
Nobiletin Improves D-Galactose-Induced Aging Mice Skeletal Muscle Atrophy by Regulating Protein Homeostasis

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs and Reagents
2.2. Animals and Treatments
2.3. Body Composition
2.4. Muscle Fatigue Test
2.5. Grip Strength Test
2.6. Histological Staining
2.7. Transmission Electron Micrographs (TEMs)
2.8. Extraction of Myofibrillar Protein
2.9. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.10. RNA Extraction and Real-Time qPCR (RT-qPCR)
2.11. Protein Extraction and Western Blotting
2.12. Determination of Inflammatory Cytokines
2.13. Statistical Analysis
3. Results
3.1. Effect of Nobiletin (Nob) on Skeletal Muscle Mass and Function in D-Galactose-Induced (D-Gal-Induced) Aging Mice
3.2. Effect of Nob on the Number and Structure of Myofiber in D-Gal-Induced Aging Mice
3.3. Effect of Nob on Skeletal Muscle Composition in D-Gal-Induced Aging Mice
3.4. Effect of Nob on Skeletal Muscle Protein Synthesis in D-Gal-Induced Aging Mice
3.5. Effect of Nob on Skeletal Muscle Protein Degradation in D-Gal-Induced Aging Mice
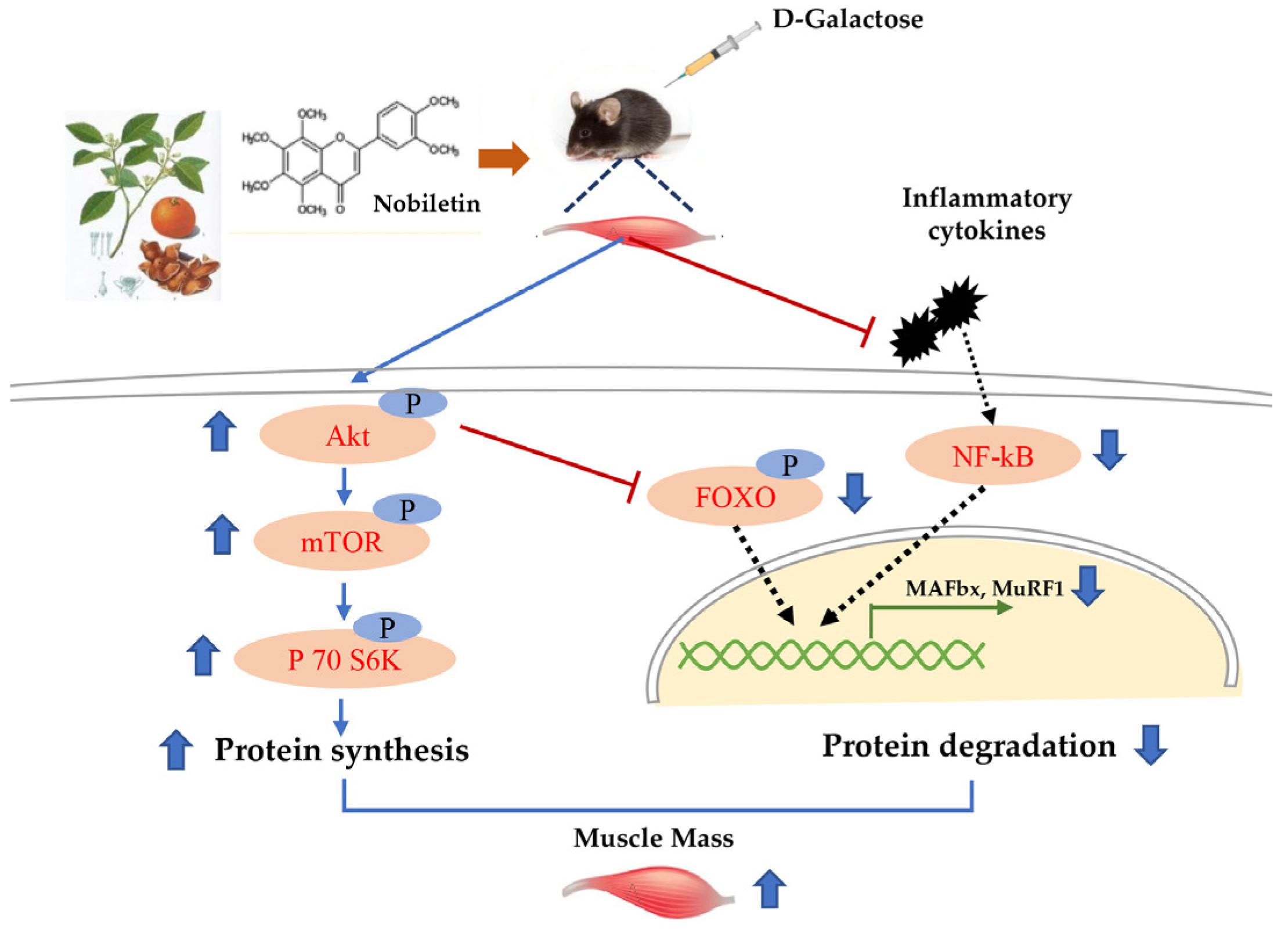
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rafi, S.T.M.; Sambandam, Y.; Sittadjody, S.; Pathak, S.; Ramachandran, I.; Kumaran, R.I. Chapter 9-Skeletal Muscle Cell Aging and Stem Cells. In Stem Cells and Aging; Pathak, S., Banerjee, A., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 125–145. [Google Scholar] [CrossRef]
- Chang, Y.C.; Liu, H.W.; Chen, Y.T.; Chen, Y.A.; Chen, Y.J.; Chang, S.J. Resveratrol protects muscle cells against palmitate-induced cellular senescence and insulin resistance through ameliorating autophagic flux. J. Food Drug Anal. 2018, 26, 1066–1074. [Google Scholar] [CrossRef]
- Palla, A.R.; Ravichandran, M.; Wang, Y.X.; Alexandrova, L.; Yang, A.V.; Kraft, P.; Holbrook, C.A.; Schurch, C.M.; Ho, A.T.V.; Blau, H.M. Inhibition of prostaglandin-degrading enzyme 15-PGDH rejuvenates aged muscle mass and strength. Science 2021, 371, eabc8059. [Google Scholar] [CrossRef]
- Tournadre, A.; Vial, G.; Capel, F.; Soubrier, M.; Boirie, Y. Sarcopenia. Jt. Bone Spine 2019, 86, 309–314. [Google Scholar]
- Tan, K.T.; Ang, S.J.; Tsai, S.Y. Sarcopenia: Tilting the Balance of Protein Homeostasis. Proteomics 2020, 20, e1800411. [Google Scholar] [CrossRef]
- Kou, X.; Li, J.; Liu, X.; Yang, X.; Fan, J.; Chen, N. Ampelopsin attenuates the atrophy of skeletal muscle from d-gal-induced aging rats through activating AMPK/SIRT1/PGC-1α signaling cascade. Biomed. Pharmacother. 2017, 90, 311–320. [Google Scholar]
- Yamauchi, Y.; Ferdousi, F.; Fukumitsu, S.; Isoda, H. Maslinic Acid Attenuates Denervation-Induced Loss of Skeletal Muscle Mass and Strength. Nutrients 2021, 13, 2950. [Google Scholar] [CrossRef]
- Janssen, I.; Shepard, D.S.; Katzmarzyk, P.T.; Roubenoff, R. The healthcare costs of sarcopenia in the United States. J. Am. Geriatr. Soc. 2004, 52, 80–85. [Google Scholar] [CrossRef]
- Ganapathy, A.; Nieves, J.W. Nutrition and Sarcopenia—What Do We Know? Nutrients 2020, 12, 1755. [Google Scholar] [CrossRef]
- Phillips, S.M.; Lau, K.J.; D’Souza, A.C.; Nunes, E.A. An umbrella review of systematic reviews of beta-hydroxy-beta-methyl butyrate supplementation in ageing and clinical practice. J. Cachexia. Sarcopenia. Muscle. 2022, 13, 2265–2275. [Google Scholar] [CrossRef]
- Bunbupha, S.; Pakdeechote, P.; Maneesai, P.; Prasarttong, P. Nobiletin alleviates high-fat diet-induced nonalcoholic fatty liver disease by modulating AdipoR1 and gp91(phox) expression in rats. J. Nutr. Biochem. 2021, 87, 108526. [Google Scholar] [CrossRef]
- Nohara, K.; Mallampalli, V.; Nemkov, T.; Wirianto, M.; Yang, J.; Ye, Y.Q.; Sun, Y.X.; Han, L.; Esser, K.A.; Mileykovskaya, E.; et al. Nobiletin fortifies mitochondrial respiration in skeletal muscle to promote healthy aging against metabolic challenge. Nat. Commun. 2019, 10, 3923. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Cha, B.Y.; Choi, S.S.; Choi, B.K.; Yonezawa, T.; Teruya, T.; Nagai, K.; Woo, J.T. Nobiletin improves obesity and insulin resistance in high-fat diet-induced obese mice. J. Nutr. Biochem. 2013, 24, 156–162. [Google Scholar] [CrossRef]
- Wang, H.; Guo, Y.; Qiao, Y.; Zhang, J.; Jiang, P. Nobiletin Ameliorates NLRP3 Inflammasome-Mediated Inflammation Through Promoting Autophagy via the AMPK Pathway. Mol. Neurobiol. 2020, 57, 5056–5068. [Google Scholar] [CrossRef]
- Rolland, Y.; Onder, G.; Morley, J.E.; Gillette-Guyonet, S.; van Kan, G.A.; Vellas, B. Current and future pharmacologic treatment of sarcopenia. Clin. Geriatr. Med. 2011, 27, 423–447. [Google Scholar] [CrossRef]
- Bonaldo, P.; Sandri, M. Cellular and molecular mechanisms of muscle atrophy. Dis. Model. Mech. 2013, 6, 25–39. [Google Scholar] [CrossRef] [Green Version]
- de Alvaro, C.; Teruel, T.; Hernandez, R.; Lorenzo, M. Tumor necrosis factor alpha produces insulin resistance in skeletal muscle by activation of inhibitor kappaB kinase in a p38 MAPK-dependent manner. J. Biol. Chem. 2004, 279, 17070–17078. [Google Scholar] [CrossRef] [Green Version]
- Dogra, C.; Changotra, H.; Wedhas, N.; Qin, X.; Wergedal, J.E.; Kumar, A. TNF-related weak inducer of apoptosis (TWEAK) is a potent skeletal muscle-wasting cytokine. Faseb. J. 2007, 21, 1857–1869. [Google Scholar] [CrossRef] [Green Version]
- Hirosumi, J.; Tuncman, G.; Chang, L.F.; Gorgun, C.Z.; Uysal, K.T.; Maeda, K.; Karin, M.; Hotamisligil, G.S. A central role for JNK in obesity and insulin resistance. Nature 2002, 420, 333–336. [Google Scholar] [CrossRef]
- Feng, W.; Liu, J.; Wang, S.; Hu, Y.; Pan, H.; Hu, T.; Guan, H.; Zhang, D.; Mao, Y. Alginate oligosaccharide alleviates D-galactose-induced cardiac ageing via regulating myocardial mitochondria function and integrity in mice. .J. Cell Mol. Med. 2021, 25, 7157–7168. [Google Scholar] [CrossRef]
- Sun, Y.X.; Wang, P.; Zheng, H.; Smith, R.G. Ghrelin stimulation of growth hormone release and appetite is mediated through the growth hormone secretagogue receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 4679–4684. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.H.; Song, Y.Y.; Liu, Z.Q.; Li, M.H.; Zhang, L.; Yu, Q.L.; Guo, Z.B.; Wei, J.M. Effects of iron-catalyzed and metmyoglobin oxidizing systems on biochemical properties of yak muscle myofibrillar protein. Meat Sci. 2020, 166, 108041. [Google Scholar] [CrossRef]
- Romanello, V.; Sandri, M. Mitochondrial Quality Control and Muscle Mass Maintenance. Front. Physiol. 2016, 6, eCollection 2015. [Google Scholar] [CrossRef]
- Sandri, M. Protein breakdown in muscle wasting: Role of autophagy-lysosome and ubiquitin-proteasome. Int. J. Biochem. Cell. B. 2013, 45, 2121–2129. [Google Scholar] [CrossRef] [Green Version]
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-Related Loss of Muscle Mass and Function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef]
- Denison, H.J.; Cooper, C.; Sayer, A.A.; Robinson, S.M. Prevention and optimal management of sarcopenia: A review of combined exercise and nutrition interventions to improve muscle outcomes in older people. Clin. Interv. Aging 2015, 10, 859–869. [Google Scholar] [CrossRef] [Green Version]
- Kwak, J.Y.; Kwon, K.S. Pharmacological Interventions for Treatment of Sarcopenia: Current Status of Drug Development for Sarcopenia. Ann. Geriatr. Med. Res. 2019, 23, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Azman, K.F.; Zakaria, R. D-Galactose-induced accelerated aging model: An overview. Biogerontology 2019, 20, 763–782. [Google Scholar] [CrossRef]
- Wang, H.H.; Sun, Y.N.; Qu, T.Q.; Sang, X.Q.; Zhou, L.M.; Li, Y.X.; Ren, F.Z. Nobiletin Prevents D-Galactose-Induced C2C12 Cell Aging by Improving Mitochondrial Function. Int. J. Mol. Sci. 2022, 23, 11963. [Google Scholar] [CrossRef]
- Wu, Y.X.; Wu, Y.X.; Yang, Y.F.; Yu, J.; Wu, J.H.; Liao, Z.Y.; Guo, A.; Sun, Y.; Zhao, Y.X.; Chen, J.L.; et al. Lysyl oxidase-like 2 inhibitor rescues D-galactose-induced skeletal muscle fibrosis. Aging Cell 2022, 21, e13659. [Google Scholar] [CrossRef]
- Chen, Q.N.; Fan, Z.; Lyu, A.K.; Wu, J.; Guo, A.; Yang, Y.F.; Chen, J.L.; Xiao, Q. Effect of sarcolipin-mediated cell transdifferentiation in sarcopenia-associated skeletal muscle fibrosis. Exp. Cell. Res. 2020, 389, 111890. [Google Scholar] [CrossRef]
- Gao, Q.; Chen, F.; Zhang, L.J.; Wei, A.; Wang, Y.X.; Wu, Z.W.; Cao, W.S. Inhibition of DNA methyltransferase aberrations reinstates antioxidant aging suppressors and ameliorates renal aging. Aging Cell 2022, 21, e13526. [Google Scholar] [CrossRef]
- He, B.; Nohara, K.; Park, N.; Park, Y.S.; Guillory, B.; Zhao, Z.; Garcia, J.M.; Koike, N.; Lee, C.C.; Takahashi, J.S.; et al. The Small Molecule Nobiletin Targets the Molecular Oscillator to Enhance Circadian Rhythms and Protect against Metabolic Syndrome. Cell Metab. 2016, 23, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Altun, M.; Besche, H.C.; Overkleeft, H.S.; Piccirillo, R.; Edelmann, M.J.; Kessler, B.M.; Goldberg, A.L.; Ulfhake, B. Muscle Wasting in Aged, Sarcopenic Rats Is Associated with Enhanced Activity of the Ubiquitin Proteasome Pathway. J. Biol. Chem. 2010, 285, 39597–39608. [Google Scholar] [CrossRef] [Green Version]
- Clavel, S.; Coldefy, A.S.; Kurkdjian, E.; Salles, J.; Margaritis, I.; Derijard, B. Atrophy-related ubiquitin ligases, atrogin-1 and MuRF1 are up-regulated in aged rat Tibialis Anterior muscle. Mech. Ageing Dev. 2006, 127, 794–801. [Google Scholar] [CrossRef]
- Welle, S.; Brooks, A.I.; Delehanty, J.M.; Needler, N.; Thornton, C.A. Gene expression profile of aging in human muscle. Physiol. Genom. 2003, 14, 149–159. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence 5′ → 3′ (Forward) | Sequence 5′ → 3′ (Reverse) |
|---|---|---|
| ACTA1 | TACCACCGGCATCGTGTTG | GCGCACAATCTCACGTTCAG |
| TMP1 | TTGAAAGCCGAGCCCAAAAAG | TCATACTTCCGGTCAGCATCTT |
| TNNC1 | GCGGTAGAACAGTTGACAGAG | GACAGAAACTCATCGAAGTCCA |
| TNNC2 | GAGGCCAGGTCCTACCTCAG | GGTGCCCAACTCTTTAACGCT |
| TNNT1 | ACTAAAAGACCGCATTGGAGG | AGCTCCCATGTTGGACAGAAC |
| TNNT3 | ACTGCTCCTAAGATCCCGGAA | ATGAGGTCCTTGTTTTGACGC |
| TNNI1 | ATGCCGGAAGTTGAGAGGAAA | TCCGAGAGGTAACGCACCTT |
| TNNI2 | CGGAGGGTGCGTATGTCTG | CAGGTCCCGTTCCTTCTCA |
| MYH1 | CTCTTCCCGCTTTGGTAAGTT | CAGGACATTTCGATTAGATCCG |
| MYH2 | TGGAGGGTGAGGTAGAGAGTG | TTGGATAGATTTGTGTTGGATTG |
| MYH4 | CCGCATCTGTAGGAAGGGG | GTGACCGAATTTGTACTGATGT |
| MYH7 | CCTGCGGAAGTCTGAGAAGG | CTCGGGACACGATCTTGGC |
| GAPDH | TGGCCTTCCGTGTTCCTAC | GAGGCTGTGAAGTCGCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-H.; Zhang, Y.; Qu, T.-Q.; Sang, X.-Q.; Li, Y.-X.; Ren, F.-Z.; Wen, P.-C.; Sun, Y.-N. Nobiletin Improves D-Galactose-Induced Aging Mice Skeletal Muscle Atrophy by Regulating Protein Homeostasis. Nutrients 2023, 15, 1801. https://doi.org/10.3390/nu15081801
Wang H-H, Zhang Y, Qu T-Q, Sang X-Q, Li Y-X, Ren F-Z, Wen P-C, Sun Y-N. Nobiletin Improves D-Galactose-Induced Aging Mice Skeletal Muscle Atrophy by Regulating Protein Homeostasis. Nutrients. 2023; 15(8):1801. https://doi.org/10.3390/nu15081801
Chicago/Turabian StyleWang, Hui-Hui, Yan Zhang, Tai-Qi Qu, Xue-Qin Sang, Yi-Xuan Li, Fa-Zheng Ren, Peng-Cheng Wen, and Ya-Nan Sun. 2023. "Nobiletin Improves D-Galactose-Induced Aging Mice Skeletal Muscle Atrophy by Regulating Protein Homeostasis" Nutrients 15, no. 8: 1801. https://doi.org/10.3390/nu15081801
APA StyleWang, H. -H., Zhang, Y., Qu, T. -Q., Sang, X. -Q., Li, Y. -X., Ren, F. -Z., Wen, P. -C., & Sun, Y. -N. (2023). Nobiletin Improves D-Galactose-Induced Aging Mice Skeletal Muscle Atrophy by Regulating Protein Homeostasis. Nutrients, 15(8), 1801. https://doi.org/10.3390/nu15081801