Metal(loid)s and Gut Microbiota in Chinese Community-Dwelling Older Adults. Nutrients 2023, 15, 1137. https://doi.org/10.3390/nu15051137
Zhang J, Wang Y, Chen G, Wang H, Sun L, Zhang D, Tao F, Zhang Z, Yang L. The Associations between Multiple Essential Metal(loid)s and Gut Microbiota in Chinese Community-Dwelling Older Adults. Nutrients. 2023; 15(5):1137. https://doi.org/10.3390/nu15051137
Chicago/Turabian StyleZhang, Jianghui, Yuan Wang, Guimei Chen, Hongli Wang, Liang Sun, Dongmei Zhang, Fangbiao Tao, Zhihua Zhang, and Linsheng Yang. 2023. "The Associations between Multiple Essential Metal(loid)s and Gut Microbiota in Chinese Community-Dwelling Older Adults" Nutrients 15, no. 5: 1137. https://doi.org/10.3390/nu15051137
APA StyleZhang, J., Wang, Y., Chen, G., Wang, H., Sun, L., Zhang, D., Tao, F., Zhang, Z., & Yang, L. (2023). The Associations between Multiple Essential Metal(loid)s and Gut Microbiota in Chinese Community-Dwelling Older Adults. Nutrients, 15(5), 1137. https://doi.org/10.3390/nu15051137