
Elevated Maternal Folate Status and Changes in Maternal Prolactin, Placental Lactogen and Placental Growth Hormone Following Folic Acid Food Fortification: Evidence from Two Prospective Pregnancy Cohorts

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
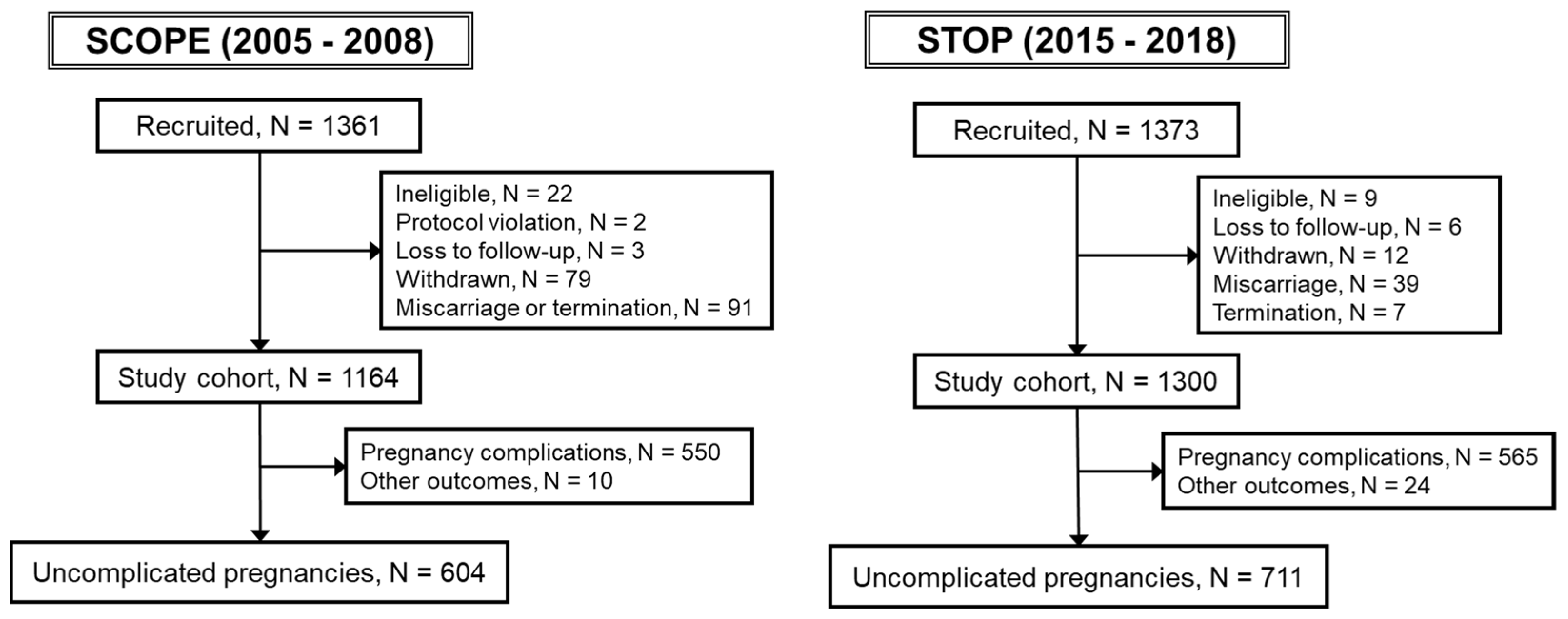
2.1. Study Population
2.2. Data Collection
2.3. Biochemical Measurements
2.4. Statistics
2.5. Ethics Approval and Patient Consent
3. Results
3.1. Descriptive Characteristics
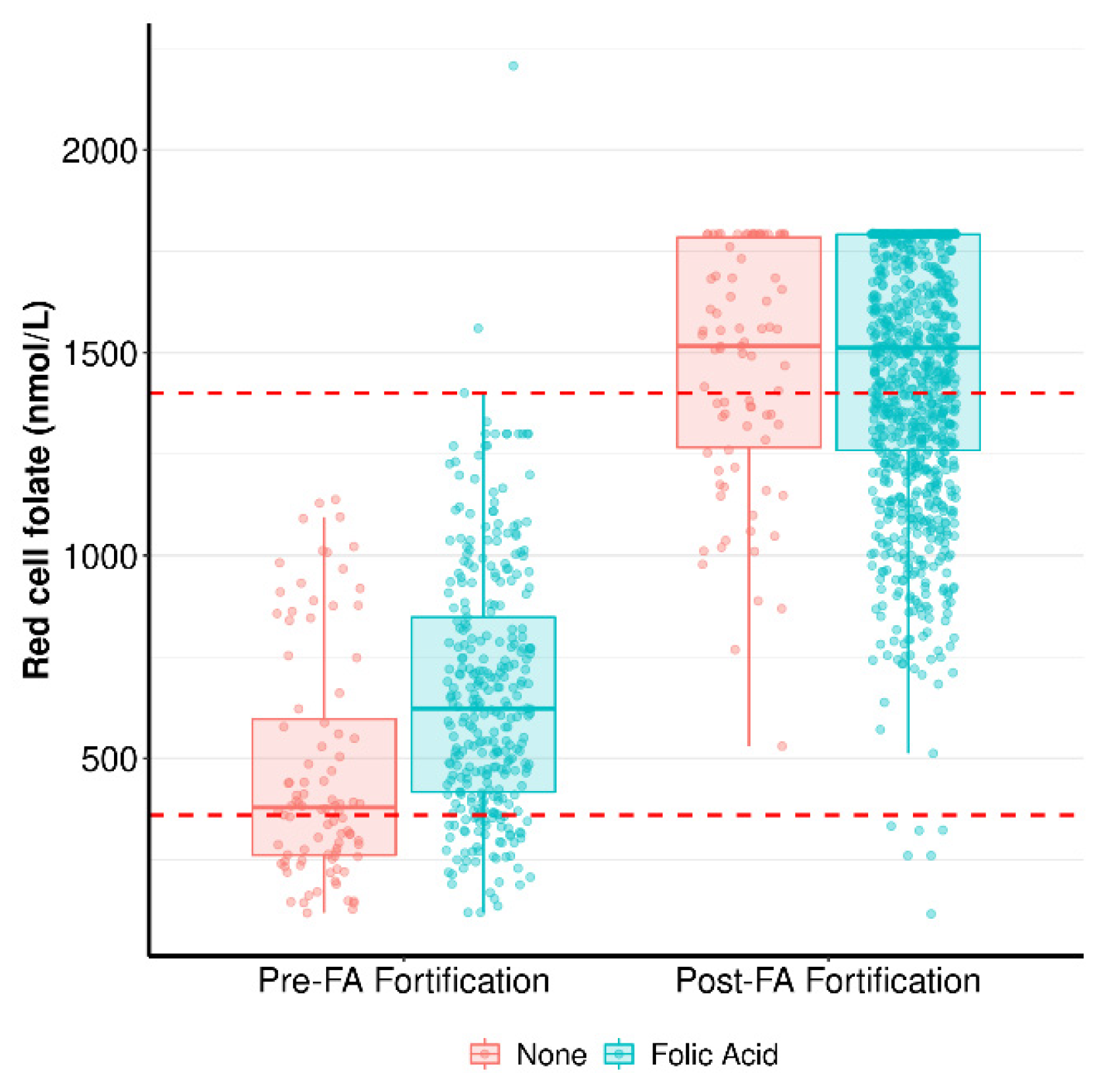
3.2. One-Carbon Metabolism Components in Early Pregnancy in Response to FA Supplementation
3.3. Pregnancy Hormones
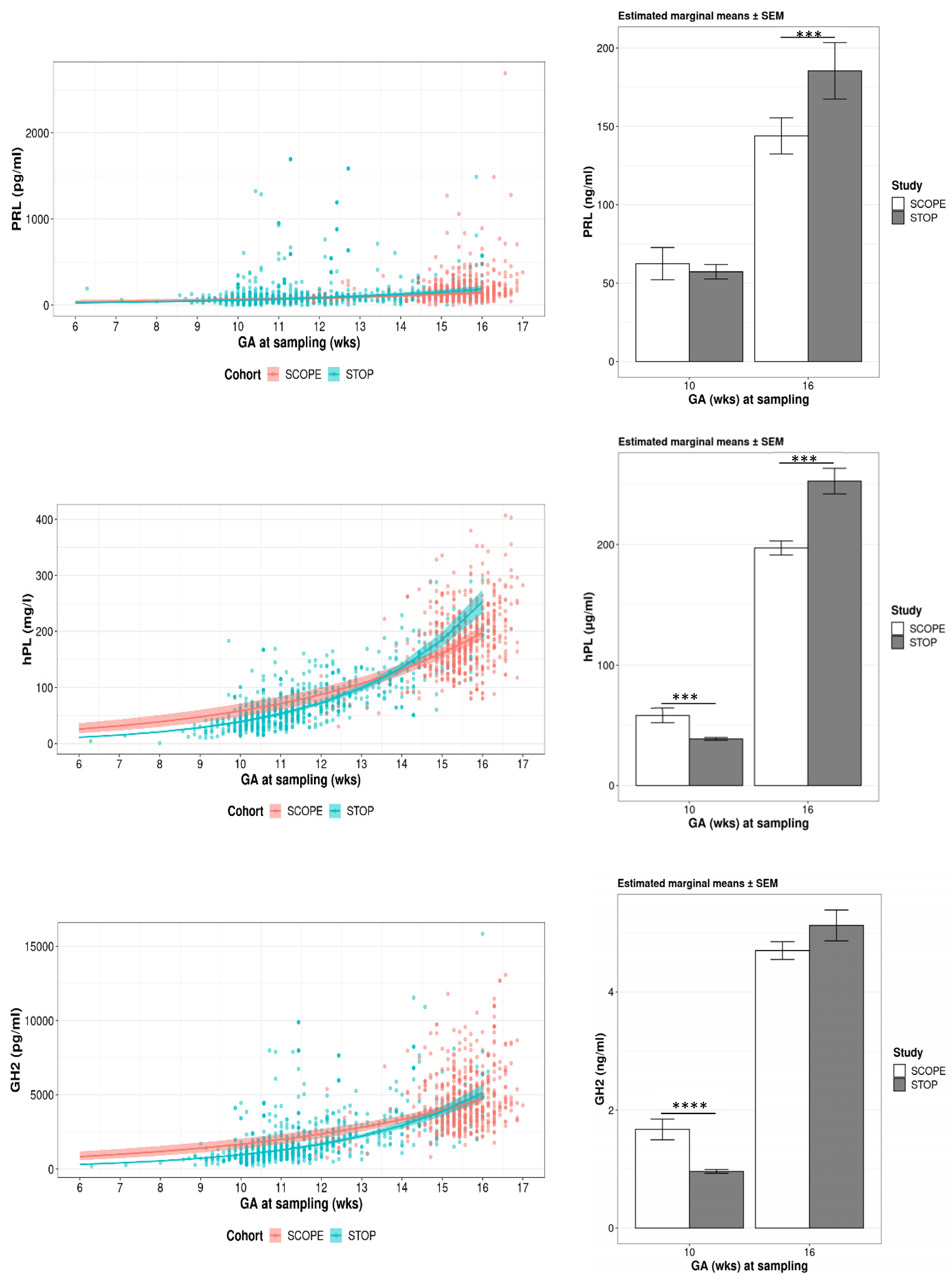
3.3.1. Gestational Modelling
3.3.2. Estimated Means for PRL, hPL and GH2
3.3.3. PRL, hPL and GH2 Concentrations Relative to FA Supplementation
3.3.4. PRL, hPL and GH2 Concentrations Relative to Blood 1C Metabolism Biochemistry
3.3.5. PRL, hPL and GH2 Concentrations Relative to Maternal Age and/or BMI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Molloy, A.M.; Kirke, P.N.; Brody, L.C.; Scott, J.M.; Mills, J.L. Effects of folate and vitamin B12 deficiencies during pregnancy on fetal, infant, and child development. Food Nutr. Bull. 2008, 29 (Suppl. S2), S101–S111; discussion S112–S105. [Google Scholar] [CrossRef] [PubMed]
- Field, M.S.; Stover, P.J. Safety of folic acid. Ann. N. Y. Acad. Sci. 2018, 1414, 59–71. [Google Scholar] [CrossRef] [Green Version]
- MRC-Vitamin-Study-Research-Group. Prevention of neural tube defects: Results of the Medical Research Council Vitamin Study. MRC Vitamin Study Research Group. Lancet 1991, 338, 131–137. [Google Scholar] [CrossRef]
- Czeizel, A.E.; Dudas, I. Prevention of the first occurrence of neural-tube defects by periconceptional vitamin supplementation. N. Engl. J. Med. 1992, 327, 1832–1835. [Google Scholar] [CrossRef]
- Murphy, M.E.; Westmark, C.J. Folic Acid Fortification and Neural Tube Defect Risk: Analysis of the Food Fortification Initiative Dataset. Nutrients 2020, 12, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food-Standards-Australia-and-New-Zealand. Folic Acid/Folate and Pregnancy. Available online: https://www.foodstandards.gov.au/consumer/nutrition/folicmandatory/Pages/default.aspx (accessed on 12 January 2023).
- Ledowsky, C.; Mahimbo, A.; Scarf, V.; Steel, A. Women Taking a Folic Acid Supplement in Countries with Mandatory Food Fortification Programs May Be Exceeding the Upper Tolerable Limit of Folic Acid: A Systematic Review. Nutrients 2022, 14, 2715. [Google Scholar] [CrossRef] [PubMed]
- Livock, M.; Anderson, P.J.; Lewis, S.; Bowden, S.; Muggli, E.; Halliday, J. Maternal micronutrient consumption periconceptionally and during pregnancy: A prospective cohort study. Public. Health Nutr. 2017, 20, 294–304. [Google Scholar] [CrossRef] [Green Version]
- Pietrzik, K.; Bailey, L.; Shane, B. Folic acid and L-5-methyltetrahydrofolate: Comparison of clinical pharmacokinetics and pharmacodynamics. Clin. Pharmacokinet. 2010, 49, 535–548. [Google Scholar] [CrossRef]
- Jhaveri, M.S.; Wagner, C.; Trepel, J.B. Impact of extracellular folate levels on global gene expression. Mol. Pharmacol. 2001, 60, 1288–1295. [Google Scholar] [CrossRef] [Green Version]
- Pufulete, M.; Al-Ghnaniem, R.; Khushal, A.; Appleby, P.; Harris, N.; Gout, S.; Emery, P.W.; Sanders, T.A. Effect of folic acid supplementation on genomic DNA methylation in patients with colorectal adenoma. Gut 2005, 54, 648–653. [Google Scholar] [CrossRef] [Green Version]
- Kok, D.E.; Dhonukshe-Rutten, R.A.; Lute, C.; Heil, S.G.; Uitterlinden, A.G.; van der Velde, N.; van Meurs, J.B.; van Schoor, N.M.; Hooiveld, G.J.; de Groot, L.C.; et al. The effects of long-term daily folic acid and vitamin B12 supplementation on genome-wide DNA methylation in elderly subjects. Clin. Epigenetics 2015, 7, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Froese, D.S.; Fowler, B.; Baumgartner, M.R. Vitamin B12, folate, and the methionine remethylation cycle-biochemistry, pathways, and regulation. J. Inherit. Metab. Dis. 2019, 42, 673–685. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, S.; Hasui, S.; Yamamoto, C.; Yoshioka, C.; Kobayashi, M.; Itagaki, S.; Hirano, T.; Iseki, K. Placental folate transport during pregnancy. Biosci. Biotechnol. Biochem. 2008, 72, 2277–2284. [Google Scholar] [CrossRef] [PubMed]
- Solanky, N.; Requena Jimenez, A.; D’Souza, S.W.; Sibley, C.P.; Glazier, J.D. Expression of folate transporters in human placenta and implications for homocysteine metabolism. Placenta 2010, 31, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.J.; Bulmer, J.N.; Innes, B.A.; Broughton Pipkin, F. Possible roles for folic acid in the regulation of trophoblast invasion and placental development in normal early human pregnancy. Biol. Reprod. 2011, 84, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Irwin, R.E.; Thursby, S.J.; Ondicova, M.; Pentieva, K.; McNulty, H.; Richmond, R.C.; Caffrey, A.; Lees-Murdock, D.J.; McLaughlin, M.; Cassidy, T.; et al. A randomized controlled trial of folic acid intervention in pregnancy highlights a putative methylation-regulated control element at ZFP57. Clin. Epigenetics 2019, 11, 31. [Google Scholar] [CrossRef]
- Ondicova, M.; Irwin, R.E.; Thursby, S.J.; Hilman, L.; Caffrey, A.; Cassidy, T.; McLaughlin, M.; Lees-Murdock, D.J.; Ward, M.; Murphy, M.; et al. Folic acid intervention during pregnancy alters DNA methylation, affecting neural target genes through two distinct mechanisms. Clin. Epigenetics 2022, 14, 63. [Google Scholar] [CrossRef]
- Tserga, A.; Binder, A.M.; Michels, K.B. Impact of folic acid intake during pregnancy on genomic imprinting of IGF2/H19 and 1-carbon metabolism. FASEB J. 2017, 31, 5149–5158. [Google Scholar] [CrossRef] [Green Version]
- Rahat, B.; Hamid, A.; Bagga, R.; Kaur, J. Folic Acid Levels During Pregnancy Regulate Trophoblast Invasive Behavior and the Possible Development of Preeclampsia. Front. Nutr. 2022, 9, 847136. [Google Scholar] [CrossRef]
- Ly, L.; Chan, D.; Landry, M.; Angle, C.; Martel, J.; Trasler, J. Impact of mothers’ early life exposure to low or high folate on progeny outcome and DNA methylation patterns. Environ. Epigenet 2020, 6, dvaa018. [Google Scholar] [CrossRef]
- Rosario, F.J.; Nathanielsz, P.W.; Powell, T.L.; Jansson, T. Maternal folate deficiency causes inhibition of mTOR signaling, down-regulation of placental amino acid transporters and fetal growth restriction in mice. Sci. Rep. 2017, 7, 3982. [Google Scholar] [CrossRef]
- Furness, D.L.; Yasin, N.; Dekker, G.A.; Thompson, S.D.; Roberts, C.T. Maternal red blood cell folate concentration at 10–12 weeks gestation and pregnancy outcome. J. Matern.-Fetal Neonatal Med. 2012, 25, 1423–1427. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Himes, K.P.; Venkataramanan, R.; Chen, J.Y.; Evans, R.W.; Meyer, J.L.; Simhan, H.N. Maternal serum folate species in early pregnancy and risk of preterm birth. Am. J. Clin. Nutr. 2010, 92, 864–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgetts, V.A.; Morris, R.K.; Francis, A.; Gardosi, J.; Ismail, K.M. Effectiveness of folic acid supplementation in pregnancy on reducing the risk of small-for-gestational age neonates: A population study, systematic review and meta-analysis. BJOG 2015, 122, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lv, L.; Zhang, H.; Zhao, N.; Qiu, J.; He, X.; Zhou, M.; Xu, X.; Cui, H.; Liu, S.; et al. Folic acid supplementation, dietary folate intake and risk of preterm birth in China. Eur. J. Nutr. 2016, 55, 1411–1422. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.; Peng, X.; Zhang, S.; Wang, X.; Zhu, C. Folic Acid and Risk of Preterm Birth: A Meta-Analysis. Front. Neurosci. 2019, 13, 1284. [Google Scholar] [CrossRef] [Green Version]
- Bailey, S.W.; Ayling, J.E. The extremely slow and variable activity of dihydrofolate reductase in human liver and its implications for high folic acid intake. Proc. Natl. Acad. Sci. USA 2009, 106, 15424–15429. [Google Scholar] [CrossRef] [Green Version]
- Patanwala, I.; King, M.J.; Barrett, D.A.; Rose, J.; Jackson, R.; Hudson, M.; Philo, M.; Dainty, J.R.; Wright, A.J.; Finglas, P.M.; et al. Folic acid handling by the human gut: Implications for food fortification and supplementation. Am. J. Clin. Nutr. 2014, 100, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Wang, B.; Sahyoun, N.R. Application of the Key Events Dose-response Framework to Folate Metabolism. Crit. Rev. Food Sci. Nutr. 2016, 56, 1325–1333. [Google Scholar] [CrossRef]
- Sweeney, M.R.; McPartlin, J.; Scott, J. Folic acid fortification and public health: Report on threshold doses above which unmetabolised folic acid appear in serum. BMC Public Health 2007, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Obeid, R.; Kirsch, S.H.; Dilmann, S.; Klein, C.; Eckert, R.; Geisel, J.; Herrmann, W. Folic acid causes higher prevalence of detectable unmetabolized folic acid in serum than B-complex: A randomized trial. Eur. J. Nutr. 2016, 55, 1021–1028. [Google Scholar] [CrossRef]
- Dai, C.; Fei, Y.; Li, J.; Shi, Y.; Yang, X. A Novel Review of Homocysteine and Pregnancy Complications. Biomed. Res. Int. 2021, 2021, 6652231. [Google Scholar] [CrossRef] [PubMed]
- Williamson, J.M.; Arthurs, A.L.; Smith, M.D.; Roberts, C.T.; Jankovic-Karasoulos, T. High Folate, Perturbed One-Carbon Metabolism and Gestational Diabetes Mellitus. Nutrients 2022, 14, 3930. [Google Scholar] [CrossRef]
- Krishnaveni, G.V.; Veena, S.R.; Karat, S.C.; Yajnik, C.S.; Fall, C.H. Association between maternal folate concentrations during pregnancy and insulin resistance in Indian children. Diabetologia 2014, 57, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.S.; Pang, W.W.; Cai, S.; Lee, Y.S.; Chan, J.K.Y.; Shek, L.P.C.; Yap, F.K.P.; Tan, K.H.; Godfrey, K.M.; van Dam, R.M.; et al. High folate and low vitamin B12 status during pregnancy is associated with gestational diabetes mellitus. Clin. Nutr. 2018, 37, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, P.; Sukumar, N.; Adaikalakoteswari, A.; Goljan, I.; Venkataraman, H.; Gopinath, A.; Bagias, C.; Yajnik, C.S.; Stallard, N.; Ghebremichael-Weldeselassie, Y. Association of maternal vitamin B12 and folate levels in early pregnancy with gestational diabetes: A prospective UK cohort study (PRiDE study). Diabetologia 2021, 64, 2170–2182. [Google Scholar] [CrossRef]
- Deng, M.; Zhou, J.; Tang, Z.; Xiang, J.; Yi, J.; Peng, Y.; Di, L.; Zhai, X.; Yang, M.; Du, Y. The correlation between plasma total homocysteine level and gestational diabetes mellitus in a Chinese Han population. Sci. Rep. 2020, 10, 18679. [Google Scholar] [CrossRef]
- Zheng, Y.; Deng, H.Y.; Qiao, Z.Y.; Gong, F.X. Homocysteine level and gestational diabetes mellitus: A systematic review and meta-analysis. Gynecol. Endocrinol. 2021, 37, 987–994. [Google Scholar] [CrossRef] [PubMed]
- AIHW. Australia’s Health. 2020. Available online: https://www.aihw.gov.au/reports/diabetes/diabetes/contents/how-many-australians-have-diabetes/gestational-diabetes (accessed on 12 January 2023).
- Laurie, J.G.; McIntyre, H.D. A Review of the Current Status of Gestational Diabetes Mellitus in Australia-The Clinical Impact of Changing Population Demographics and Diagnostic Criteria on Prevalence. Int. J. Environ. Res. Public. Health 2020, 17, 9387. [Google Scholar] [CrossRef]
- Huang, Y.; He, Y.; Sun, X.; He, Y.; Li, Y.; Sun, C. Maternal high folic acid supplement promotes glucose intolerance and insulin resistance in male mouse offspring fed a high-fat diet. Int. J. Mol. Sci. 2014, 15, 6298–6313. [Google Scholar] [CrossRef]
- Keating, E.; Correia-Branco, A.; Araujo, J.R.; Meireles, M.; Fernandes, R.; Guardao, L.; Guimaraes, J.T.; Martel, F.; Calhau, C. Excess perigestational folic acid exposure induces metabolic dysfunction in post-natal life. J. Endocrinol. 2015, 224, 245–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koseki, K.; Maekawa, Y.; Bito, T.; Yabuta, Y.; Watanabe, F. High-dose folic acid supplementation results in significant accumulation of unmetabolized homocysteine, leading to severe oxidative stress in Caenorhabditis elegans. Redox Biol. 2020, 37, 101724. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, Y.; Liu, Z.; Tian, Z.; Pei, X.; Liu, L.; Li, Y. Folic acid oversupplementation during pregnancy disorders lipid metabolism in male offspring via regulating arginase 1-associated NOS3-AMPKalpha pathway. Clin. Nutr. 2022, 41, 21–32. [Google Scholar] [CrossRef]
- Pannia, E.; Yang, N.V.; Ho, M.; Chatterjee, D.; Hammoud, R.; Kubant, R.; Anderson, G.H. Folic acid content of diet during pregnancy determines post-birth re-set of metabolism in Wistar rat dams. J. Nutr. Biochem. 2020, 83, 108414. [Google Scholar] [CrossRef] [PubMed]
- Stern, C.; Schwarz, S.; Moser, G.; Cvitic, S.; Jantscher-Krenn, E.; Gauster, M.; Hiden, U. Placental Endocrine Activity: Adaptation and Disruption of Maternal Glucose Metabolism in Pregnancy and the Influence of Fetal Sex. Int. J. Mol. Sci. 2021, 22, 12722. [Google Scholar] [CrossRef] [PubMed]
- Kampmann, U.; Knorr, S.; Fuglsang, J.; Ovesen, P. Determinants of Maternal Insulin Resistance during Pregnancy: An Updated Overview. J. Diabetes Res. 2019, 2019, 5320156. [Google Scholar] [CrossRef] [Green Version]
- Napso, T.; Yong, H.E.J.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. The Role of Placental Hormones in Mediating Maternal Adaptations to Support Pregnancy and Lactation. Front. Physiol. 2018, 9, 1091. [Google Scholar] [CrossRef] [Green Version]
- Barbour, L.A.; Shao, J.; Qiao, L.; Leitner, W.; Anderson, M.; Friedman, J.E.; Draznin, B. Human placental growth hormone increases expression of the p85 regulatory unit of phosphatidylinositol 3-kinase and triggers severe insulin resistance in skeletal muscle. Endocrinology 2004, 145, 1144–1150. [Google Scholar] [CrossRef] [Green Version]
- Barbour, L.A.; Shao, J.; Qiao, L.; Pulawa, L.K.; Jensen, D.R.; Bartke, A.; Garrity, M.; Draznin, B.; Friedman, J.E. Human placental growth hormone causes severe insulin resistance in transgenic mice. Am. J. Obstet. Gynecol. 2002, 186, 512–517. [Google Scholar] [CrossRef]
- Sonagra, A.D.; Biradar, S.M.; K, D.; Murthy, D.S.J. Normal pregnancy—A state of insulin resistance. J. Clin. Diagn. Res. 2014, 8, CC01–CC03. [Google Scholar] [CrossRef]
- Simpson, S.; Smith, L.; Bowe, J. Placental peptides regulating islet adaptation to pregnancy: Clinical potential in gestational diabetes mellitus. Curr. Opin. Pharmacol. 2018, 43, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, R.R.; Cyphert, H.A.; Walker, E.M.; Chakravarthy, H.; Peiris, H.; Gu, X.; Liu, Y.; Conrad, E.; Goodrich, L.; Stein, R.W.; et al. Gestational Diabetes Mellitus From Inactivation of Prolactin Receptor and MafB in Islet beta-Cells. Diabetes 2016, 65, 2331–2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brelje, T.C.; Scharp, D.W.; Lacy, P.E.; Ogren, L.; Talamantes, F.; Robertson, M.; Friesen, H.G.; Sorenson, R.L. Effect of homologous placental lactogens, prolactins, and growth hormones on islet B-cell division and insulin secretion in rat, mouse, and human islets: Implication for placental lactogen regulation of islet function during pregnancy. Endocrinology 1993, 132, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.A.; Enns, L. Role of gestational hormones in the induction of insulin resistance. J. Clin. Endocrinol. Metab. 1988, 67, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Richard, K.; Holland, O.; Landers, K.; Vanderlelie, J.J.; Hofstee, P.; Cuffe, J.S.M.; Perkins, A.V. Review: Effects of maternal micronutrient supplementation on placental function. Placenta 2017, 54, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, B.C.; Hayes, D.J.; Jones, R.L. Effects of micronutrients on placental function: Evidence from clinical studies to animal models. Reproduction 2018, 156, R69–R82. [Google Scholar] [CrossRef] [Green Version]
- Rasool, A.; Alvarado-Flores, F.; O’Tierney-Ginn, P. Placental Impact of Dietary Supplements: More Than Micronutrients. Clin. Ther. 2021, 43, 226–245. [Google Scholar] [CrossRef]
- Meher, A.; Sundrani, D.; Joshi, S. Maternal nutrition influences angiogenesis in the placenta through peroxisome proliferator activated receptors: A novel hypothesis. Mol. Reprod. Dev. 2015, 82, 726–734. [Google Scholar] [CrossRef]
- Davis, P.; McLeod, K.; Ransom, M.; Ongley, P.; Pearce, N.; Howden-Chapman, P. The New Zealand Socioeconomic Index: Developing and validating an occupationally-derived indicator of socio-economic status. Aust. N. Z. J. Public. Health 1999, 23, 27–33. [Google Scholar] [CrossRef]
- van Gool, J.D.; Hirche, H.; Lax, H.; De Schaepdrijver, L. Folic acid and primary prevention of neural tube defects: A review. Reprod. Toxicol. 2018, 80, 73–84. [Google Scholar] [CrossRef]
- Pinunuri, R.; Castano, E.; Ronco, A. Folates during Pregnancy: Food Fortification versus Supplementation with Folic Acid. Acad. J. Pediatr. Neonatol. 2017, 5, 555705. [Google Scholar]
- Kalhan, S.C.; Marczewski, S.E. Methionine, homocysteine, one carbon metabolism and fetal growth. Rev. Endocr. Metab. Disord. 2012, 13, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Azadibakhsh, N.; Hosseini, R.S.; Atabak, S.; Nateghiyan, N.; Golestan, B.; Rad, A.H. Efficacy of folate and vitamin B12 in lowering homocysteine concentrations in hemodialysis patients. Saudi J. Kidney Dis. Transpl. 2009, 20, 779–788. [Google Scholar] [PubMed]
- Study of the Effectiveness of Additional Reductions in Cholesterol and Homocysteine (SEARCH) Collaborative Group; Armitage, J.M.; Bowman, L.; Clarke, R.J.; Wallendszus, K.; Bulbulia, R.; Rahimi, K.; Haynes, R.; Parish, S.; Sleight, P.; et al. Effects of homocysteine-lowering with folic acid plus vitamin B12 vs placebo on mortality and major morbidity in myocardial infarction survivors: A randomized trial. JAMA 2010, 303, 2486–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.J.; Huang, M.C.; Chung, L.J.; Cheng, C.H.; Lin, K.L.; Su, K.H.; Huang, Y.C. Folic acid and vitamin B12 are more effective than vitamin B6 in lowering fasting plasma homocysteine concentration in patients with coronary artery disease. Eur. J. Clin. Nutr. 2004, 58, 481–487. [Google Scholar] [CrossRef]
- Smith, A.D.; Warren, M.J.; Refsum, H. Vitamin B(12). Adv. Food Nutr. Res. 2018, 83, 215–279. [Google Scholar] [CrossRef]
- Ortbauer, M.; Ripper, D.; Fuhrmann, T.; Lassi, M.; Auernigg-Haselmaier, S.; Stiegler, C.; Konig, J. Folate deficiency and over-supplementation causes impaired folate metabolism: Regulation and adaptation mechanisms in Caenorhabditis elegans. Mol. Nutr. Food Res. 2016, 60, 949–956. [Google Scholar] [CrossRef]
- Maher, A.; Sobczynska-Malefora, A. The Relationship Between Folate, Vitamin B12 and Gestational Diabetes Mellitus With Proposed Mechanisms and Foetal Implications. J. Family Reprod. Health 2021, 15, 141–149. [Google Scholar] [CrossRef]
- Zhang, X.; Qu, Y.Y.; Liu, L.; Qiao, Y.N.; Geng, H.R.; Lin, Y.; Xu, W.; Cao, J.; Zhao, J.Y. Homocysteine inhibits pro-insulin receptor cleavage and causes insulin resistance via protein cysteine-homocysteinylation. Cell Rep. 2021, 37, 109821. [Google Scholar] [CrossRef]
- Salbaum, J.M.; Kappen, C. Genetic and epigenomic footprints of folate. Prog. Mol. Biol. Transl. Sci. 2012, 108, 129–158. [Google Scholar] [CrossRef] [Green Version]
- Haas, C.B.; Su, Y.R.; Petersen, P.; Wang, X.; Bien, S.A.; Lin, Y.; Albanes, D.; Weinstein, S.J.; Jenkins, M.A.; Figueiredo, J.C.; et al. Interactions between folate intake and genetic predictors of gene expression levels associated with colorectal cancer risk. Sci. Rep. 2022, 12, 18852. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.; Li, L.; Jiang, Y.; Tan, J.; Ji, J.; Zhang, Y.; Jin, N.; Liu, F. Excess Folic Acid Supplementation Before and During Pregnancy and Lactation Activates Fos Gene Expression and Alters Behaviors in Male Mouse Offspring. Front. Neurosci. 2019, 13, 313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, H.; Hossain, M.B.; Lera, L.; Hirsch, S.; Albala, C.; Uauy, R.; Broberg, K.; Ronco, A.M. High levels of circulating folate concentrations are associated with DNA methylation of tumor suppressor and repair genes p16, MLH1, and MGMT in elderly Chileans. Clin. Epigenetics 2017, 9, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penailillo, R.; Guajardo, A.; Llanos, M.; Hirsch, S.; Ronco, A.M. Folic acid supplementation during pregnancy induces sex-specific changes in methylation and expression of placental 11beta-hydroxysteroid dehydrogenase 2 in rats. PLoS ONE 2015, 10, e0121098. [Google Scholar] [CrossRef] [Green Version]
- Alnabbat, K.I.; Fardous, A.M.; Cabelof, D.C.; Heydari, A.R. Excessive Folic Acid Mimics Folate Deficiency in Human Lymphocytes. Curr. Issues Mol. Biol. 2022, 44, 1452–1462. [Google Scholar] [CrossRef]
- Lombardo, M.F.; De Angelis, F.; Bova, L.; Bartolini, B.; Bertuzzi, F.; Nano, R.; Capuani, B.; Lauro, R.; Federici, M.; Lauro, D.; et al. Human placental lactogen (hPL-A) activates signaling pathways linked to cell survival and improves insulin secretion in human pancreatic islets. Islets 2011, 3, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Baeyens, L.; Hindi, S.; Sorenson, R.L.; German, M.S. beta-Cell adaptation in pregnancy. Diabetes Obes. Metab. 2016, 18 (Suppl. S1), 63–70. [Google Scholar] [CrossRef] [Green Version]
- Handwerger, S.; Freemark, M. Role of placental lactogen and prolactin in human pregnancy. Adv. Exp. Med. Biol. 1987, 219, 399–420. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y.; Chen, H.; Jiang, Y.; Wang, Y.; Wang, D.; Li, M.; Dou, Y.; Sun, X.; Huang, G.; et al. Association of Maternal Folate and Vitamin B12 in Early Pregnancy With Gestational Diabetes Mellitus: A Prospective Cohort Study. Diabetes Care 2021, 44, 217–223. [Google Scholar] [CrossRef]
- Gong, T.; Wang, J.; Yang, M.; Shao, Y.; Liu, J.; Wu, Q.; Xu, Q.; Wang, H.; He, X.; Chen, Y.; et al. Serum homocysteine level and gestational diabetes mellitus: A meta-analysis. J. Diabetes Investig. 2016, 7, 622–628. [Google Scholar] [CrossRef] [Green Version]
- Waterland, R.A.; Jirtle, R.L. Transposable elements: Targets for early nutritional effects on epigenetic gene regulation. Mol. Cell. Biol. 2003, 23, 5293–5300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornet, D.; Clement, A.; Clement, P.; Menezo, Y. High doses of folic acid induce a pseudo-methylenetetrahydrofolate syndrome. SAGE Open. Med. Case Rep. 2019, 7, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.; Leclerc, D.; Cosin-Tomas, M.; Malysheva, O.V.; Wasek, B.; Bottiglieri, T.; Caudill, M.A.; Rozen, R. Moderate Folic Acid Supplementation in Pregnant Mice Results in Altered Methyl Metabolism and in Sex-Specific Placental Transcription Changes. Mol. Nutr. Food Res. 2021, 65, e2100197. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Jiang, J.; Guo, L. Effects of maternal folate and vitamin B12 on gestational diabetes mellitus: A dose-response meta-analysis of observational studies. Eur. J. Clin. Nutr. 2022, 76, 1502–1512. [Google Scholar] [CrossRef]
- Huang, L.; Yu, X.; Li, L.; Chen, Y.; Yang, Y.; Yang, Y.; Hu, Y.; Zhao, Y.; Tang, H.; Xu, D.; et al. Duration of periconceptional folic acid supplementation and risk of gestational diabetes mellitus. Asia Pac. J. Clin. Nutr. 2019, 28, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Sha, T.; Gao, X.; He, Q.; Wu, X.; Tian, Q.; Yang, F.; Tang, C.; Wu, X.; Xie, Q.; et al. The Associations between the Duration of Folic Acid Supplementation, Gestational Diabetes Mellitus, and Adverse Birth Outcomes based on a Birth Cohort. Int. J. Environ. Res. Public. Health 2019, 16, 4511. [Google Scholar] [CrossRef] [Green Version]
- Lean, S.C.; Heazell, A.E.P.; Dilworth, M.R.; Mills, T.A.; Jones, R.L. Placental Dysfunction Underlies Increased Risk of Fetal Growth Restriction and Stillbirth in Advanced Maternal Age Women. Sci. Rep. 2017, 7, 9677. [Google Scholar] [CrossRef]
- Haavaldsen, C.; Fedorcsak, P.; Tanbo, T.; Eskild, A. Maternal age and serum concentration of human chorionic gonadotropin in early pregnancy. Acta Obstet. Gynecol. Scand. 2014, 93, 1290–1294. [Google Scholar] [CrossRef]
- Cattini, P.A.; Jin, Y.; Jarmasz, J.S.; Noorjahan, N.; Bock, M.E. Obesity and regulation of human placental lactogen production in pregnancy. J. Neuroendocrinol. 2020, 32, e12859. [Google Scholar] [CrossRef]
- Lassance, L.; Haghiac, M.; Minium, J.; Catalano, P.; Hauguel-de Mouzon, S. Obesity-induced down-regulation of the mitochondrial translocator protein (TSPO) impairs placental steroid production. J. Clin. Endocrinol. Metab. 2015, 100, E11–E18. [Google Scholar] [CrossRef] [Green Version]
- Tuzcu, A.; Bahceci, M.; Dursun, M.; Turgut, C.; Bahceci, S. Insulin sensitivity and hyperprolactinemia. J. Endocrinol. Investig. 2003, 26, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Lin, J.; Li, H.; Liu, Z.; Chen, X.; Chen, Q. Prolactin Is Associated With Insulin Resistance and Beta-Cell Dysfunction in Infertile Women With Polycystic Ovary Syndrome. Front. Endocrinol. 2021, 12, 571229. [Google Scholar] [CrossRef] [PubMed]
- Atmaca, A.; Bilgici, B.; Ecemis, G.C.; Tuncel, O.K. Evaluation of body weight, insulin resistance, leptin and adiponectin levels in premenopausal women with hyperprolactinemia. Endocrine 2013, 44, 756–761. [Google Scholar] [CrossRef]
- Ekinci, E.I.; Torkamani, N.; Ramchand, S.K.; Churilov, L.; Sikaris, K.A.; Lu, Z.X.; Houlihan, C.A. Higher maternal serum prolactin levels are associated with reduced glucose tolerance during pregnancy. J. Diabetes Investig. 2017, 8, 697–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngala, R.A.; Fondjo, L.A.; Gmagna, P.; Ghartey, F.N.; Awe, M.A. Placental peptides metabolism and maternal factors as predictors of risk of gestational diabetes in pregnant women. A case-control study. PLoS ONE 2017, 12, e0181613. [Google Scholar] [CrossRef] [Green Version]
- Lolis, D.; Tzingounis, V.; Kaskarelis, D. Maternal serums and amniotic fluid levels of human placental lactogen in gestational diabetes. Eur. J. Clin. Investig. 1978, 8, 259–260. [Google Scholar] [CrossRef]
- Ursell, W.; Brudenell, M.; Chard, T. Placental lactogen levels in diabetic pregnancy. Br. Med. J. 1973, 2, 80–82. [Google Scholar] [CrossRef] [Green Version]
- Soler, N.G.; Nicholson, H.O.; Malins, J.M. Serial determinations of human placental lactogen in the management of diabetic pregnancy. Lancet 1975, 2, 54–57. [Google Scholar] [CrossRef]
- Hu, L.; Lytras, A.; Bock, M.E.; Yuen, C.K.; Dodd, J.G.; Cattini, P.A. Detection of placental growth hormone variant and chorionic somatomammotropin-L RNA expression in normal and diabetic pregnancy by reverse transcriptase-polymerase chain reaction. Mol. Cell. Endocrinol. 1999, 157, 131–142. [Google Scholar] [CrossRef]
- Kirwan, J.P.; Hauguel-De Mouzon, S.; Lepercq, J.; Challier, J.C.; Huston-Presley, L.; Friedman, J.E.; Kalhan, S.C.; Catalano, P.M. TNF-alpha is a predictor of insulin resistance in human pregnancy. Diabetes 2002, 51, 2207–2213. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Hou, Y.; Yan, X.; Wang, Y.; Shi, C.; Wu, X.; Liu, H.; Zhang, L.; Zhang, X.; Liu, J. Joint effects of folate and vitamin B12 imbalance with maternal characteristics on gestational diabetes mellitus. Journal. Diabetes 2019, 11, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Du, Y.; Xia, S.; Li, Z.; Liu, J. Vitamin B(12) and gestational diabetes mellitus: A systematic review and meta-analysis. Br. J. Nutr. 2022, 129, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Adaikalakoteswari, A.; Vatish, M.; Alam, M.T.; Ott, S.; Kumar, S.; Saravanan, P. Low Vitamin B12 in Pregnancy Is Associated With Adipose-Derived Circulating miRs Targeting PPARgamma and Insulin Resistance. J. Clin. Endocrinol. Metab. 2017, 102, 4200–4209. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SCOPE (N = 604) | STOP (N = 711) | p-Value | |
|---|---|---|---|
| Maternal age (y): Median (IQ range) | 23.0 (20.0–26.0) | 25.0 (22.0–29.0) | <0.0001 |
| Maternal BMI: Median (IQ range) | 25.1 (22.0–29.1) | 25.3 (22.4–29.5) | 0.27 |
| * SEI: Median (IQ range) | 24.0 (20.0–30.0) | 29.0 (22.0–45.0) | <0.0001 |
| Ethnicity: White N (%) | 0.0005 | ||
| Yes | 555 (91.9) | 595 (83.7) | |
| No | 49 (8.1) | 116 (16.3) | |
| Smoking: N (%) | 0.002 | ||
| Missing | 0 (0.0) | 6 (0.8) | |
| No | 476 (78.8) | 602 (84.7) | |
| Yes | 128 (21.2) | 103 (14.5) | |
| Folic acid supplementation (μg/day): N (%) | <0.0001 | ||
| Missing | 0 (0.0) | 87 (12.2) | |
| None | 122 (20.2) | 49 (6.9) | |
| ≤400 | 66 (10.9) | 7 (1.0) | |
| >400 to <800 | 269 (44.5) | 127 (17.9) | |
| ≥800 | 147 (24.3) | 441 (62.0) | |
| B12 supplementation (μg/day): N (%) | <0.0001 | ||
| None | 284 (47) | 202 (28.4) | |
| Yes | 320 (53) | 509 (71.6) | |
| Blood biochemistry (** 1C metabolism) | |||
| Serum folate (nmol/L): Median (IQ range) | 36.4 (28.5–43.0) | 40.5 (36.4–45.2) | <0.0001 |
| B12 (pmol/L): Median (IQ range) | 248.0 (195.5–316.0) | 277.5 (219.0–355.0) | <0.0001 |
| Homocysteine (μmol/L): Mean (SD) | 4.6 (1.1) | 5.0 (1.1) | <0.0001 |
| Induction of labour: N (%) | <0.0001 | ||
| No | 470 (77.8) | 415 (58.4) | |
| Yes | 132 (22.2) | 296 (41.6) | |
| Birth Outcomes | |||
| Baby Sex: N (%) | 0.55 | ||
| Missing | 0 (0.0) | 1 (0.1) | |
| Female | 313 (51.8) | 355 (49.9) | |
| Male | 291 (48.2) | 355 (49.9) | |
| Gestational age at birth (weeks): Median (IQ range) | 40.3 (39.4–41.0) | 40.0 (39.1–40.9) | 0.01 |
| Birthweight (g): Median (IQ range) | 3543 (3290–3825) | 3506 (3250–3786) | 0.03 |
| Birthweight customised centile: Mean (SD) | 54.2 (25.1) | 53.3 (26.1) | 0.52 |
| Folate (nmol/L) Median (IQ Range) | B12 (pmol/L) Median (IQ Range) | Folate:B12 Ratio Median (IQ Range) | Homocysteine (μmol/L) Mean (SD) | ||
|---|---|---|---|---|---|
| SCOPE | None (N = 122) | 27.1 (18.5–34.3) | 217.5 (189.0–280.5) | 117.7 (80.2–162.5) | 5.1 (1.4) |
| ≤400 (N = 66) | 35.6 (28.9–41.6) | 251 (192.5–338.0) | 131.6 (107.4–184.5) | 4.5 (1.1) | |
| >400 to <800 (N = 269) | 38.4 (30.4–44.2) | 252 (198.0–318.0) | 146.9 (107.8–228.4) | 4.5 (1.1) | |
| ≥800 (N = 147) | 39.3 (34.8–44.2) | 260 (208.0–315.5) | 156.5 (115.7–238.9) | 4.3 (0.9) | |
| STOP | None (N = 49) | 38.9 (34.3–43.2) | 277.5 (219.2–316.5) | 137.7 (109.8–211.8) | 5.2 (1.4) |
| ≤400 (N = 7) | 42.1 (36.6–56.3) | 261 (203.5–315.5) | 180.2 (165.0–201.9) | 5.1 (1.1) | |
| >400 to <800 (N = 127) | 39.8 (35.4–44.5) | 260 (205.0–346.5) | 168 (116.3–248.2) | 5.3 (1.3) | |
| ≥800 (N = 441) | 41.3 (37.3–55.2) | 281 (231.0–370.0) | 158.1 (118.6–222.1) | 4.8 (1.0) |
| Study | FA Supplementation | PRL Estimated Marginal Mean (ng/mL): (SE) | hPL Estimated Marginal Mean (μg/mL): (SE) | GH2 Estimated Marginal Mean (ng/mL): (SE) |
|---|---|---|---|---|
| SCOPE | None (N = 122) | 71.12 (6.89) | 90.59 (3.87) | 2.36 (0.12) |
| ≤400 (N = 66) | 88.11 (9.58) | 94.1 (4.89) | 2.57 (0.15) | |
| >400 to <800 (N = 269) | 96.52 (8.81) | 92.3 (3.4) | 2.43 (0.10) | |
| ≥800 (N = 147) | 109.84 (10.75) | 92.4 (3.84) | 2.53 (0.13) | |
| STOP | None (N = 49) | 124.42 (15.42) | 110.09 (6.22) | 2.59 (0.19) |
| ≤400 (N = 7) | 175.23 (45.83) | 127.63 (16.72) | 1.90 (0.34) | |
| >400 to <800 (N = 127) | 151.39 (14.13) | 107.24 (4.19) | 2.29 (0.10) | |
| ≥800 (N = 441) | 90.05 (7.63) | 104.86 (3.33) | 2.21 (0.08) |
| B12 Ratio of Geometric Means (95% CI) | p-Value | Folate:B12 Ratio of Geometric Means (95% CI) | p-Value | Hcy Ratio of Geometric Means (95% CI) | p-Value | |
|---|---|---|---|---|---|---|
| Prolactin | 1.05 (1.02, 1.09) | 0.0005 | 1.01 (0.99, 1.03) | 0.4 | 0.93 (0.90, 0.96) | <0.0001 |
| hPL | 1.02 (1.00, 1.04) | 0.03 | 0.99 (0.98, 1.01) | 0.3 | 0.98 (0.97, 1.00) | 0.06 |
| GH2 | 1.03 (1.01, 1.05) | 0.008 | 0.98 (0.96, 0.99) | 0.004 | 0.98 (0.96, 1.00) | 0.1 |
| Study | Contrast | PRL | p-Value | hPL | p-Value | GH2 | p-Value |
|---|---|---|---|---|---|---|---|
| SCOPE | ≥35y/<35y | 1.11 (0.83, 1.50) | 0.5 | 0.92 (0.77, 1.10) | 0.4 | 0.93 (0.75, 1.14) | 0.5 |
| STOP | ≥35y/<35y | 0.69 (0.55, 0.87) | 0.002 | 0.92 (0.80, 1.05) | 0.2 | 0.93 (0.79, 1.09) | 0.4 |
| Study | Contrast | PRL | p-Value | hPL | p-Value | GH2 | p-Value |
|---|---|---|---|---|---|---|---|
| SCOPE | Obese/Non-obese | 0.81 (0.67, 0.98) | 0.02 | 0.79 (0.72, 0.88) | <0.0001 | 0.73 (0.64, 0.83) | <0.0001 |
| STOP | Obese/Non-obese | 0.70 (0.59, 0.82) | <0.0001 | 0.77 (0.71, 0.84) | <0.0001 | 0.92 (0.82, 1.03) | 0.2 |
| Subgroup | Contrast | Ratio of Geometric Means (95% CI) | p-Value |
|---|---|---|---|
| SCOPE | <35y obese/<35y non-obese | 0.74 (0.62, 0.89) | <0.0001 |
| ≥35y non-obese/<35y non-obese | 0.78 (0.45, 1.37) | 0.7 | |
| ≥35y obese/<35y non-obese | 1.17 (0.68, 2.01) | 0.9 | |
| ≥35y non-obese/<35y obese | 1.05 (0.59, 1.88) | 1 | |
| ≥35y obese/<35y obese | 1.58 (0.92, 2.72) | 0.1 | |
| ≥35y obese/≥35y non-obese | 1.50 (0.69, 3.25) | 0.5 | |
| STOP | <35y obese/<35y non-obese | 0.76 (0.65, 0.89) | <0.0001 |
| ≥35y non-obese/<35y non-obese | 0.67 (0.47, 0.95) | 0.02 | |
| ≥35y obese/<35y non-obese | 0.52 (0.29, 0.94) | 0.02 | |
| ≥35y non-obese/<35y obese | 0.88 (0.61, 1.27) | 0.8 | |
| ≥35y obese/<35y obese | 0.68 (0.37, 1.24) | 0.4 | |
| ≥35y obese/≥35y non-obese | 0.77 (0.39, 1.53) | 0.8 | |
| STOP/SCOPE | <35y and non-obese | 1.17 (1.03, 1.33) | 0.01 |
| <35y and obese | 1.20 (0.99, 1.46) | 0.06 | |
| ≥35y and non-obese | 1.00 (0.60, 1.67) | 1 | |
| ≥35y and obese | 0.52 (0.28, 0.96) | 0.04 |
| Subgroup | Contrast | Ratio of Geometric Means (95% CI) | p-Value |
|---|---|---|---|
| SCOPE | <35y obese/<35y non-obese | 0.82 (0.75, 0.90) | <0.0001 |
| ≥35y non-obese/<35y non-obese | 1.04 (0.76, 1.42) | 1 | |
| ≥35y obese/<35y non-obese | 0.69 (0.50, 0.96) | 0.02 | |
| ≥35y non-obese/<35y obese | 1.26 (0.92, 1.74) | 0.2 | |
| ≥35y obese/<35y obese | 0.84 (0.61, 1.17) | 0.5 | |
| ≥35y obese/≥35y non-obese | 0.67 (0.43, 1.04) | 0.09 | |
| STOP | <35y obese/<35y non-obese | 0.79 (0.73, 0.86) | <0.0001 |
| ≥35y non-obese/<35y non-obese | 0.92 (0.76, 1.13) | 0.7 | |
| ≥35y obese/<35y non-obese | 0.70 (0.52, 0.96) | 0.02 | |
| ≥35y non-obese/<35y obese | 1.17 (0.95, 1.44) | 0.2 | |
| ≥35y obese/<35y obese | 0.89 (0.65, 1.22) | 0.8 | |
| ≥35y obese/≥35y non-obese | 0.76 (0.53, 1.10) | 0.2 | |
| STOP/SCOPE | <35y and non-obese | 1.12 (1.04, 1.20) | 0.002 |
| <35y and obese | 1.08 (0.97, 1.19) | 0.1 | |
| ≥35y and non-obese | 1.00 (0.75, 1.33) | 1 | |
| ≥35y and obese | 1.14 (0.81, 1.60) | 0.5 |
| Subgroup | Contrast | Ratio of Geometric Means (95% CI) | p-Value |
|---|---|---|---|
| SCOPE | <35y obese/<35y non-obese | 0.77 (0.68, 0.87) | <0.0001 |
| ≥35y non-obese/<35y non-obese | 1.03 (0.70, 1.51) | 1 | |
| ≥35y obese/<35y non-obese | 0.69 (0.47, 1.01) | 0.06 | |
| ≥35y non-obese/<35y obese | 1.34 (0.90, 2.00) | 0.2 | |
| ≥35y obese/<35y obese | 0.90 (0.61, 1.33) | 0.9 | |
| ≥35y obese/≥35y non-obese | 0.68 (0.39, 1.16) | 0.2 | |
| STOP | <35y obese/<35y non-obese | 0.88 (0.79, 0.98) | 0.02 |
| ≥35y non-obese/<35y non-obese | 0.87 (0.68, 1.10) | 0.4 | |
| ≥35y obese/<35y non-obese | 0.91 (0.59, 1.41) | 0.9 | |
| ≥35y non-obese/<35y obese | 0.99 (0.76, 1.28) | 1 | |
| ≥35y obese/<35y obese | 1.03 (0.66, 1.61) | 1 | |
| ≥35y obese/≥35y non-obese | 1.05 (0.64, 1.71) | 1 | |
| STOP/SCOPE | <35y and non-obese | 0.90 (0.82, 0.98) | 0.01 |
| <35y and obese | 1.03 (0.90, 1.17) | 0.7 | |
| ≥35y and non-obese | 0.76 (0.53, 1.08) | 0.1 | |
| ≥35y and obese | 1.17 (0.76, 1.83) | 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankovic-Karasoulos, T.; Smith, M.D.; Leemaqz, S.; Williamson, J.; McCullough, D.; Arthurs, A.L.; Jones, L.A.; Bogias, K.J.; Mol, B.W.; Dalton, J.; et al. Elevated Maternal Folate Status and Changes in Maternal Prolactin, Placental Lactogen and Placental Growth Hormone Following Folic Acid Food Fortification: Evidence from Two Prospective Pregnancy Cohorts. Nutrients 2023, 15, 1553. https://doi.org/10.3390/nu15071553
Jankovic-Karasoulos T, Smith MD, Leemaqz S, Williamson J, McCullough D, Arthurs AL, Jones LA, Bogias KJ, Mol BW, Dalton J, et al. Elevated Maternal Folate Status and Changes in Maternal Prolactin, Placental Lactogen and Placental Growth Hormone Following Folic Acid Food Fortification: Evidence from Two Prospective Pregnancy Cohorts. Nutrients. 2023; 15(7):1553. https://doi.org/10.3390/nu15071553
Chicago/Turabian StyleJankovic-Karasoulos, Tanja, Melanie D. Smith, Shalem Leemaqz, Jessica Williamson, Dylan McCullough, Anya L. Arthurs, Lauren A. Jones, Konstantinos Justin Bogias, Ben W. Mol, Julia Dalton, and et al. 2023. "Elevated Maternal Folate Status and Changes in Maternal Prolactin, Placental Lactogen and Placental Growth Hormone Following Folic Acid Food Fortification: Evidence from Two Prospective Pregnancy Cohorts" Nutrients 15, no. 7: 1553. https://doi.org/10.3390/nu15071553
APA StyleJankovic-Karasoulos, T., Smith, M. D., Leemaqz, S., Williamson, J., McCullough, D., Arthurs, A. L., Jones, L. A., Bogias, K. J., Mol, B. W., Dalton, J., Dekker, G. A., & Roberts, C. T. (2023). Elevated Maternal Folate Status and Changes in Maternal Prolactin, Placental Lactogen and Placental Growth Hormone Following Folic Acid Food Fortification: Evidence from Two Prospective Pregnancy Cohorts. Nutrients, 15(7), 1553. https://doi.org/10.3390/nu15071553