

Antidiabetic Activity of Potential Probiotics Limosilactobacillus spp., Levilactobacillus spp., and Lacticaseibacillus spp. Isolated from Fermented Sugarcane Juice: A Comprehensive In Vitro and In Silico Study
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sugarcane Juice Fermentation, Isolation, and Preliminary Biochemical Characterization
2.2. Fermented Sugarcane Juice Strains Molecular Identification and Phylogenetic Analysis
2.3. Probiotic Properties
2.3.1. Assessment of Adhesion Capability
Cell Surface Hydrophobicity, Autoaggregation, and Coaggregation Assay
Human Buccal Epithelial Cells, and HT-29 Cells In Vitro Adhesion Assay
2.3.2. Tolerance Assay
Tolerance to Bile Salt in Acidic Conditions
Assay for Simulated Gastric Juice Tolerance
2.4. Antibacterial Activity
2.5. Antibiotic Susceptibility
2.6. Hemolytic Activity
2.7. Screening for Antioxidant Activity
2.8. Inhibitory Assay for Carbohydrate Hydrolyzing Enzymes
2.9. Profiling of Organic Acids by LCMS
2.10. Pass Pharmacological Analysis
2.11. Molecular Docking Simulation
2.12. Molecular Dynamics Simulation
2.13. Binding Free Energy Calculations
2.14. Statistical Analysis
3. Results and Discussion
3.1. Preliminary Biochemical Characterization
3.2. Molecular Identification of LAB
3.3. Probiotic Properties
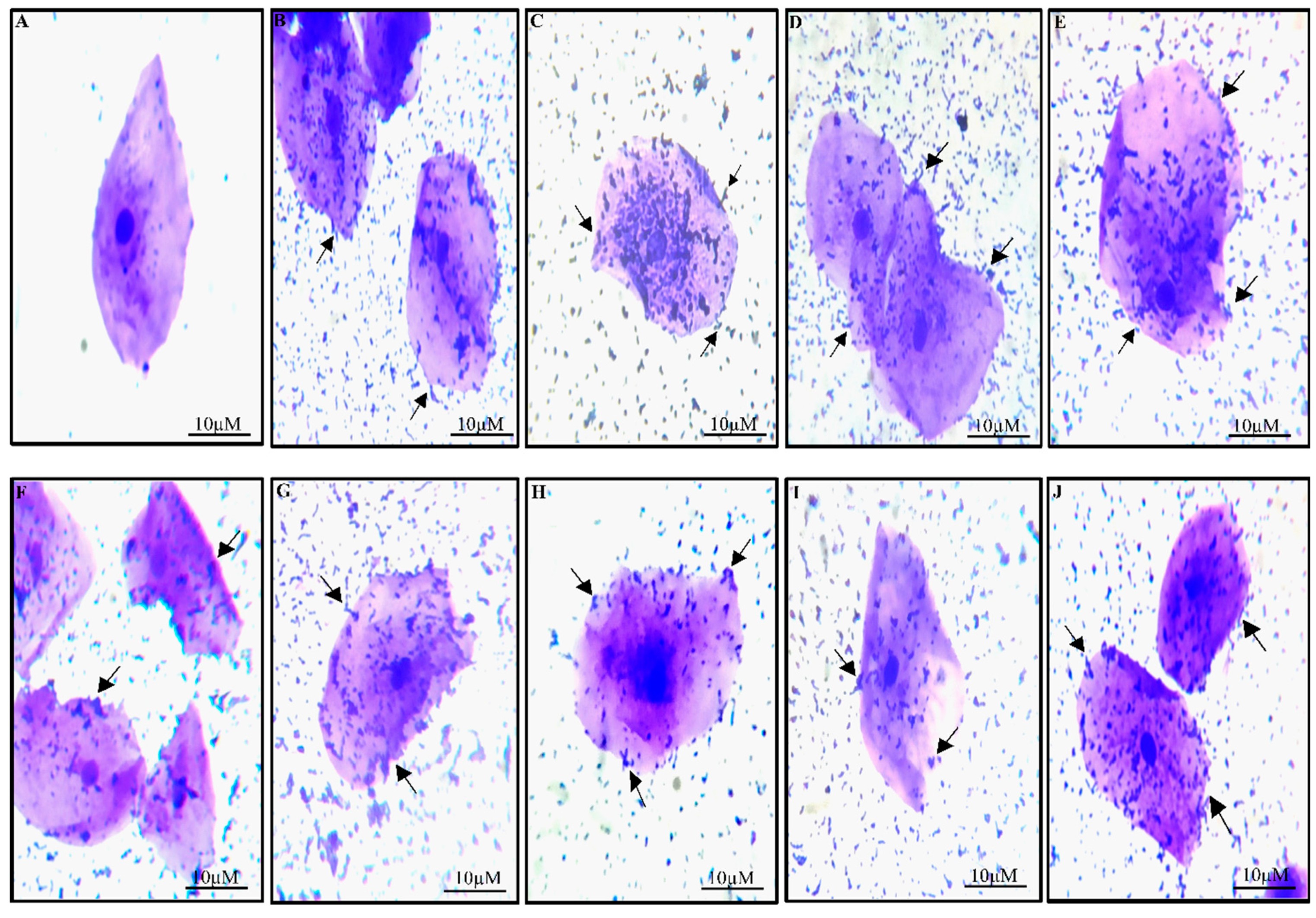
3.3.1. Adherence Assay
Hydrophobicity, Autoaggregation, and Coaggregation of Fermented Sugarcane Juice Isolates
Adhesion to HT-29 Cell Lines and Buccal Epithelial Cells
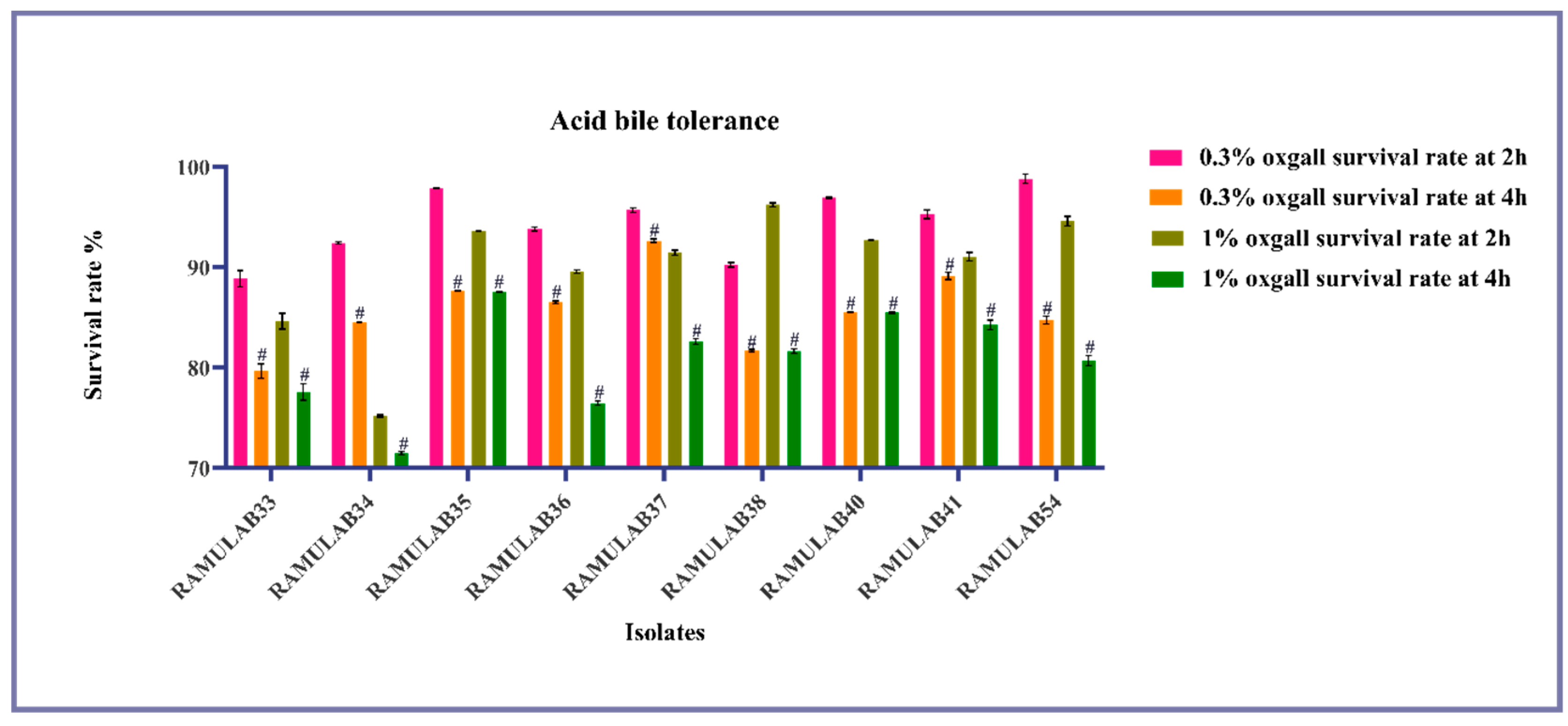
3.3.2. Tolerance Assay
Tolerance for Bile Salt in Acidic Conditions
Simulated Gastrointestinal Juice Tolerance Assay
3.4. Safety Assessments
3.5. Antibiotic Sensitivity
3.6. Hemolytic Assay
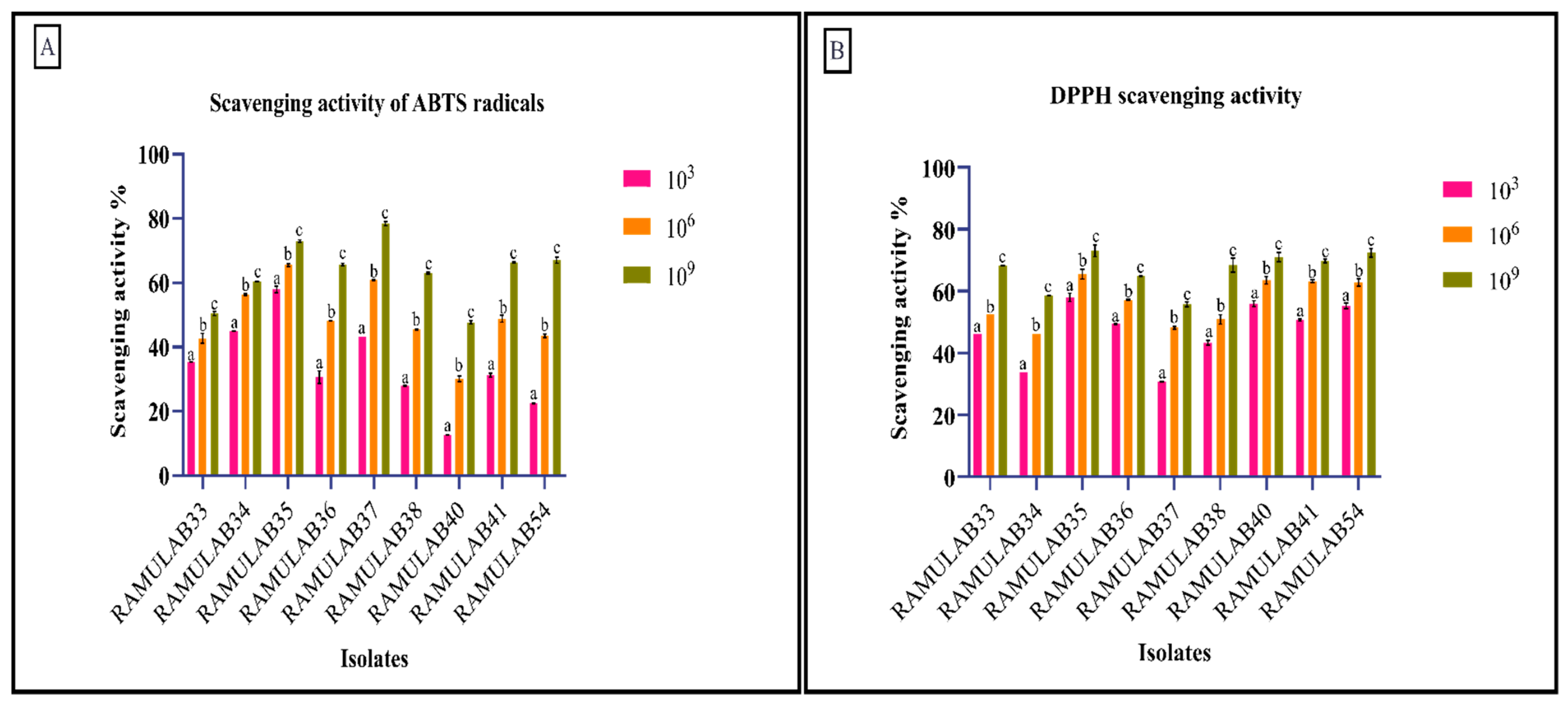
3.7. Antioxidant Assay
3.8. Inhibitory Assay for the Carbohydrate Hydrolyzing Enzymes (α-Glucosidase and α-Amylase)
3.9. Profiling of Organic Acid
3.10. Pass Pharmacological Potential Analysis
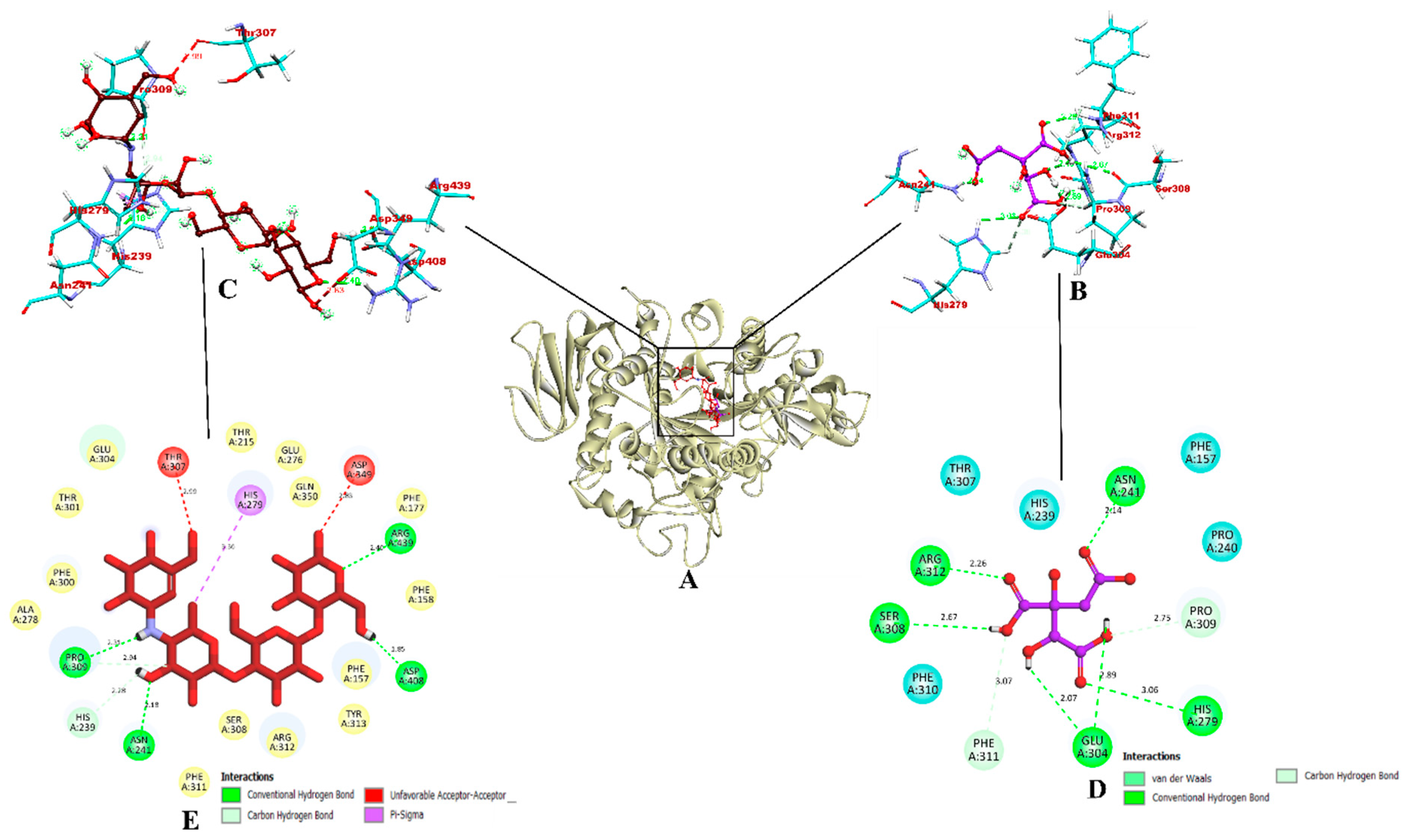
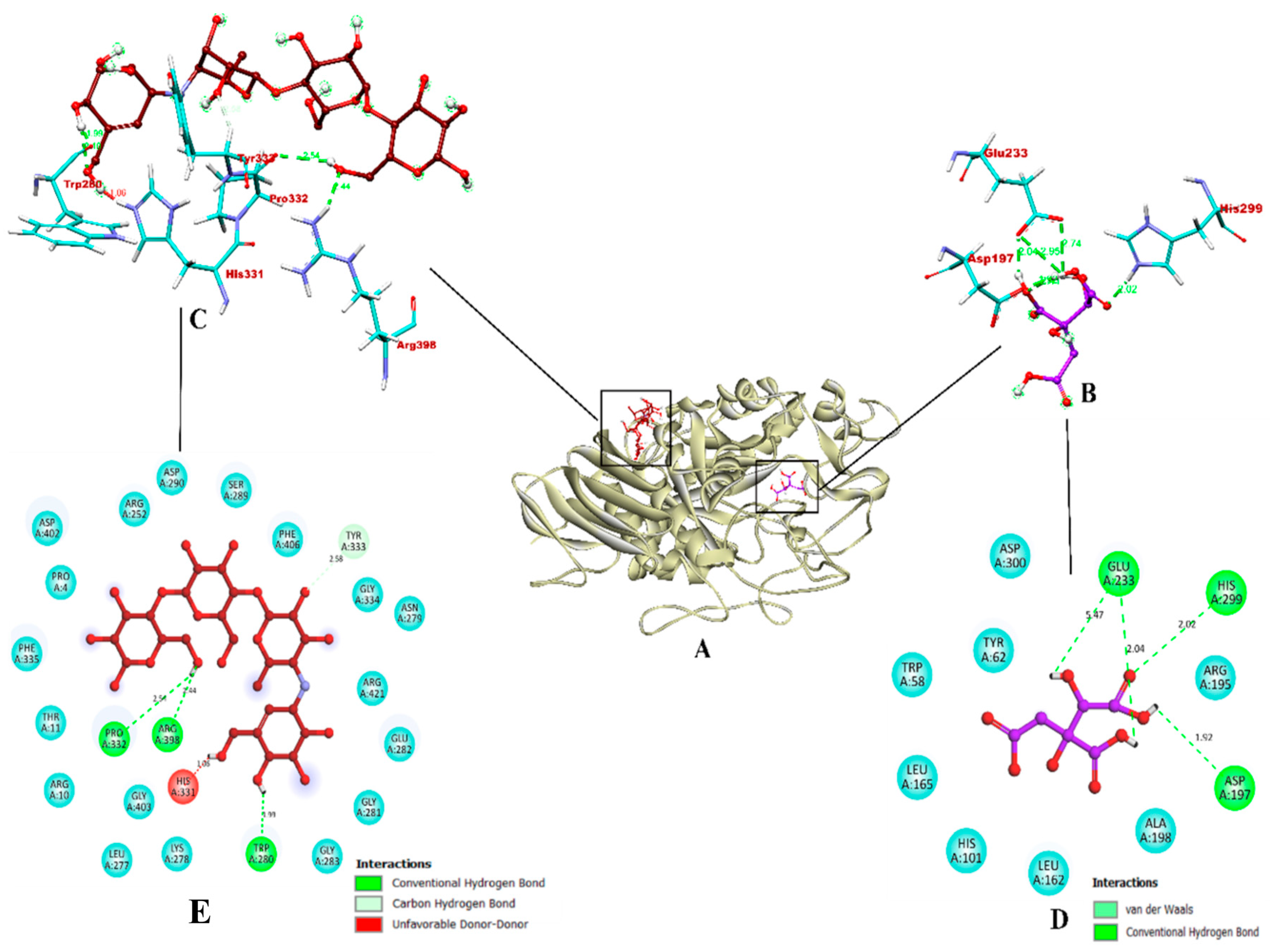
3.11. Molecular Docking Studies
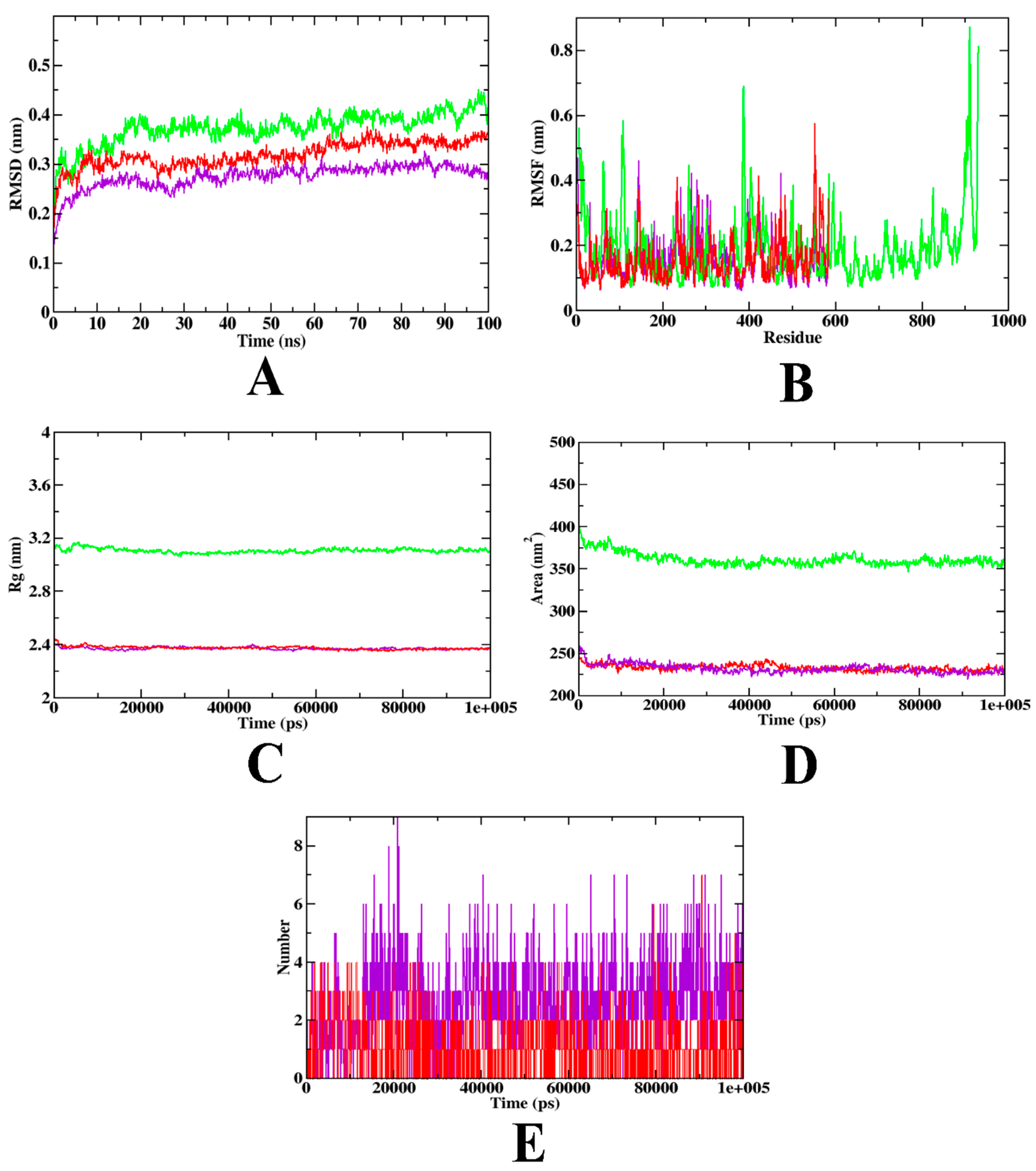
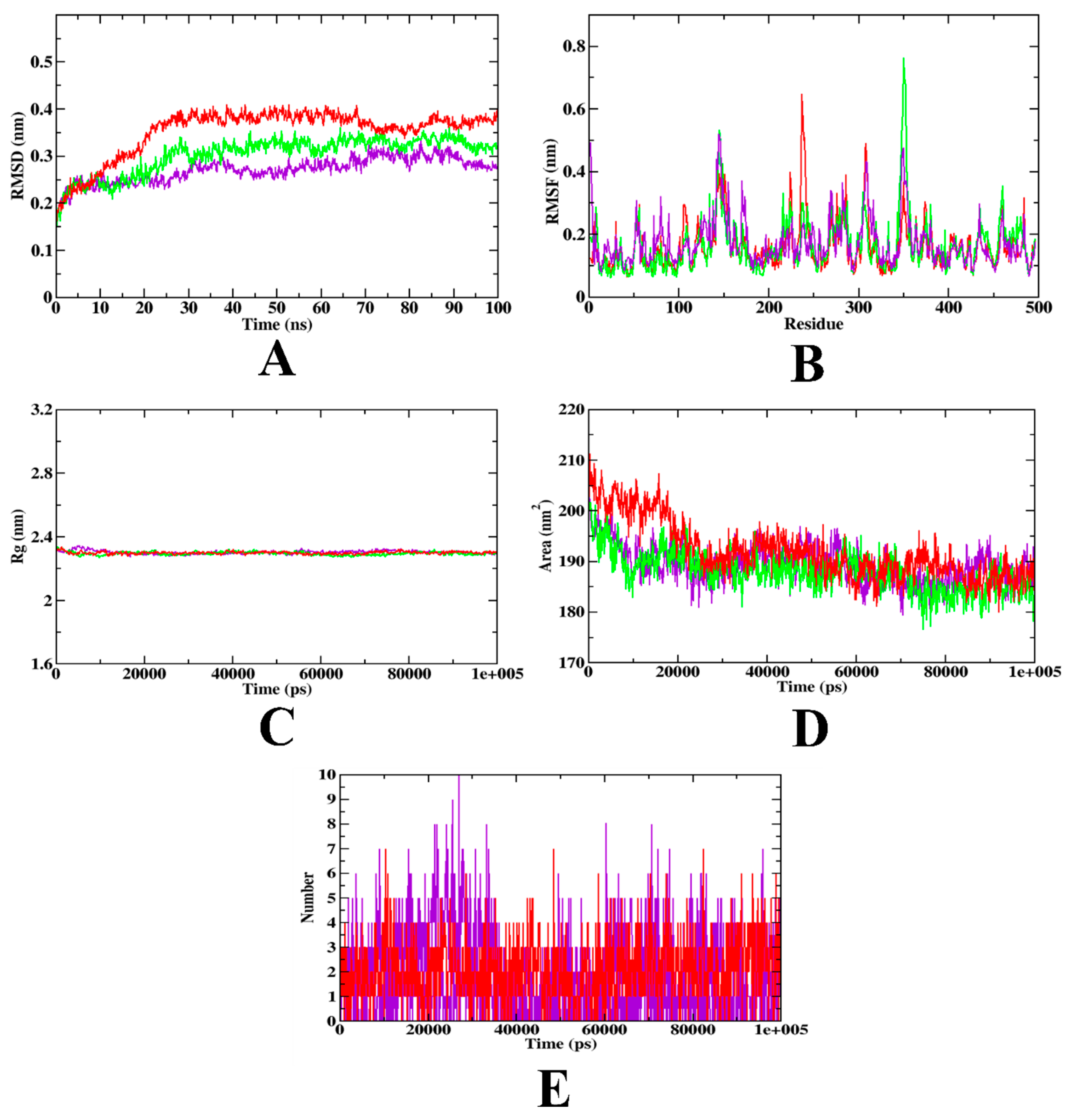
3.12. Molecular Dynamics Simulation
3.13. Binding Free Energy Calculations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramu, R.; Shirahatti, P.S.; Zameer, F.; Ranganatha, L.V.; Nagendra Prasad, M.N. Inhibitory Effect of Banana (Musa Sp. Var. Nanjangud Rasa Bale) Flower Extract and Its Constituents Umbelliferone and Lupeol on α-Glucosidase, Aldose Reductase and Glycation at Multiple Stages. S. Afr. J. Bot. 2014, 95, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, N.; Gao, P.; Kondapally Seshasai, S.R.; Gobin, R.; Kaptoge, S.; di Angelantonio, E.; Ingelsson, E.; Lawlor, D.A.; Selvin, E.; Stampfer, M.; et al. Diabetes Mellitus, Fasting Blood Glucose Concentration, and Risk of Vascular Disease: A Collaborative Meta-Analysis of 102 Prospective Studies. Lancet 2010, 375, 2215–2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salis, S.; Virmani, A.; Priyambada, L.; Mohan, M.; Hansda, K.; de Beaufort, C. ‘Old Is Gold’: How Traditional Indian Dietary Practices Can Support Pediatric Diabetes Management. Nutrients 2021, 13, 4427. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and Prebiotics in Intestinal Health and Disease: From Biology to the Clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Namai, F.; Shigemori, S.; Ogita, T.; Sato, T.; Shimosato, T. Microbial Therapeutics for Acute Colitis Based on Genetically Modified Lactococcus Lactis Hypersecreting IL-1Ra in Mice. Exp. Mol. Med. 2020, 52, 1627–1636. [Google Scholar] [CrossRef]
- Kumar, V.; Ramu, R.; Shirahatti, P.S.; Kumari, V.B.C.; Sushma, P.; Mandal, S.P.; Patil, S.M. α-Glucosidase, α-Amylase Inhibition, Kinetics and Docking Studies of Novel (2-Chloro-6-(Trifluoromethyl)Benzyloxy)Arylidene) Based Rhodanine and Rhodanine Acetic Acid Derivatives. ChemistrySelect 2021, 6, 9637–9644. [Google Scholar] [CrossRef]
- Han, J.L.; Lin, H.L. Intestinal Microbiota and Type 2 Diabetes: From Mechanism Insights to Therapeutic Perspective. World J. Gastroenterol. WJG 2014, 20, 17737. [Google Scholar] [CrossRef]
- Oh, J.; Jo, S.H.; Kim, J.S.; Ha, K.S.; Lee, J.Y.; Choi, H.Y.; Yu, S.Y.; Kwon, Y.I.; Kim, Y.C. Selected Tea and Tea Pomace Extracts Inhibit Intestinal α-Glucosidase Activity in Vitro and Postprandial Hyperglycemia in Vivo. Int. J. Mol. Sci. 2015, 16, 8811–8825. [Google Scholar] [CrossRef]
- Kim, S.H.; Jo, S.H.; Kwon, Y.I.; Hwang, J.K. Effects of Onion (Allium Cepa L.) Extract Administration on Intestinal α-Glucosidases Activities and Spikes in Postprandial Blood Glucose Levels in SD Rats Model. Int. J. Mol. Sci. 2011, 12, 3757–3769. [Google Scholar] [CrossRef] [Green Version]
- Panwar, H.; Calderwood, D.; Grant, I.R.; Grover, S.; Green, B.D. Lactobacillus Strains Isolated from Infant Faeces Possess Potent Inhibitory Activity against Intestinal Alpha- and Beta-Glucosidases Suggesting Anti-Diabetic Potential. Eur. J. Nutr. 2014, 53, 1465–1474. [Google Scholar] [CrossRef]
- Reale, A.; di Renzo, T.; Rossi, F.; Zotta, T.; Iacumin, L.; Preziuso, M.; Parente, E.; Sorrentino, E.; Coppola, R. Tolerance of Lactobacillus Casei, Lactobacillus Paracasei and Lactobacillus Rhamnosus Strains to Stress Factors Encountered in Food Processing and in the Gastro-Intestinal Tract. LWT 2015, 60, 721–728. [Google Scholar] [CrossRef]
- Saad, M.J.A.; Santos, A.; Prada, P.O. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, R.; Dey, D.K.; Vij, R.; Meena, S.; Kapila, R.; Kapila, S. Evaluation of Anti-Diabetic Attributes of Lactobacillus Rhamnosus MTCC: 5957, Lactobacillus Rhamnosus MTCC: 5897 and Lactobacillus Fermentum MTCC: 5898 in Streptozotocin Induced Diabetic Rats. Microb. Pathog. 2018, 125, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Ding, W.; Liu, Y. Anti-Diabetic Effects of Emodin Involved in the Activation of PPARγ on High-Fat Diet-Fed and Low Dose of Streptozotocin-Induced Diabetic Mice. Fitoterapia 2010, 81, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, E.; Yin, B.; Fang, D.; Chen, P.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Effects of Lactobacillus Casei CCFM419 on Insulin Resistance and Gut Microbiota in Type 2 Diabetic Mice. Benef. Microbes 2017, 8, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Ramu, R.; Shirahatti, P.S.; TH, D.; Bajpe, S.N.; SreepathI, N.; Patil, S.M.; MN, N.P. Investigating Musa Paradisiaca (Var. Nanjangud Rasa Bale) Pseudostem in Preventing Hyperglycemia along with Improvement of Diabetic Complications. J. Appl. Biol. Biotechnol. 2022, 10, 56–65. [Google Scholar] [CrossRef]
- Li, X.; Wang, N.; Yin, B.; Fang, D.; Jiang, T.; Fang, S.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Effects of Lactobacillus Plantarum CCFM0236 on Hyperglycaemia and Insulin Resistance in High-Fat and Streptozotocin-Induced Type 2 Diabetic Mice. J. Appl. Microbiol. 2016, 121, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.; Bora, J.; Sharma, V. Growth Studies of Potentially Probiotic Lactic Acid Bacteria (Lactobacillus Plantarum, Lactobacillus Acidophilus, and Lactobacillus Casei) in Carrot and Beetroot Juice Substrates. J. Food Process Preserv. 2019, 43, 1–8. [Google Scholar] [CrossRef]
- Gupta, A.; Tiwari, S.K. Probiotic Potential of Lactobacillus Plantarum LD1 Isolated from Batter of Dosa, a South Indian Fermented Food. Probiotics Antimicrob. Proteins 2014, 6, 73–81. [Google Scholar] [CrossRef]
- Palanisamy, B.D.; Rajendran, V.; Sathyaseelan, S.; Bhat, R.; Venkatesan, B.P. Enhancement of Nutritional Value of Finger Millet-Based Food (Indian Dosa) by Co-Fermentation with Horse Gram Flour. Int. J. Food Sci. Nutr. 2012, 63, 5–15. [Google Scholar] [CrossRef]
- Jeon, H.H.; Kim, K.H.; Chun, B.H.; Ryu, B.H.; Han, N.S.; Jeon, C.O. A Proposal of Leuconostoc Mesenteroides Subsp. Jonggajibkimchii Subsp. Nov. and Reclassification of Leuconostoc Mesenteroides Subsp. Suionicum (Gu et al., 2012) as Leuconostoc Suionicum Sp. Nov. Based on Complete Genome Sequences. Int. J. Syst. Evol. Microbiol. 2017, 67, 2225–2230. [Google Scholar] [CrossRef] [PubMed]
- Nel, S.; Davis, S.B.; Endo, A.; Dicks, L.M.T. Differentiation between Bacillus Amyloliquefaciens and Bacillus Subtilis Isolated from a South African Sugarcane Processing Factory Using ARDRA and RpoB Gene Sequencing. Arch. Microbiol. 2019, 201, 1453–1457. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Lal, U.R.; Mukhtar, H.M.; Singh, P.S.; Shah, G.; Dhawan, R.K. Phytochemical Profile of Sugarcane and Its Potential Health Aspects. Pharm. Rev. 2015, 9, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molavian, M.; Ghorbani, G.R.; Rafiee, H.; Beauchemin, K.A. Substitution of Wheat Straw with Sugarcane Bagasse in Low-Forage Diets Fed to Mid-Lactation Dairy Cows: Milk Production, Digestibility, and Chewing Behavior. J. Dairy Sci. 2020, 103, 8034–8047. [Google Scholar] [CrossRef]
- Pattnaik, B.; Sarangi, P.K.; Jena, P.K.; Sahoo, H.P.; Bhatia, L. Production of Phenolic Flavoring Compounds from Sugarcane Bagasse by Lactobacillus Acidophilus MTCC 10307. Arch. Microbiol. 2021, 204, 23. [Google Scholar] [CrossRef]
- So, S.; Cherdthong, A.; Wanapat, M. Improving Sugarcane Bagasse Quality as Ruminant Feed with Lactobacillus, Cellulase, and Molasses. J. Anim. Sci. Technol. 2020, 62, 648–658. [Google Scholar] [CrossRef]
- Nel, S.; Davis, S.B.; Endo, A.; Dicks, L.M.T. Phylogenetic Analysis of Leuconostoc and Lactobacillus Species Isolated from Sugarcane Processing Streams. Microbiologyopen 2020, 9, e1065. [Google Scholar] [CrossRef]
- da Silva, L.D.; Pereira, O.G.; Roseira, J.P.S.; Agarussi, M.C.N.; da Silva, V.P.; da Silva, T.C.; Leandro, E.D.S.; de Paula, R.A.; Santos, S.A.; Ribeiro, K.G.; et al. Effect of Wild Lactobacillus Buchneri Strains on the Fermentation Profile and Microbial Populations of Sugarcane Silage. Recent Pat. Food Nutr. Agric. 2020, 11, 63–68. [Google Scholar] [CrossRef]
- Qiao, H.; Chen, L.; Yang, J.; Zhi, W.; Chen, R.; Lu, T.; Tan, H.; Sheng, Z. Effect of Lactic Acid Bacteria on Bacterial Community Structure and Characteristics of Sugarcane Juice. Foods 2022, 11, 3134. [Google Scholar] [CrossRef]
- Kitahara, M.; Sakamoto, M.; Benno, Y. Lactobacillus Similis Sp. Nov., Isolated from Fermented Cane Molasses. Int. J. Syst. Evol. Microbiol. 2010, 60, 187–190. [Google Scholar] [CrossRef]
- Ellis, T.P.; Wright, A.G.; Clifton, P.M.; Ilag, L.L. Postprandial Insulin and Glucose Levels Are Reduced in Healthy Subjects When a Standardised Breakfast Meal Is Supplemented with a Filtered Sugarcane Molasses Concentrate. Eur. J. Nutr. 2016, 55, 2365–2376. [Google Scholar] [CrossRef] [PubMed]
- Ayuningtyas, R.A.; Wijayanti, C.; Hapsari, N.R.P.; Sari, B.F.P. Subandi Preliminary Study: The Use of Sugarcane Juice to Replace White Sugar in an Effort to Overcome Diabetes Mellitus. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Malang, East Java, Indonesia, 26–27 August 2019; Institute of Physics Publishing: Bristol, UK, 2020; Volume 475. [Google Scholar]
- Abduldileep, S.; Narayanasamy, R.; Usharani, D.; Singh, A.; Rajasekharan, R. A Bioactive Polypeptide from Sugarcane Selectively Inhibits Intestinal Sucrase. Int. J. Biol. Macromol. 2020, 156, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Kumari V. B., C.; Huligere, S.S.; Shbeer, A.M.; Ageel, M.; Jayanthi, M.K.; Jagadeep, C.S.; Ramu, R. Probiotic Potential Lacticaseibacillus Casei and Limosilactobacillus Fermentum Strains Isolated from Dosa Batter Inhibit α-Glucosidase and α-Amylase Enzymes. Microorganisms 2022, 10, 1195. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Tuo, Y.; Yu, H.; Ai, L.; Wu, Z.; Guo, B.; Chen, W. Aggregation and Adhesion Properties of 22 Lactobacillus Strains. J. Dairy Sci. 2013, 96, 4252–4257. [Google Scholar] [CrossRef] [Green Version]
- Kumari, V.B.C.; Huligere, S.S.; Ramu, R.; Naik Bajpe, S.; Sreenivasa, M.Y.; Silina, E.; Stupin, V.; Achar, R.R. Evaluation of Probiotic and Antidiabetic Attributes of Lactobacillus Strains Isolated From Fermented Beetroot. Front. Microbiol. 2022, 13, 911243. [Google Scholar] [CrossRef]
- Begley, M.; Gahan, C.G.M.; Hill, C. The Interaction between Bacteria and Bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [Green Version]
- Faye, T.; Tamburello, A.; Vegarud, G.E.; Skeie, S. Survival of Lactic Acid Bacteria from Fermented Milks in an in Vitro Digestion Model Exploiting Sequential Incubation in Human Gastric and Duodenum Juice. J. Dairy Sci. 2012, 95, 558–566. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Zhang, Y.; Zhang, Y.; Liu, Y.; Wang, S.; Dong, X.; Wang, Y.; Zhang, H. Screening of Potential Probiotic Properties of Lactobacillus Fermentum Isolated from Traditional Dairy Products. Food Control 2010, 21, 695–701. [Google Scholar] [CrossRef]
- Li, S.; Huang, R.; Shah, N.P.; Tao, X.; Xiong, Y.; Wei, H. Antioxidant and Antibacterial Activities of Exopolysaccharides from Bifidobacterium Bifidum WBIN03 and Lactobacillus Plantarum R315. J. Dairy Sci. 2014, 97, 7334–7343. [Google Scholar] [CrossRef] [Green Version]
- Hana, J.; Chena, D.; Lic, S.; Lia, X.; Zhoud, W.W.; Zhangb, B.; Jiaa, Y. Antibiotic Susceptibility of Potentially Probiotic Lactobacillus Strains. Ital. J. Food Sci. 2015, 27, 282–289. [Google Scholar]
- Husain, F.; Duraisamy, S.; Balakrishnan, S.; Ranjith, S.; Chidambaram, P.; Kumarasamy, A. Phenotypic Assessment of Safety and Probiotic Potential of Native Isolates from Marine Fish Moolgarda Seheli towards Sustainable Aquaculture. Biologia (Bratisl) 2022, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Silva-Rivas, R.; Bailon-Moscoso, N.; Cartuche, L.; Romero-Benavides, J.C. The Antioxidant and Hypoglycemic Properties and Phytochemical Profile of Clusia Latipes Extracts. Pharmacogn. J. 2020, 12, 144–149. [Google Scholar] [CrossRef] [Green Version]
- Shimojo, Y.; Ozawa, Y.; Toda, T.; Igami, K.; Shimizu, T. Probiotic Lactobacillus Paracasei A221 Improves the Functionality and Bioavailability of Kaempferol-Glucoside in Kale by Its Glucosidase Activity. Sci. Rep. 2018, 8, 9239. [Google Scholar] [CrossRef] [Green Version]
- Talamond, P.; Desseaux, V.; Moreau, Y.; Santimone, M.; Marchis-Mouren, G. Isolation, Characterization and Inhibition by Acarbose of the α-Amylase from Lactobacillus Fermentum: Comparison with Lb. Manihotivorans and Lb. Plantarum Amylases. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2002, 133, 351–360. [Google Scholar] [CrossRef]
- Kwun, S.Y.; Bae, Y.W.; Yoon, J.A.; Park, E.H.; Kim, M.D. Isolation of Acid Tolerant Lactic Acid Bacteria and Evaluation of α-Glucosidase Inhibitory Activity. Food Sci. Biotechnol. 2020, 29, 1125. [Google Scholar] [CrossRef]
- Patil, S.M.; Phanindra, B.; Shirahatti, P.S.; Martiz, R.M.; Sajal, H.; Babakr, A.T.; Ramu, R. Computational Approaches to Define Poncirin from Magnolia Champaka Leaves as a Novel Multi-Target Inhibitor of SARS-CoV-2. J. Biomol. Struct. Dyn. 2023, 1–20. [Google Scholar] [CrossRef]
- Maradesha, T.; Patil, S.M.; Al-Mutairi, K.A.; Ramu, R.; Madhunapantula, S.V.; Alqadi, T. Inhibitory Effect of Polyphenols from the Whole Green Jackfruit Flour against α-Glucosidase, α-Amylase, Aldose Reductase and Glycation at Multiple Stages and Their Interaction: Inhibition Kinetics and Molecular Simulations. Molecules 2022, 27, 1888. [Google Scholar] [CrossRef]
- Patil, S.M.; Martiz, R.M.; Ramu, R.; Shirahatti, P.S.; Prakash, A.; Kumar, B.R.P.; Kumar, N. Evaluation of Flavonoids from Banana Pseudostem and Flower (Quercetin and Catechin) as Potent Inhibitors of α-Glucosidase: An in Silico Perspective. J. Biomol. Struct. Dyn. 2021, 40, 12491–12505. [Google Scholar] [CrossRef]
- Pushpa, V.H.; Jayanthi, M.K.; Rashmi, H.R.; Shivamurthy, V.K.N.; Patil, S.M.; Shirahatti, P.S.; Ramu, R. New Insights on the Phytochemical Intervention for the Treatment of Neuropsychiatric Disorders Using the Leaves of Michelia Champaca: An in Vivo and in Silico Approach. Pharm. Biol. 2022, 60, 1656–1668. [Google Scholar] [CrossRef]
- Martiz, R.; Patil, S.M.; Martiz, R.M.; Ramu, R.; Shirahatti, P.S.; Prakash, A.; Chandra, J.; Ranganatha, V.L.; Mary Martiz, R.; Lakshmi Ranganatha, V. In Silico Identification of Novel Benzophenone–Coumarin Derivatives as SARS-CoV-2 RNA-Dependent RNA Polymerase (RdRp) Inhibitors. J. Biomol. Struct. Dyn. 2021, 40, 13032–13048. [Google Scholar] [CrossRef]
- Martiz, R.M.; Patil, S.M.; Thirumalapura Hombegowda, D.; Shbeer, A.M.; Alqadi, T.; Al-Ghorbani, M.; Ramu, R.; Prasad, A. Phyto-Computational Intervention of Diabetes Mellitus at Multiple Stages Using Isoeugenol from Ocimum Tenuiflorum: A Combination of Pharmacokinetics and Molecular Modelling Approaches. Molecules 2022, 27, 6222. [Google Scholar] [CrossRef]
- Martiz, R.M.; Patil, S.M.; Ramu, R.; Jayanthi, M.K.; Ashwini, P.; Ranganatha, L.V.; Khanum, S.A.; Silina, E.; Stupin, V.; Achar, R.R. Discovery of Novel Benzophenone Integrated Derivatives as Anti-Alzheimer’s Agents Targeting Presenilin-1 and Presenilin-2 Inhibition: A Computational Approach. PLoS ONE 2022, 17, 0265022. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.M.; Maruthi, K.; Bajpe, S.N.; Vyshali, V.; Sushmitha, S.; Akhila, C.; Ramu, R. Comparative Molecular Docking and Simulation Analysis of Molnupiravir and Remdesivir with SARS-CoV-2 RNA Dependent RNA Polymerase (RdRp). Bioinformation 2021, 17, 932. [Google Scholar] [CrossRef]
- Sajal, H.; Patil, S.M.; Raj, R.; Shbeer, A.M.; Ageel, M.; Ramu, R. Computer-Aided Screening of Phytoconstituents from Ocimum Tenuiflorum against Diabetes Mellitus Targeting DPP4 Inhibition: A Combination of Molecular Docking, Molecular Dynamics, and Pharmacokinetics Approaches. Molecules 2022, 27, 5133. [Google Scholar] [CrossRef]
- Divyashree, S.; Anjali, P.G.; Somashekaraiah, R.; Sreenivasa, M.Y. Probiotic Properties of Lactobacillus Casei—MYSRD 108 and Lactobacillus Plantarum-MYSRD 71 with Potential Antimicrobial Activity against Salmonella Paratyphi. Biotechnol. Rep. 2021, 32, e00672. [Google Scholar] [CrossRef]
- Narendranath, N.V.; Power, R. Relationship between PH and Medium Dissolved Solids in Terms of Growth and Metabolism of Lactobacilli and Saccharomyces Cerevisiae during Ethanol Production. Appl. Environ. Microbiol. 2005, 71, 2239–2243. [Google Scholar] [CrossRef] [Green Version]
- Rokana, N.; Singh, B.P.; Thakur, N.; Sharma, C.; Gulhane, R.D.; Panwar, H. Screening of Cell Surface Properties of Potential Probiotic Lactobacilli Isolated from Human Milk. J. Dairy Res. 2018, 85, 347–354. [Google Scholar] [CrossRef]
- Kim, J.H.; Jang, H.J.; Lee, N.K.; Paik, H.D. Antibacterial and Antibiofilm Effect of Cell-Free Supernatant of Lactobacillus Brevis KCCM 202399 Isolated from Korean Fermented Food against Streptococcus Mutans KCTC 5458. J. Microbiol. Biotechnol. 2022, 32, 56–63. [Google Scholar] [CrossRef]
- Singh, T.P.; Kaur, G.; Malik, R.K.; Schillinger, U.; Guigas, C.; Kapila, S. Characterization of Intestinal Lactobacillus Reuteri Strains as Potential Probiotics. Probiotics Antimicrob. Proteins 2012, 4, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, Y.; Cui, H.; Li, Y.; Sun, Y.; Qiu, H.J. Characterization of Lactic Acid Bacteria Isolated From the Gastrointestinal Tract of a Wild Boar as Potential Probiotics. Front. Vet. Sci. 2020, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorov, S.D.; Botes, M.; Guigas, C.; Schillinger, U.; Wiid, I.; Wachsman, M.B.; Holzapfel, W.H.; Dicks, L.M.T. Boza, a Natural Source of Probiotic Lactic Acid Bacteria. J. Appl. Microbiol. 2008, 104, 465–477. [Google Scholar] [CrossRef]
- Vlková, E.; Rada, V.; Šmehilová, M.; Killer, J. Auto-Aggregation and Co-Aggregation Ability in Bifidobacteria and Clostridia. Folia Microbiol. 2008, 53, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E.Z. Antimicrobial and Anti-Adhesive Properties of Biosurfactant Produced by Lactobacilli Isolates, Biofilm Formation and Aggregation Ability. J. Gen. Appl. Microbiol. 2013, 59, 425–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanani, A.S.; Bagchi, T. Lactobacillus Plantarum CS24.2 Prevents Escherichia Coli Adhesion to HT-29 Cells and Also down-Regulates Enteropathogen-Induced Tumor Necrosis Factor-α and Interleukin-8 Expression. Microbiol. Immunol. 2013, 57, 309–315. [Google Scholar] [CrossRef]
- Duary, R.K.; Rajput, Y.S.; Batish, V.K.; Grover, S. Assessing the Adhesion of Putative Indigenous Probiotic Lactobacilli to Human Colonic Epithelial Cells. Indian J. Med. Res. 2011, 134, 664. [Google Scholar] [CrossRef]
- Somashekaraiah, R.; Shruthi, B.; Deepthi, B.V.; Sreenivasa, M.Y. Probiotic Properties of Lactic Acid Bacteria Isolated from Neera: A Naturally Fermenting Coconut Palm Nectar. Front. Microbiol. 2019, 10, 1382. [Google Scholar] [CrossRef]
- Fonseca, H.C.; de Sousa Melo, D.; Ramos, C.L.; Dias, D.R.; Schwan, R.F. Probiotic Properties of Lactobacilli and Their Ability to Inhibit the Adhesion of Enteropathogenic Bacteria to Caco-2 and HT-29 Cells. Probiotics Antimicrob. Proteins 2020, 13, 102–112. [Google Scholar] [CrossRef]
- Wang, J.; Guo, Z.; Zhang, Q.; Yan, L.; Chen, W.; Liu, X.M.; Zhang, H.P. Fermentation Characteristics and Transit Tolerance of Probiotic Lactobacillus Casei Zhang in Soymilk and Bovine Milk during Storage. J. Dairy Sci. 2009, 92, 2468–2476. [Google Scholar] [CrossRef] [Green Version]
- Vidhyasagar, V.; Jeevaratnam, K. Evaluation of Pediococcus Pentosaceus Strains Isolated from Idly Batter for Probiotic Properties in Vitro. J. Funct. Foods 2013, 5, 235–243. [Google Scholar] [CrossRef]
- Jiang, H.; Tang, X.; Zhou, Q.; Zou, J.; Li, P.; Breukink, E.; Gu, Q. Plantaricin NC8 from Lactobacillus Plantarum Causes Cell Membrane Disruption to Micrococcus Luteus without Targeting Lipid II. Appl. Microbiol. Biotechnol. 2018, 102, 7465–7473. [Google Scholar] [CrossRef]
- Li, J.; Aroutcheva, A.A.; Faro, S.; Chikindas, M.L. Mode of Action of Lactocin 160, a Bacteriocin from Vaginal Lactobacillus Rhamnosus. Infect. Dis. Obs. Gynecol. 2005, 13, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Mollendorff, J.W.; Todorov, S.D.; Dicks, L.M.T. Comparison of Bacteriocins Produced by Lactic-Acid Bacteria Isolated from Boza, a Cereal-Based Fermented Beverage from the Balkan Peninsula. Curr. Microbiol. 2006, 53, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Serata, M.; Yasuda, E.; Sako, T. Effect of Superoxide Dismutase and Manganese on Superoxide Tolerance in Lactobacillus Casei Strain Shirota and Analysis of Multiple Manganese Transporters. Biosci. Microbiota Food Health 2018, 37, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Jamuna, M.; Jeevaratnam, K. Isolation and Characterization of Lactobacilli from Some Traditional Fermented Foods and Evaluation of the Bacteriocins. J. Gen. Appl. Microbiol. 2004, 50, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Temmerman, R.; Pot, B.; Huys, G.; Swings, J. Identification and Antibiotic Susceptibility of Bacterial Isolates from Probiotic Products. Int. J. Food Microbiol. 2003, 81, 1–10. [Google Scholar] [CrossRef]
- Georgieva, R.; Yocheva, L.; Tserovska, L.; Zhelezova, G.; Stefanova, N.; Atanasova, A.; Danguleva, A.; Ivanova, G.; Karapetkov, N.; Rumyan, N.; et al. Antimicrobial Activity and Antibiotic Susceptibility of Lactobacillus and Bifidobacterium Spp. Intended for Use as Starter and Probiotic Cultures. Biotechnol. Biotechnol. Equip. 2015, 29, 84–91. [Google Scholar] [CrossRef]
- Nassar, M.S.M.; Hazzah, W.A.; Bakr, W.M.K. Evaluation of Antibiotic Susceptibility Test Results: How Guilty a Laboratory Could Be? J. Egypt. Public Health Assoc. 2019, 94, 4. [Google Scholar] [CrossRef] [Green Version]
- Yasmin, I.; Saeed, M.; Khan, W.A.; Khaliq, A.; Chughtai, M.F.J.; Iqbal, R.; Tehseen, S.; Naz, S.; Liaqat, A.; Mehmood, T.; et al. In Vitro Probiotic Potential and Safety Evaluation (Hemolytic, Cytotoxic Activity) of Bifidobacterium Strains Isolated from Raw Camel Milk. Microorganisms 2020, 8, 354. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.J.; Jung, D.S. Evaluation of Probiotic Properties of Lactobacillus and Pediococcus Strains Isolated from Omegisool, a Traditionally Fermented Millet Alcoholic Beverage in Korea. LWT Food Sci. Technol. 2015, 63, 437–444. [Google Scholar] [CrossRef]
- Wang, X.; Wang, W.; Lv, H.; Zhang, H.; Liu, Y.; Zhang, M.; Wang, Y.; Tan, Z. Probiotic Potential and Wide-Spectrum Antimicrobial Activity of Lactic Acid Bacteria Isolated from Infant Feces. Probiotics Antimicrob. Proteins 2021, 13, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Leopold, J.A. Antioxidants and Coronary Artery Disease: From Pathophysiology to Preventive Therapy. Coron Artery Dis. 2015, 26, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathiyaseelan, A.; Saravanakumar, K.; Han, K.; Naveen, K.V.; Wang, M.H. Antioxidant and Antibacterial Effects of Potential Probiotics Isolated from Korean Fermented Foods. Int. J. Mol. Sci. 2022, 23, 62. [Google Scholar] [CrossRef]
- Kim, K.T.; Kim, J.W.; Kim, S.I.; Kim, S.; Nguyen, T.H.; Kang, C.H. Antioxidant and Anti-Inflammatory Effect and Probiotic Properties of Lactic Acid Bacteria Isolated from Canine and Feline Feces. Microorganisms 2021, 9, 1971. [Google Scholar] [CrossRef]
- Chooruk, A.; Piwat, S.; Teanpaisan, R. Antioxidant Activity of Various Oral Lactobacillus Strains. J. Appl. Microbiol. 2017, 123, 271–279. [Google Scholar] [CrossRef]
- Coman, C.; Ruginǎ, O.D.; Socaciu, C. Plants and Natural Compounds with Antidiabetic Action. Not Bot. Horti. Agrobot. Cluj Napoca 2012, 40, 314–325. [Google Scholar] [CrossRef] [Green Version]
- Giles-rivas, D.; Estrada-soto, S.; Aguilar-guadarrama, A.B.; Almanza-pérez, J.; García-jiménez, S.; Colín-lozano, B.; Navarrete-vázquez, G.; Villalobos-molina, R. Antidiabetic e Ff Ect of Cordia Morelosana, Chemical and Pharmacological Studies. J. Ethnopharmacol. 2020, 251, 112543. [Google Scholar] [CrossRef]
- Yadav, H.; Jain, S.; Sinha, P.R. Antidiabetic Effect of Probiotic Dahi Containing Lactobacillus Acidophilus and Lactobacillus Casei in High Fructose Fed Rats. Nutrition 2007, 23, 62–68. [Google Scholar] [CrossRef]
- Chaudhary, J.K.; Mudgal, S. Antidiabetic and Hypolipidemic Action of Finger Millet (Eleusine Coracana) Enriched Probiotic Fermented Milk: An in Vivo Rat Study. Food Technol. Biotechnol. 2020, 58, 192–202. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Antidiabetic Effect of Lactobacillus Casei CCFM0412 on Mice with Type 2 Diabetes Induced by a High-Fat Diet and Streptozotocin. Nutrition 2014, 30, 1061–1068. [Google Scholar] [CrossRef]
- Huligere, S.S.; Kumari, V.B.C.; Alqadi, T.; Kumar, S.; Cull, C.A.; Amachawadi, R.G.; Ramu, R. Isolation and Characterization of Lactic Acid Bacteria with Potential Probiotic Activity and Further Investigation of Their Activity by α-Amylase and α-Glucosidase Inhibitions of Fermented Batters. Front. Microbiol. 2022, 13, 1042263. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Screening for Potential New Probiotic Based on Probiotic Properties and α-Glucosidase Inhibitory Activity. Food Control 2014, 35, 65–72. [Google Scholar] [CrossRef]
- Son, S.H.; Jeon, H.L.; Yang, S.J.; Lee, N.K.; Paik, H.D. In Vitro Characterization of Lactobacillus Brevis KU15006, an Isolate from Kimchi, Reveals Anti-Adhesion Activity against Foodborne Pathogens and Antidiabetic Properties. Microb. Pathog. 2017, 112, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Lin, F.; Zhu, X.; Zhang, C.; Jiang, M.; Lu, Z. An Exopolysaccharide from Lactobacillus Plantarum H31 in Pickled Cabbage Inhibits Pancreas α-Amylase and Regulating Metabolic Markers in HepG2 Cells by AMPK/PI3K/Akt Pathway. Int. J. Biol. Macromol. 2020, 143, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Ramchandran, L.; Shah, N.P. Proteolytic Profiles and Angiotensin-I Converting Enzyme and α-Glucosidase Inhibitory Activities of Selected Lactic Acid Bacteria. J. Food Sci. 2008, 73, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Nuryana, I.; Andriani, A.; Lisdiyanti, P. Yopi Analysis of Organic Acids Produced by Lactic Acid Bacteria. IOP Conf. Ser. Earth Environ. Sci. 2019, 251, 012054. [Google Scholar] [CrossRef]
- Basa, E.L.U.; Julendra, H.; Abinawanto, A.; Sofyan, A.; Sophian, A. Analysis of Organic Acids from Lactobacillus Plantarum with Gas Chromatography-Mass Spectrometry (GC-MS). AIP Conf. Proc. 2019, 2168. [Google Scholar] [CrossRef]
- Zalán, Z.; Hudáček, J.; Štětina, J.; Chumchalová, J.; Halász, A. Production of Organic Acids by Lactobacillus Strains in Three Different Media. Eur. Food Res. Technol. 2010, 230, 395–404. [Google Scholar] [CrossRef]
- Prabhakaran, S.; Nivetha, N.; Patil, S.M.; Mary Martiz, R.; Ramu, R.; Sreenivasa, S.; Velmathi, S. One-Pot Three-Component Synthesis of Novel Phenyl-Pyrano-Thiazol-2-One Derivatives and Their Anti-Diabetic Activity Studies. Results Chem. 2022, 4, 100439. [Google Scholar] [CrossRef]
- Patil, S.M.; Martiz, R.M.; Satish, A.M.; Shbeer, A.M.; Ageel, M.; Al-Ghorbani, M.; Ranganatha, L.V.; Parameswaran, S.; Ramu, R. Discovery of Novel Coumarin Derivatives as Potential Dual Inhibitors against α-Glucosidase and α-Amylase for the Management of Post-Prandial Hyperglycemia via Molecular Modelling Approaches. Molecules 2022, 27, 3888. [Google Scholar] [CrossRef] [PubMed]
- Shivanna, C.; Patil, S.M.; Mallikarjunaswamy, C.; Ramu, R.; Akhileshwari, P.; Nagaraju, L.R.; Sridhar, M.A.; Khanum, S.A.; Ranganatha, V.L.; Silina, E.; et al. Synthesis, Characterization, Hirshfeld Surface Analysis, Crystal Structure and Molecular Modeling Studies of 1-(4-(Methoxy(Phenyl)Methyl)-2-Methylphenoxy)Butan-2-One Derivative as a Novel α-Glucosidase Inhibitor. Crystals 2022, 12, 960. [Google Scholar] [CrossRef]
- Maradesha, T.; Patil, S.M.; Phanindra, B.; Achar, R.R.; Silina, E.; Stupin, V.; Ramu, R. Multiprotein Inhibitory Effect of Dietary Polyphenol Rutin from Whole Green Jackfruit Flour Targeting Different Stages of Diabetes Mellitus: Defining a Bio-Computational Stratagem. Separations 2022, 9, 262. [Google Scholar] [CrossRef]
- Ganavi, D.; Ramu, R.; Kumar, V.; Patil, S.M.; Martiz, R.M.; Shirahatti, P.S.; Sathyanarayana, R.; Poojary, B.; Holla, B.S.; Poojary, V.; et al. In Vitro and in Silico Studies of Fluorinated 2,3-Disubstituted Thiazolidinone-Pyrazoles as Potential α-Amylase Inhibitors and Antioxidant Agents. Arch. Pharm. 2022, 355, 2100342. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.M.; Manu, G.; Shivachandra, J.C.; Anil Kumar, K.M.; Vigneswaran, J.; Ramu, R.; Shirahatti, P.S.; Lakshmi Ranganatha, V. Computational Screening of Benzophenone Integrated Derivatives (BIDs) Targeting the NACHT Domain of the Potential Target NLRP3 Inflammasome. Adv. Cancer Biol. Metastasis 2022, 5, 100056. [Google Scholar] [CrossRef]
- Gurupadaswamy, H.D.; Ranganatha, V.L.; Ramu, R.; Patil, S.M.; Khanum, S.A. Competent Synthesis of Biaryl Analogs via Asymmetric Suzuki–Miyaura Cross-Coupling for the Development of Anti-Inflammatory and Analgesic Agents. J. Iran. Chem. Soc. 2022, 19, 2421–2436. [Google Scholar] [CrossRef]
- Nivetha, N.; Martiz, R.M.; Patil, S.M.; Ramu, R.; Sreenivasa, S.; Velmathi, S. Benzodioxole Grafted Spirooxindole Pyrrolidinyl Derivatives: Synthesis, Characterization, Molecular Docking and Anti-Diabetic Activity. RSC Adv. 2022, 12, 24192–24207. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tests | Gram Staining | Catalase | Morphology | Temperature (°C) * | NaCl Concentration (%) * | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 | 10 | 37 | 45 | 50 | 2 | 4 | 6 | 8 | 10 | |||||
| Isolates | RAMULAB33 | Positive | Negative | Bacilli | A | A | P | A | A | P | P | A | A | A |
| RAMULAB34 | Positive | Negative | Bacilli | A | A | P | A | A | P | P | A | A | A | |
| RAMULAB35 | Positive | Negative | Bacilli | A | A | P | A | A | P | P | A | A | A | |
| RAMULAB36 | Positive | Negative | Bacilli | A | A | P | A | A | P | P | A | A | A | |
| RAMULAB37 | Positive | Negative | Bacilli | A | A | P | A | A | P | P | A | A | A | |
| RAMULAB38 | Positive | Negative | Bacilli | A | A | P | A | A | P | P | A | A | A | |
| RAMULAB40 | Positive | Negative | Bacilli | A | A | P | A | A | P | P | A | A | A | |
| RAMULAB41 | Positive | Negative | Bacilli | A | A | P | A | A | P | P | A | A | A | |
| RAMULAB54 | Positive | Negative | Bacilli | A | A | P | p | p | P | P | A | A | A | |
| Phenol Tolerance (CFU/ mL) * | Growth at Different pH (CFU/ mL) * | |||||
|---|---|---|---|---|---|---|
| Isolates | 0 h | 24 h | 2 | 4 | 6 | 7.4 |
| RAMULAB33 | 9.81 ± 0.02 c | 8.08 ± 0.11 c | 6.24 ± 0.11 d | 7.13 ± 0.01 c | 9.73 ± 0.02 d | 9.89 ± 0.12 c |
| RAMULAB34 | 8.66 ± 0.01 a | 8.15 ± 0.14 c | 6.02 ± 0.03 a | 7.16 ± 0.01 c | 8.96 ± 0.11 b | 9.16 ± 0.01 b |
| RAMULAB35 | 9.78 ± 0.15 c | 7.25 ± 0.31 a | 6.12 ± 0.02 b | 7.32 ± 0.01 d | 8.78 ± 0.04 b | 9.98 ± 0.03 c |
| RAMULAB36 | 9.84 ± 0.24 c | 8.25 ± 0.01 c | 6.19 ± 0.09 c | 6.41 ± 0.12 b | 8.21 ± 0.31 a | 9.71 ± 0.01 c |
| RAMULAB37 | 9.12 ± 0.02 b | 8.44 ± 0.07 c | 6.00 ± 0.13 a | 6.14 ± 0.02 a | 8.99 ± 0.12 b | 9.39 ± 0.04 b |
| RAMULAB38 | 9.65 ± 0.05 c | 7.89 ± 0.01 b | 6.13 ± 0.01 b | 7.05 ± 0.03 c | 8.85 ± 0.15 b | 9.95 ± 0.01 c |
| RAMULAB40 | 9.11 ± 0.18 b | 7.29 ± 0.10 a | 6.15 ± 0.16 c | 7.25 ± 0.01 d | 9.15 ± 0.23 c | 9.45 ± 0.03 b |
| RAMULAB41 | 8.81 ± 0.10 a | 7.23 ± 0.09 a | 6.11 ± 0.05 b | 6.91 ± 0.03 b | 8.81 ± 0.11 b | 9.01 ± 0.01 a |
| RAMULAB54 | 9.91 ± 0.01 d | 8.47 ± 0.03 c | 6.23 ± 0.01 d | 7.99 ± 0.11 e | 9.89 ± 0.01 d | 10.11 ± 0.71 d |
| Carbohydrates Fermentation | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Isolates | G | DX | LX | S | M | Mal | L | Gal | Ara | ST |
| RAMULAB33 | + | - | + | + | - | + | - | - | - | - |
| RAMULAB34 | + | - | + | + | + | + | + | + | - | - |
| RAMULAB35 | + | + | - | + | - | + | - | - | - | - |
| RAMULAB36 | + | - | + | + | + | + | + | + | - | - |
| RAMULAB37 | + | + | - | + | + | + | + | + | - | - |
| RAMULAB38 | + | + | - | + | + | + | + | + | - | - |
| RAMULAB40 | + | + | - | + | + | + | + | + | - | - |
| RAMULAB41 | + | + | - | + | + | + | + | + | - | - |
| RAMULAB54 | + | + | + | + | + | + | + | + | - | - |
| Cell Surface Hydrophobicity (%) * | |
|---|---|
| RAMULAB33 | 56.12 ± 0.09 a |
| RAMULAB34 | 70.54 ±0.05 d |
| RAMULAB35 | 57.81 ± 0.01 a |
| RAMULAB36 | 68.15 ± 0.09 c |
| RAMULAB37 | 65.33 ± 0.12 b |
| RAMULAB38 | 73.53 ± 0.04 d |
| RAMULAB40 | 73.82 ± 0.05 d |
| RAMULAB41 | 63.95 ± 0.04 b |
| RAMULAB54 | 75.23 ± 0.06 d |
| Isolates | HT-29 Adhesion (%) * |
|---|---|
| RAMULAB33 | 64.34 ± 0.11 b |
| RAMULAB34 | 69.16 ± 0.19 c |
| RAMULAB35 | 75.23 ± 0.08 d |
| RAMULAB36 | 85.12 ± 0.05 e |
| RAMULAB37 | 54.15 ± 0.01 a |
| RAMULAB38 | 65.12 ± 0.03 b |
| RAMULAB40 | 72.14 ± 0.06 d |
| RAMULAB41 | 81.15 ± 0.04 e |
| RAMULAB54 | 88.56 ± 0.03 f |
| Isolates | RAMULAB33 | RAMULAB34 | RAMULAB35 | RAMULAB36 | RAMULAB37 | RAMULAB38 | RAMULAB40 | RAMULAB41 | RAMULAB54 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Pathogens | K. pneumoniae | + | + | + | + | + | + | + | + | + |
| E. coli | ++ | ++ | ++ | ++ | +++ | ++ | ++ | ++ | +++ | |
| S. typhimurium | - | - | - | - | - | - | - | - | - | |
| S. aureus | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | +++ | |
| P. aeruginosa | ++ | +++ | ++ | +++ | +++ | +++ | +++ | +++ | +++ | |
| B. cereus | + | + | + | + | + | + | + | + | + | |
| M. luteus | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | |
| B. subtilis | + | + | + | + | + | + | + | + | + | |
| P. florescens | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | |
| K. aerogenes | - | - | - | - | - | - | - | - | - | |
| Sl. No. | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Antibiotic | Streptomycin (STR) | Vancomycin (V) | Tetracycline (TET) | Azithromycin (AZM) | Ampicillin (AMP) | Methicillin (MET) |
| The Inhibitory Zone (S/R mm) | (≥15/≤12) | (≥17/≤14) | (≥19/≤14) | (≥13/≤12) | (≥17/≤14) | (≥22/≤17) |
| RAMULAB33 | S | R | S | S | S | R |
| RAMULAB34 | S | R | S | S | S | R |
| RAMULAB35 | S | R | S | S | S | R |
| RAMULAB36 | S | R | S | S | S | R |
| RAMULAB37 | S | R | S | S | S | R |
| RAMULAB38 | S | R | S | S | S | R |
| RAMULAB40 | S | R | S | S | S | R |
| RAMULAB41 | S | R | S | S | S | R |
| RAMULAB54 | S | R | S | S | S | R |
| Organic Acids | RAMULAB54 (mg/mL) |
|---|---|
| Lactic acid | 7.09 ± 0.18 |
| Pyruvic acid | 0.59 ± 0.09 |
| Malonic acid | 3.62 ± 0.14 |
| Maleic acid | 0.04 ± 0.00 |
| Fumaric acid | 0.05 ± 0.00 |
| Succinic acid | 6.68 ± 0.48 |
| Malic acid | 5.68 ± 0.11 |
| Tartaric acid | 0.03 ± 0.00 |
| Shikimic acid | 0.33 ± 0.03 |
| Citric acid | 12.27 ± 0.39 |
| Hydroxycitric acid | 14.05 ± 0.58 |
| Compound | Activity | Pa | Pi |
|---|---|---|---|
| Citric acid | Antidiabetic | 0.648 | 0.009 |
| Fumaric acid | Antidiabetic | 0.512 | 0.021 |
| Hydroxycitric acid | Antidiabetic | 0.708 | 0.006 |
| Lactic acid | Antidiabetic | 0.680 | 0.007 |
| Malic acid | Antidiabetic | 0.639 | 0.009 |
| Malonic acid | Antidiabetic | 0.270 | 0.100 |
| Pyruvic acid | Antidiabetic symptomatic | 0.228 | 0.095 |
| Shikimic acid | Antidiabetic | 0.203 | 0.160 |
| Succinic acid | Antidiabetic | 0.440 | 0.034 |
| Tartaric acid | Antidiabetic | 0.719 | 0.005 |
| Acarbose | Antidiabetic | 0.693 | 0.007 |
| Compound | BA (kcal/mol) | TIN | THB | |||
|---|---|---|---|---|---|---|
| AG | AM | AG | AM | AG | AM | |
| Citric acid | −5.4 | −5.4 | 6 | 5 | 6 | 5 |
| Fumaric acid | −5.3 | −4.2 | 6 | 2 | 6 | 2 |
| Hydroxycitric acid | −5.8 | −5.5 | 8 | 4 | 8 | 4 |
| Lactic acid | −4.5 | −3.6 | 3 | 4 | 3 | 3 |
| Maleic acid | −5.3 | −4.1 | 5 | 4 | 5 | 4 |
| Malic acid | −5.2 | −4.4 | 4 | 3 | 4 | 3 |
| Malonic acid | −4.8 | −3.8 | 5 | 3 | 5 | 3 |
| Pyruvic acid | −4.5 | −3.3 | 5 | 2 | 4 | 2 |
| Shikimic acid | −5.3 | −5.3 | 5 | 3 | 5 | 3 |
| Succinic acid | −5.1 | −4.0 | 3 | 2 | 3 | 2 |
| Tartaric acid | −5.7 | −5.5 | 7 | 4 | 7 | 4 |
| Acarbose | −5.6 | −5.4 | 7 | 4 | 6 | 4 |
| MD Trajectory Values | Apo-Protein | Protein-Acarbose Complex | Protein- Hydroxycitric Acid Complex |
|---|---|---|---|
| RMSD (nm) | 0.30–0.40 | 0.25–0.32 | 0.20–0.25 |
| Rg (nm) | 3.10–3.14 | 2.39–2.40 | 2.39–2.40 |
| SASA (nm2) | 350–370 | 240–250 | 240–250 |
| Ligand H-bonds | - | 7 | 8 |
| MD Trajectory Values | Apo-Protein | Protein-Acarbose Complex | Protein- Hydroxycitric Acid Complex |
|---|---|---|---|
| RMSD (nm) | 0.20–0.30 | 0.25–0.35 | 0.20–0.25 |
| Rg (nm) | 2.31 | 2.31 | 2.31 |
| SASA (nm2) | 190–200 | 190–205 | 190–200 |
| Ligand H-bonds | - | 7 | 10 |
| Protein-Ligand Complexes | Types of Binding Free Energies | ||||
|---|---|---|---|---|---|
| VDWE (kj/mol) | EE (kj/mol) | PSE (kj/mol) | SASAE (kj/mol) | BE (kj/mol) | |
| AG-hydroxycitric acid | −220.118 | −9.313 | 96.102 | −28.166 | −189.1022 |
| AG-acarbose | −134.192 | −4.813 | 62.125 | −9.310 | −90.102 |
| AM-hydroxycitric acid | −218.568 | −29.891 | 62.172 | −21.886 | −180.194 |
| AM-acarbose | −130.161 | −2.106 | 39.340 | −9.564 | −87.109 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumari V. B., C.; Huligere, S.S.; Alotaibi, G.; Al Mouslem, A.K.; Bahauddin, A.A.; Shivanandappa, T.B.; Ramu, R. Antidiabetic Activity of Potential Probiotics Limosilactobacillus spp., Levilactobacillus spp., and Lacticaseibacillus spp. Isolated from Fermented Sugarcane Juice: A Comprehensive In Vitro and In Silico Study. Nutrients 2023, 15, 1882. https://doi.org/10.3390/nu15081882
Kumari V. B. C, Huligere SS, Alotaibi G, Al Mouslem AK, Bahauddin AA, Shivanandappa TB, Ramu R. Antidiabetic Activity of Potential Probiotics Limosilactobacillus spp., Levilactobacillus spp., and Lacticaseibacillus spp. Isolated from Fermented Sugarcane Juice: A Comprehensive In Vitro and In Silico Study. Nutrients. 2023; 15(8):1882. https://doi.org/10.3390/nu15081882
Chicago/Turabian StyleKumari V. B., Chandana, Sujay S. Huligere, Ghallab Alotaibi, Abdulaziz K. Al Mouslem, Ammar Abdulraheem Bahauddin, Thippeswamy Boreddy Shivanandappa, and Ramith Ramu. 2023. "Antidiabetic Activity of Potential Probiotics Limosilactobacillus spp., Levilactobacillus spp., and Lacticaseibacillus spp. Isolated from Fermented Sugarcane Juice: A Comprehensive In Vitro and In Silico Study" Nutrients 15, no. 8: 1882. https://doi.org/10.3390/nu15081882
APA StyleKumari V. B., C., Huligere, S. S., Alotaibi, G., Al Mouslem, A. K., Bahauddin, A. A., Shivanandappa, T. B., & Ramu, R. (2023). Antidiabetic Activity of Potential Probiotics Limosilactobacillus spp., Levilactobacillus spp., and Lacticaseibacillus spp. Isolated from Fermented Sugarcane Juice: A Comprehensive In Vitro and In Silico Study. Nutrients, 15(8), 1882. https://doi.org/10.3390/nu15081882