
What the Lactate Shuttle Means for Sports Nutrition
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Exercise and Cell Work
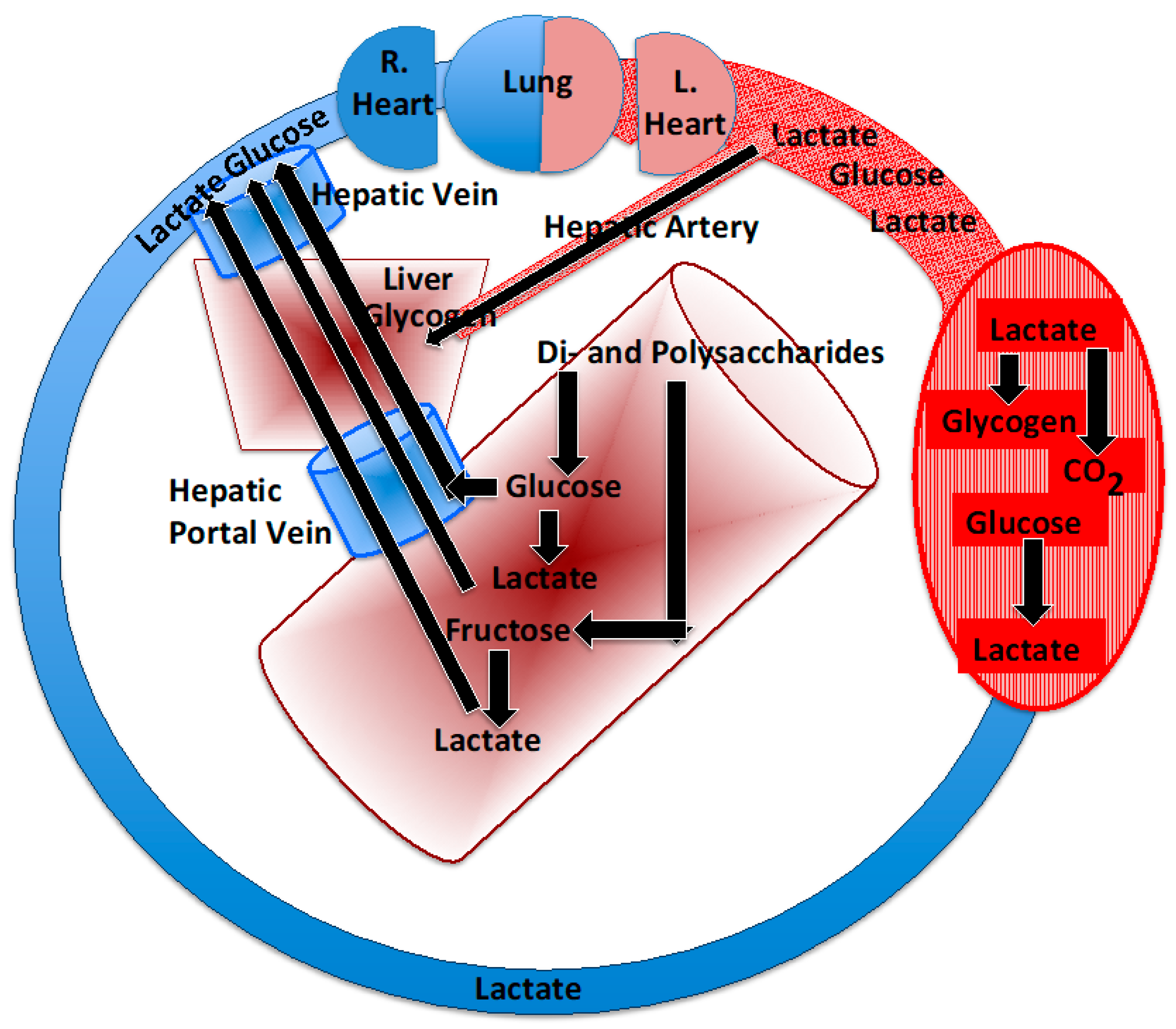
2.2. The Postprandial Lactate Shuttle
2.3. Role of the Gut in Lactate Shuttling
2.4. Brainless Sports Nutrition
2.5. Sour News
2.6. Why Lactate and Not Other Bioenergetic Substances?
2.7. Generally Accepted as Safe
3. Discussion
4. Conclusions
5. Patents
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brooks, G.A. Glycolytic end product and oxidative substrate during sustained exercise in mammals—The “actate shuttle”. In Comparative Physiology and Biochemistry—Current Topics and Trends, Volume A, Respiration—Metabolism—Circulation; Springer: Berlin/Heidelberg, Germany, 1985; pp. 208–218. [Google Scholar]
- Brooks, G.A. Lactate production under fully aerobic conditions: The lactate shuttle during rest and exercise. Fed. Proc. 1986, 45, 2924–2929. [Google Scholar] [PubMed]
- Brooks, G.A. Lactate shuttles in nature. Biochem. Soc. Trans. 2002, 30, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef]
- Brooks, G.A. Lactate as a fulcrum of metabolism. Redox Biol. 2020, 35, 101454. [Google Scholar] [CrossRef]
- Brooks, G.A. Role of the Heart in Lactate Shuttling. Front. Nutr. 2021, 8, 663560. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A.; Arevalo, J.A.; Osmond, A.D.; Leija, R.G.; Curl, C.C.; Tovar, A.P. Lactate in contemporary biology: A phoenix risen. J. Physiol. 2022, 600, 1229–1251. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. Cell-cell and intracellular lactate shuttles. J Physiol 2009, 587, 5591–5600. [Google Scholar] [CrossRef]
- Hui, S.; Ghergurovich, J.M.; Morscher, R.J.; Jang, C.; Teng, X.; Lu, W.; Esparza, L.A.; Reya, T.; Le, Z.; Yanxiang Guo, J.; et al. Glucose feeds the TCA cycle via circulating lactate. Nature 2017, 551, 115–118. [Google Scholar] [CrossRef]
- Bartman, C.R.; TeSlaa, T.; Rabinowitz, J.D. Quantitative flux analysis in mammals. Nat. Metab. 2021, 3, 896–908. [Google Scholar] [CrossRef]
- Chen, Y.J.; Mahieu, N.G.; Huang, X.; Singh, M.; Crawford, P.A.; Johnson, S.L.; Gross, R.W.; Schaefer, J.; Patti, G.J. Lactate metabolism is associated with mammalian mitochondria. Nat. Chem. Biol. 2016, 12, 937–943. [Google Scholar] [CrossRef]
- Schurr, A.; Passarella, S. Aerobic Glycolysis: A DeOxymoron of (Neuro)Biology. Metabolites 2022, 12, 72. [Google Scholar] [CrossRef]
- Brooks, G.A.; Osmond, A.D.; Arevalo, J.A.; Duong, J.J.; Curl, C.C.; Moreno-Santillan, D.D.; Leija, R.G. Lactate as a myokine and exerkine: Drivers and signals of physiology and metabolism. J. Appl. Physiol. 2023, 134, 529–548. [Google Scholar] [CrossRef] [PubMed]
- Bergman, B.C.; Wolfel, E.E.; Butterfield, G.E.; Lopaschuk, G.D.; Casazza, G.A.; Horning, M.A.; Brooks, G.A. Active muscle and whole body lactate kinetics after endurance training in men. J. Appl. Physiol. 1999, 87, 1684–1696. [Google Scholar] [CrossRef]
- Bergman, B.C.; Tsvetkova, T.; Lowes, B.; Wolfel, E.E. Myocardial glucose and lactate metabolism during rest and atrial pacing in humans. J. Physiol. 2009, 587, 2087–2099. [Google Scholar] [CrossRef]
- Gertz, E.W.; Wisneski, J.A.; Stanley, W.C.; Neese, R.A. Myocardial substrate utilization during exercise in humans. Dual carbon-labeled carbohydrate isotope experiments. J. Clin. Invest. 1988, 82, 2017–2025. [Google Scholar] [CrossRef]
- Glenn, T.C.; Martin, N.A.; Horning, M.A.; McArthur, D.L.; Hovda, D.; Vespa, P.M.; Brooks, G.A. Lactate: Brain Fuel in Human Traumatic Brain Injury. A Comparison to Normal Healthy Control Subjects. J. Neurotrauma 2015, 32, 820–832. [Google Scholar] [CrossRef]
- Bergman, B.C.; Horning, M.A.; Casazza, G.A.; Wolfel, E.E.; Butterfield, G.E.; Brooks, G.A. Endurance training increases gluconeogenesis during rest and exercise in men. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E244–E251. [Google Scholar] [CrossRef]
- Stanley, W.C.; Gertz, E.W.; Wisneski, J.A.; Morris, D.L.; Neese, R.A.; Brooks, G.A. Systemic lactate kinetics during graded exercise in man. Am. J. Physiol. 1985, 249, E595–E602. [Google Scholar] [CrossRef] [PubMed]
- Gerich, J.E.; Meyer, C.; Woerle, H.J.; Stumvoll, M. Renal gluconeogenesis: Its importance in human glucose homeostasis. Diabetes Care 2001, 24, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.; Stumvoll, M.; Dostou, J.; Welle, S.; Haymond, M.; Gerich, J. Renal substrate exchange and gluconeogenesis in normal postabsorptive humans. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E428–E434. [Google Scholar] [CrossRef]
- Brooks, G.A.; Donovan, C.M. Effect of endurance training on glucose kinetics during exercise. Am. J. Physiol. 1983, 244, E505–E512. [Google Scholar] [CrossRef] [PubMed]
- Donovan, C.M.; Brooks, G.A. Endurance training affects lactate clearance, not lactate production. Am. J. Physiol. 1983, 244, E83–E92. [Google Scholar] [CrossRef]
- Mazzeo, R.S.; Brooks, G.A.; Budinger, T.F.; Schoeller, D.A. Pulse injection, 13C tracer studies of lactate metabolism in humans during rest and two levels of exercise. Biomed. Mass Spectrom. 1982, 9, 310–314. [Google Scholar] [CrossRef]
- Mazzeo, R.S.; Brooks, G.A.; Schoeller, D.A.; Budinger, T.F. Disposal of blood [1-13C]lactate in humans during rest and exercise. J. Appl. Physiol. 1986, 60, 232–241. [Google Scholar] [CrossRef]
- Stanley, W.C.; Gertz, E.W.; Wisneski, J.A.; Neese, R.A.; Morris, D.L.; Brooks, G.A. Lactate extraction during net lactate release in legs of humans during exercise. J. Appl. Physiol. 1986, 60, 1116–1120. [Google Scholar] [CrossRef]
- Gertz, E.W.; Wisneski, J.A.; Neese, R.; Bristow, J.D.; Searle, G.L.; Hanlon, J.T. Myocardial lactate metabolism: Evidence of lactate release during net chemical extraction in man. Circulation 1981, 63, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, L.; Magistretti, P.J. Glutamate uptake into astrocytes stimulates aerobic glycolysis: A mechanism coupling neuronal activity to glucose utilization. Proc. Natl. Acad. Sci. USA 1994, 91, 10625–10629. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Pellerin, L.; Rothman, D.L.; Shulman, R.G. Energy on demand. Science 1999, 283, 496–497. [Google Scholar] [CrossRef]
- Foster, D.W. Banting lecture 1984. From glycogen to ketones--and back. Diabetes 1984, 33, 1188–1199. [Google Scholar] [CrossRef] [PubMed]
- Woerle, H.J.; Meyer, C.; Dostou, J.M.; Gosmanov, N.R.; Islam, N.; Popa, E.; Wittlin, S.D.; Welle, S.L.; Gerich, J.E. Pathways for glucose disposal after meal ingestion in humans. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E716–E725. [Google Scholar] [CrossRef]
- Stender, S.; Zaha, V.G.; Malloy, C.R.; Sudderth, J.; DeBerardinis, R.J.; Park, J.M. Assessment of Rapid Hepatic Glycogen Synthesis in Humans Using Dynamic (13)C Magnetic Resonance Spectroscopy. Hepatol. Commun. 2020, 4, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Barnard, R.J.; Edgerton, V.R.; Furukawa, T.; Peter, J.B. Histochemical, biochemical, and contractile properties of red, white, and intermediate fibers. Am. J. Physiol. 1971, 220, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Barnard, R.J.; Edgerton, V.R.; Peter, J.B. Effect of exercise on skeletal muscle. I. Biochemical and histochemical properties. J. Appl. Physiol. 1970, 28, 762–766. [Google Scholar] [CrossRef] [PubMed]
- James, D.E.; Kraegen, E.W.; Chisholm, D.J. Effects of exercise training on in vivo insulin action in individual tissues of the rat. J. Clin. Investig. 1985, 76, 657–666. [Google Scholar] [CrossRef] [PubMed]
- James, D.E.; Zorzano, A.; Boni-Schnetzler, M.; Nemenoff, R.A.; Powers, A.; Pilch, P.F.; Ruderman, N.B. Intrinsic differences of insulin receptor kinase activity in red and white muscle. J. Biol. Chem. 1986, 261, 14939–14944. [Google Scholar] [CrossRef]
- Smadja, C.; Morin, J.; Ferre, P.; Girard, J. Metabolic fate of a gastric glucose load in unrestrained rats bearing a portal vein catheter. Am. J. Physiol. 1988, 254, E407–E413. [Google Scholar] [CrossRef] [PubMed]
- Lecoultre, V.; Benoit, R.; Carrel, G.; Schutz, Y.; Millet, G.P.; Tappy, L.; Schneiter, P. Fructose and glucose co-ingestion during prolonged exercise increases lactate and glucose fluxes and oxidation compared with an equimolar intake of glucose. Am. J. Clin. Nutr. 2010, 92, 1071–1079. [Google Scholar] [CrossRef]
- Theytaz, F.; de Giorgi, S.; Hodson, L.; Stefanoni, N.; Rey, V.; Schneiter, P.; Giusti, V.; Tappy, L. Metabolic fate of fructose ingested with and without glucose in a mixed meal. Nutrients 2014, 6, 2632–2649. [Google Scholar] [CrossRef]
- Brooks, G.A.; Curl, C.C.; Leija, R.G.; Osmond, A.D.; Duong, J.J.; Arevalo, J.A. Tracing the lactate shuttle to the mitochondrial reticulum. Exp. Mol. Med. 2022, 54, 1332–1347. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojarvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916 e917. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar] [CrossRef] [PubMed]
- Scheiman, J.; Luber, J.M.; Chavkin, T.A.; MacDonald, T.; Tung, A.; Pham, L.D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T.; et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Coady, M.J.; Chang, M.H.; Charron, F.M.; Plata, C.; Wallendorff, B.; Sah, J.F.; Markowitz, S.D.; Romero, M.F.; Lapointe, J.Y. The human tumour suppressor gene SLC5A8 expresses a Na+-monocarboxylate cotransporter. J. Physiol. 2004, 557, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Teramae, H.; Yoshikawa, T.; Inoue, R.; Ushida, K.; Takebe, K.; Nio-Kobayashi, J.; Iwanaga, T. The cellular expression of SMCT2 and its comparison with other transporters for monocarboxylates in the mouse digestive tract. Biomed.Res. 2010, 31, 239–249. [Google Scholar] [CrossRef]
- Brooks, G.A.; Dubouchaud, H.; Brown, M.; Sicurello, J.P.; Butz, C.E. Role of mitochondrial lactate dehydrogenase and lactate oxidation in the intracellular lactate shuttle. Proc. Natl. Acad. Sci. USA 1999, 96, 1129–1134. [Google Scholar] [CrossRef]
- Brooks, G.A.; Fahey, T.D. Exercise Physiology: Human Bioenergetics and Its Applications, 1st ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1984. [Google Scholar]
- Brooks, G.A.; Fahey, T.D.; Baldwin, K.M. Exercise Physiology: Human Bioenergetics and It’s Applications, 4th ed.; McGraw-Hill: Mountain View, CA, USA, 2004; ISBN 978-0072556421. [Google Scholar]
- Schurr, A. Lactate: The ultimate cerebral oxidative energy substrate? J. Cereb. Blood Flow Metab. 2006, 26, 142–152. [Google Scholar] [CrossRef]
- Thurn, J.R.; Pierpont, G.L.; Ludvigsen, C.W.; Eckfeldt, J.H. D-lactate encephalopathy. Am. J. Med. 1985, 79, 717–721. [Google Scholar] [CrossRef]
- Chan, L.; Slater, J.; Hasbargen, J.; Herndon, D.N.; Veech, R.L.; Wolf, S. Neurocardiac toxicity of racemic D,L-lactate fluids. Integr. Physiol. Behav. Sci. 1994, 29, 383–394. [Google Scholar] [CrossRef]
- Nalos, M.; Leverve, X.; Huang, S.; Weisbrodt, L.; Parkin, R.; Seppelt, I.; Ting, I.; McLean, A. Half-molar sodium lactate infusion improves cardiac performance in acute heart failure: A pilot randomised controlled clinical trial. Crit. Care 2014, 18, R48. [Google Scholar] [CrossRef]
- Bouzat, P.; Sala, N.; Suys, T.; Zerlauth, J.B.; Marques-Vidal, P.; Feihl, F.; Bloch, J.; Messerer, M.; Levivier, M.; Meuli, R.; et al. Cerebral metabolic effects of exogenous lactate supplementation on the injured human brain. Intensive Care Med. 2014, 40, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Quintard, H.; Patet, C.; Zerlauth, J.B.; Suys, T.; Bouzat, P.; Pellerin, L.; Meuli, R.; Magistretti, P.; Oddo, M. Improvement of neuroenergetics by hypertonic lactate therapy in patients with traumatic brain injury is dependent on baseline cerebral lactate/pyruvate ratio. J. Neurotrauma 2015, 33, 681–687. [Google Scholar] [CrossRef]
- Brooks, G.A.; Martin, N.A. Cerebral metabolism following traumatic brain injury: New discoveries with implications for treatment. Front. Neurosci. 2014, 8, 408. [Google Scholar] [CrossRef]
- Garcia-Alvarez, M.; Marik, P.; Bellomo, R. Sepsis-associated hyperlactatemia. Crit. Care 2014, 18, 503. [Google Scholar] [CrossRef]
- Garcia-Alvarez, M.; Marik, P.; Bellomo, R. Stress hyperlactataemia: Present understanding and controversy. Lancet Diabetes Endocrinol. 2014, 2, 339–347. [Google Scholar] [CrossRef]
- See, E.J.; Bellomo, R. The importance of applying physiological principles of hyperlactataemia to the study of human disease. J. Physiol. 2021, 599, 1933. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.F.; Fattor, J.A.; Jacobs, K.A.; Horning, M.A.; Navazio, F.; Lindinger, M.I.; Brooks, G.A. Lactate and glucose interactions during rest and exercise in men: Effect of exogenous lactate infusion. J. Physiol. 2002, 544, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.F.; Fattor, J.A.; Jacobs, K.A.; Horning, M.A.; Suh, S.H.; Navazio, F.; Brooks, G.A. Metabolic and cardiorespiratory responses to “the lactate clamp”. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E889–E898. [Google Scholar] [CrossRef]
- Miller, B.F.; Lindinger, M.I.; Fattor, J.A.; Jacobs, K.A.; Leblanc, P.J.; Duong, M.; Heigenhauser, G.J.; Brooks, G.A. Hematological and acid-base changes in men during prolonged exercise with and without sodium-lactate infusion. J. Appl. Physiol. 2005, 98, 856–865. [Google Scholar] [CrossRef]
- Messonnier, A.L.; Emhoff, C.W.; Fattor, J.A.; Horning, M.A.; Carlson, T.J.; Brooks, G.A. Lactate kinetics at the lactate threshold in trained and untrained men. J. Appl. Physiol. 2013, 114, 1593–1602. [Google Scholar] [CrossRef]
- Sumegi, B.; Podanyi, B.; Forgo, P.; Kover, K.E. Metabolism of [3-13C]pyruvate and [3-13C]propionate in normal and ischaemic rat heart in vivo: 1H- and 13C-NMR studies. Biochem. J. 1995, 312, 75–81. [Google Scholar] [CrossRef]
- Johnson, M.L.; Hussien, R.; Horning, M.A.; Brooks, G.A. Transpulmonary pyruvate kinetics. Am. J. Physiol. Regul. Integr Comp. Physiol. 2011, 301, R769–R774. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.C.; Horning, M.A.; Wallis, G.A.; Brooks, G.A. Pyruvate metabolism in working human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E366. [Google Scholar] [CrossRef]
- Osuna-Prieto, F.J.; Martinez-Tellez, B.; Ortiz-Alvarez, L.; Di, X.; Jurado-Fasoli, L.; Xu, H.; Ceperuelo-Mallafre, V.; Nunez-Roa, C.; Kohler, I.; Segura-Carretero, A.; et al. Elevated plasma succinate levels are linked to higher cardiovascular disease risk factors in young adults. Cardiovasc. Diabetol. 2021, 20, 151. [Google Scholar] [CrossRef] [PubMed]
- Stegink, L.D.; Filer, L.J., Jr.; Baker, G.L.; Bell, E.F. Plasma glutamate concentrations in 1-year-old infants and adults ingesting monosodium L-glutamate in consomme. Pediatr. Res. 1986, 20, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A.; Mercier, J. Balance of carbohydrate and lipid utilization during exercise: The “crossover” concept. J. Appl. Physiol. 1994, 76, 2253–2261. [Google Scholar] [CrossRef]
- Saddik, M.; Gamble, J.; Witters, L.A.; Lopaschuk, G.D. Acetyl-CoA carboxylase regulation of fatty acid oxidation in the heart. J. Biol. Chem. 1993, 268, 25836–25845. [Google Scholar] [CrossRef] [PubMed]
- San-Millan, I.; Sparagna, G.C.; Chapman, H.L.; Warkins, V.L.; Chatfield, K.C.; Shuff, S.R.; Martinez, J.L.; Brooks, G.A. Chronic Lactate Exposure Decreases Mitochondrial Function by Inhibition of Fatty Acid Uptake and Cardiolipin Alterations in Neonatal Rat Cardiomyocytes. Front. Nutr. 2022, 9, 809485. [Google Scholar] [CrossRef]
- Chiolero, R.; Mavrocordatos, P.; Burnier, P.; Cayeux, M.C.; Schindler, C.; Jequier, E.; Tappy, L. Effects of infused sodium acetate, sodium lactate, and sodium beta-hydroxybutyrate on energy expenditure and substrate oxidation rates in lean humans. Am. J. Clin. Nutr. 1993, 58, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.; Tchabanenko, K.; Pawlosky, R.; Carter, E.; Todd King, M.; Musa-Veloso, K.; Ho, M.; Roberts, A.; Robertson, J.; Vanitallie, T.B.; et al. Kinetics, safety and tolerability of (R)-3-hydroxybutyl (R)-3-hydroxybutyrate in healthy adult subjects. Regul. Toxicol. Pharmacol. 2012, 63, 401–408. [Google Scholar] [CrossRef]
- Roth, D.A.; Brooks, G.A. Lactate transport is mediated by a membrane-bound carrier in rat skeletal muscle sarcolemmal vesicles. Arch. Biochem. Biophys. 1990, 279, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Roth, D.A.; Brooks, G.A. Lactate and pyruvate transport is dominated by a pH gradient-sensitive carrier in rat skeletal muscle sarcolemmal vesicles. Arch. Biochem. Biophys. 1990, 279, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, J.D.; Enerback, S. Lactate: The ugly duckling of energy metabolism. Nat. Metab. 2020, 2, 566–571. [Google Scholar] [CrossRef]
- Brooks, G.A.; Osmond, A.D.; Arevalo, J.A.; Curl, C.C.; Duong, J.J.; Horning, M.A.; Moreno Santillan, D.D.; Leija, R.G. Lactate as a major myokine and exerkine. Nat. Rev. Endocrinol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Tsukamoto, H.; Takenaka, S.; Olesen, N.D.; Petersen, L.G.; Sorensen, H.; Nielsen, H.B.; Secher, N.H.; Ogoh, S. Maintained exercise-enhanced brain executive function related to cerebral lactate metabolism in men. FASEB J. 2018, 32, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. Method and Composition For Energy Source Supplementation during Exercise and Recovery. U.S. Patent 5420107A, 25 July 1994. [Google Scholar]
- Brooks, G.A. Lactate Thiolester for Cardiac Energy Resuscitation and Prevention of Reperfusion Injury and Use as an Energy Supplement during Exercise and Recovery. U.S. Patent 6,482,853 B1, 19 November 2002. [Google Scholar]
- Brooks, G.A. Glycerol-Lactate Esters for Use as an Energy Supplement during Exercise and Recovery. U.S. Patent 6,743,821 B2, 1 June 2004. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brooks, G.A. What the Lactate Shuttle Means for Sports Nutrition. Nutrients 2023, 15, 2178. https://doi.org/10.3390/nu15092178
Brooks GA. What the Lactate Shuttle Means for Sports Nutrition. Nutrients. 2023; 15(9):2178. https://doi.org/10.3390/nu15092178
Chicago/Turabian StyleBrooks, George A. 2023. "What the Lactate Shuttle Means for Sports Nutrition" Nutrients 15, no. 9: 2178. https://doi.org/10.3390/nu15092178
APA StyleBrooks, G. A. (2023). What the Lactate Shuttle Means for Sports Nutrition. Nutrients, 15(9), 2178. https://doi.org/10.3390/nu15092178