Current Trends on the Involvement of Zinc, Copper, and Selenium in the Process of Hepatocarcinogenesis

Abstract
:1. Introduction
2. Zinc
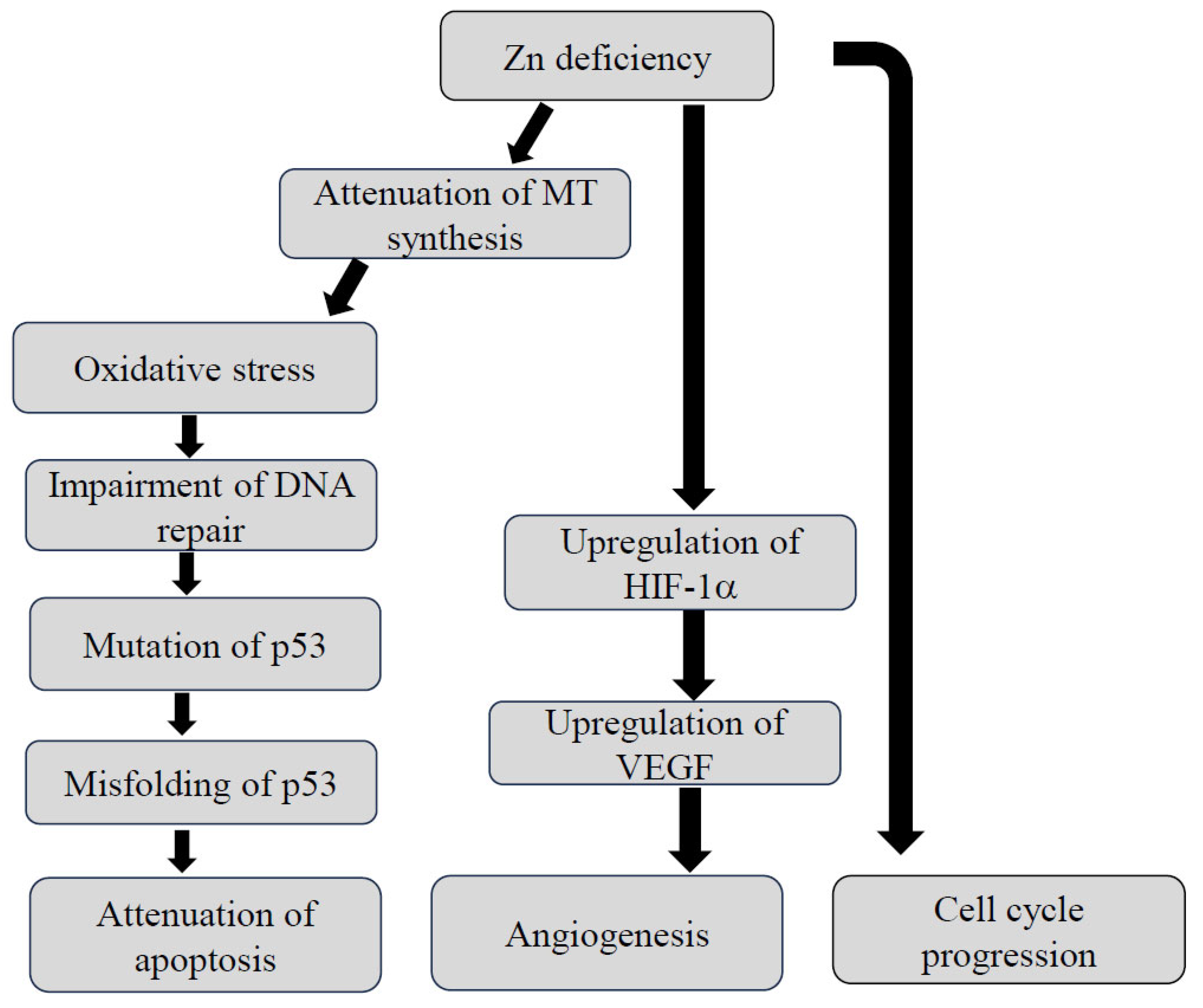
2.1. The Interaction between Zn Status and Carcinogenesis
2.2. Zn Status in Patients with HCC
2.3. The Effects of Zn Supplementation in HCC Cell Lines or Experimental HCC Models
2.4. The Preventive Effects of Zinc Supplementation on the Progression of HCC in the Clinical Trials
3. Copper
3.1. Cu Homeostasis
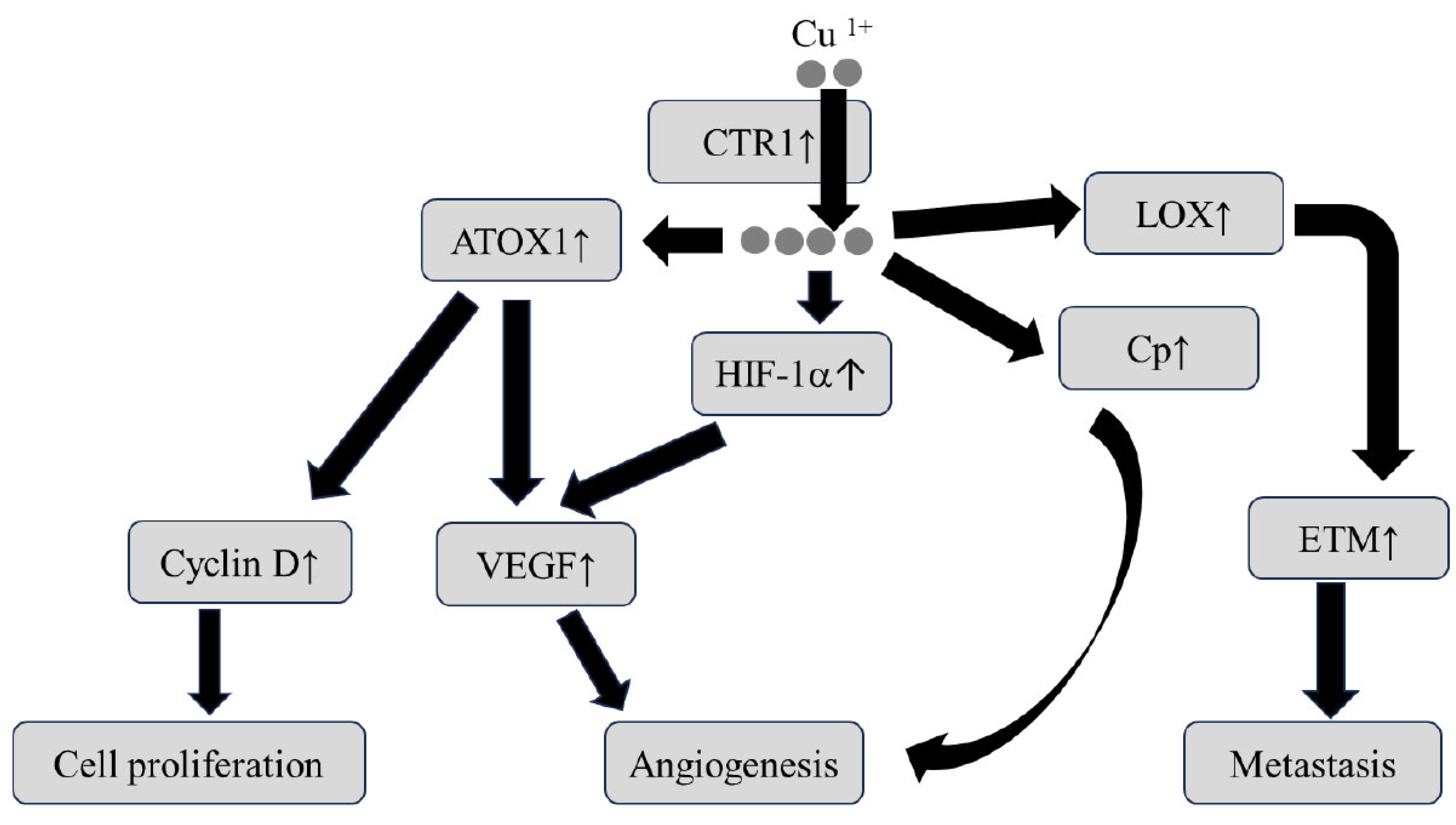
3.2. The Relationship between Cu Status and Carcinogenesis
3.3. The Relationship between Cu Metabolism and Chemoresistance in Cancer
3.4. Cu Status in Patients with HCC
3.5. Therapeutic Strategies Which Target Cu in HCC Cell Lines or Experimental HCC Models
3.6. Therapeutic Strategies Which Target Cu in Patients with HCC
4. Selenium
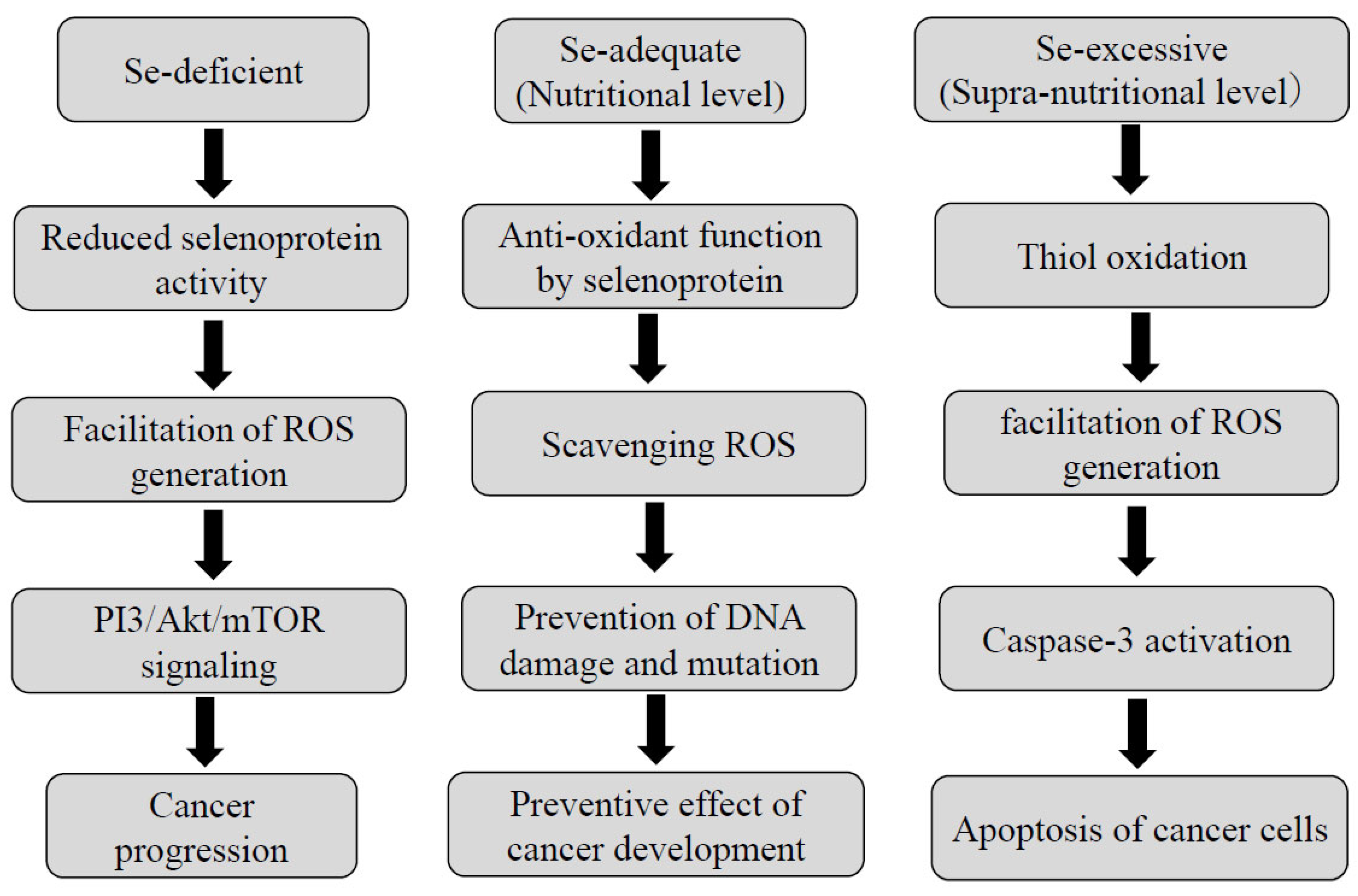
4.1. The Relationship between Se Status and Carcinogenesis
4.2. Interaction of Se with Other Essential Trace Elements
4.3. Se and Chemosensitivity in Cancer
4.4. Se Status in Patients with HCC
4.5. Therapeutic Strategies Which Target Se in HCC Cell Lines or Experimental HCC Models
4.6. Therapeutic Strategies Which Targeting Se in Clinical Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ATOX1 | Antioxidant protein 1 |
| ATP7A/B | Copper-transporting ATPase 1/2 |
| CLD | Chronic liver disease |
| COX17 | Cytochrome C oxidase 17 |
| CTL | Cytotoxic T lymphocytes |
| CTR1 | Copper transporter receptor 1 |
| Cu | Copper |
| DSF | Disulfiram |
| EMT | Epithelial mesenchymal transition |
| GPx | Glutathione peroxidase |
| HBV | Hepatitis B virus |
| HCC | Hepatocellular carcinoma |
| HCV | Hepatitis C virus |
| HIF-1α | Hypoxia-inducible factor-1α |
| LOX | Lysyl oxidase |
| LOXL | Lysyl oxidase-like |
| MAPK | Mitogen-activated protein kinase |
| MT | Methallothioneine |
| NK | Natural killer |
| PD-L1 | Programed death-ligand 1 |
| ROS | Reactive oxygen species |
| Se | Selenium |
| SeNP | Selenium nanoparticle |
| SVR | Sustained viral response |
| TrxR | Thioredoxin reductase |
| TTM | Tetrathiomolybdate |
| VEGF | Vascular endothelial growth factor |
| ZIP | Zrt- and Irk-like protein |
| Zn | Zinc |
References
- Yang, J.D.; Hainaut, P.; Gores, G.J.; Amadou, A.; Plymoth, A.; Roberts, L.R. A global view of hepatocellular carcinoma: Trends, risk, prevention and management. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 589–604. [Google Scholar] [CrossRef]
- Villanueva, A. Hepatocellular carcinoma. N. Engl. J. Med. 2019, 380, 1450–1462. [Google Scholar] [CrossRef]
- McGlynn, K.A.; Petrick, J.L.; El-Serag, H.B. Epidemiology of hepatocellular carcinoma. Hepatology 2021, 73 (Suppl. S1), 4–13. [Google Scholar] [CrossRef]
- Kulik, L.; El-Serag, H.B. Epidemiology and management of hepatocellular carcinoma. Gastroenterology 2019, 156, 477–491. [Google Scholar] [CrossRef]
- Shah, P.A.; Patil, R.; Harrison, S.A. NAFLD-related hepatocellular carcinoma: The growing challenge. Hepatology 2023, 77, 323–338. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver. EASL Clinical practice guidelines: Management of hepatocellular carcinoma. J. Hepatol. 2018, 69, 182–236. [Google Scholar] [CrossRef] [PubMed]
- Marrero, J.A.; Kulik, L.M.; Sirlin, C.B.; Zhu, A.X.; Finn, R.S.; Abecassis, M.M.; Roberts, L.R.; Heimbach, J.K. Diagnosis, staging, and management of hepatocellular carcinoma: 2018 practice guideline by the American Association for the Study of the Liver Diseases. Hepatology 2018, 68, 723–750. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Takemura, N.; Yamashita, T.; Watadani, T.; Kaibori, M.; Kubo, S.; Shimada, M.; Nagano, H.; Hatano, E.; Aikata, H.; et al. Clinical practice guideline for hepatocellular carcinoma: The Japan Society of Hepatology 2021 version (5th JSH-HCC guidelines). Hepatol. Res. 2023, 53, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Siddique, A.; Kowdley, K.V. Insulin resistance and other metabolic risk factors in the pathogenesis of hepatocellular carcinoma. Clin. Liver Dis. 2011, 15, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Chettouh, H.; Lequoy, M.; Fartoux, L.; Vigouroux, C.; Desbois-Mouthon, C. Hyperinsulinemia and insulin signaling in the pathogenesis and the clinical course of hepatocellular carcinoma. Liver Int. 2015, 35, 2203–2217. [Google Scholar] [CrossRef] [PubMed]
- Mandair, D.S.; Rossi, R.E.; Pericleous, M.; Whyand, T.; Caplin, M. The impact of diet and nutrition in the prevention and progression of hepatocellular carcinoma. Expert. Rev. Gastroenterol. Hepatol. 2014, 8, 369–382. [Google Scholar] [CrossRef]
- Koumbi, L. Dietary factors can protect against liver cancer development. World J. Hepatol. 2017, 9, 119–125. [Google Scholar] [CrossRef]
- Ruiz-Margáin, A.; Román-Calleja, B.M.; Moreno-Guillén, P.; González-Regueiro, J.A.; Kúsulas-Delint, D.; Campos-Murguía, A.; Flores-García, N.C.; Macías-Rodríguez, R.U. Nutrition therapy for hepatocellular carcinoma. World J. Oncol. 2021, 13, 1440–1452. [Google Scholar]
- Yang, W.-S.; Zeng, X.-F.; Liu, Z.-N.; Zhao, Q.-H.; Tan, Y.-T.; Gao, J.; Li, H.-L.; Xiang, Y.-B. Diet and liver cancer risk: A narrative review of epidemiological evidence. Br. J. Nutr. 2020, 24, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Juloski, J.T.; Rakic, A.; Ćuk, V.V.; Ćuk, V.M.; Stefanović, S.; Nikolić, D.; Janković, S.; Trbovich, A.M.; De Luka, S.R. Colorectal cancer and trace elements alteration. J. Trace Elem. Med. Biol. 2020, 59, 126451. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Ali, I. Dietary trace element intake and risk of breast cancer: A mini review. Biol. Trace Elem. Res. 2022, 200, 4936–4948. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Tang, Z.; Li, J.; Jiang, J. Esophagus cancer and essential trace elements. Front. Public Health 2022, 10, 1038153. [Google Scholar] [CrossRef] [PubMed]
- Gurusamy, K. Trace element concentration in primary liver cancers-a systemic review. Biol. Trace Elem. Res. 2007, 118, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Res. 1993, 73, 79–118. [Google Scholar] [CrossRef]
- Prasad, A.S. Zinc is an antioxidant and anti-inflammatory agent: Its role in human health. Front. Nutr. 2014, 1, 14. [Google Scholar] [CrossRef]
- Lubna, S.; Ahmad, R. Clinical and biochemical understanding of zinc interaction during liver diseases: A paradigm shift. J. Trace Elem. Med. Biol. 2023, 77, 127130. [Google Scholar] [CrossRef]
- Vasak, M. Advances in metallothionein structure and functions. J. Trace Elem. Med. Biol. 2005, 19, 13–17. [Google Scholar] [CrossRef]
- Bell, S.G.; Valee, B.L. The metallothionein/thionein system: An oxidoreductive metabolic zinc link. Chemobiochem 2009, 10, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Krizkova, S.; Ryvolova, M.; Hrabeta, J.; Adam, V.; Stiborova, M.; Eckschlager, T.; Kizek, R. Metallothioneins and zinc in cancer diagnosis and therapy. Drug Metab. Rev. 2012, 44, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Moleirinho, A.; Carneiro, J.; Matthiesen, R.; Silva, R.M.; Amorim, A.; Azevedo, L. Gains, losses and changes of function after gene duplication: Study of the metallothionein family. PLoS ONE 2011, 6, e18487. [Google Scholar] [CrossRef] [PubMed]
- Si, M.; Lang, J. The roles of metallothioneins in carcinogenesis. J. Hematol. Oncol. 2018, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.B.; Costello, L.C. The important roles of apoptotic effects of zinc in the development of cancers. J. Cell Biochem. 2009, 106, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Ho, E. Zinc deficiency, DNA damage and cancer risk. J. Nutr. Biochem. 2004, 15, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Chasapis, C.T.; Loutsidou, A.C.; Spiliopoulou, C.A.; Stefanidou, M.E. Zinc and human health: An update. Arch. Toxicol. 2012, 86, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Tian, Y.; Zhang, H.; Xu, B.; Chen, H. Potential pathways of zinc deficiency-promoted tumorigenesis. Biomed. Pharmacother. 2021, 133, 110983. [Google Scholar] [CrossRef]
- Nardinocchi, L.; Pantisano, V.; Puca, R.; Porru, M.; Aiello, A.; Grasselli, A.; Leonetti, C.; Safran, M.; Rechavi, G.; Givol, D.; et al. Zinc downregulates HIF-1 and inhibited its activity in tumor cells in vitro and in vivo. PLoS ONE 2010, 5, e15048. [Google Scholar] [CrossRef]
- Alder, H.; Taccioli, C.; Chen, H.; Jiang, Y.; Smalley, K.J.; Fadda, P.; Ozer, H.G.; Huebner, K.; Farber, J.L.; Croce, C.M.; et al. Dysregulation of miR-31 and miR-21 induced by zinc-deficiency promotes esophageal cancer. Carcinogenesis 2012, 33, 1736–1744. [Google Scholar] [CrossRef] [PubMed]
- Janakiram, N.B.; Mohammed, A.; Madka, V.; Rao, C.V. Prevention and treatment of cancers by immune modulating nutrients. Mol. Nutr. Food Res. 2016, 60, 1275–1294. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S.; Beck, F.W.J.; Snell, D.C.; Kucuk, O. Zinc in cancer prevention. Nutr. Cancer 2009, 61, 879–887. [Google Scholar] [CrossRef]
- John, E.; Laskow, T.C.; Buchser, W.J.; Pitt, B.R.; Basse, P.H.; Butterfield, L.H.; Kalinski, P.; Lotze, M.T. Zinc in innate and adaptive tumor immunity. J. Trans. Med. 2010, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Skarajnowska, D.; Bobrowska-Korczak, B. Role of zinc in immune system and anti-cancer defense mechanisms. Nutrients 2019, 11, 2273. [Google Scholar] [CrossRef]
- Stamoulis, I.; Kouraklis, G.; Theocharis, S. Zin and the liver: An active interaction. Dig. Dis. Sci. 2007, 52, 1595–1612. [Google Scholar] [CrossRef]
- Grüngreiff, K.; Reinhold, D.; Wedemeyer, H. The role of zinc in liver cirrhosis. Ann. Hepatol. 2016, 15, 7–16. [Google Scholar] [CrossRef]
- Stepien, M.; Hughes, D.J.; Hybsier, S.; Bamia, C.; Tjønneland, A.; Overvad, K.; Affret, A.; His, M.; Boutron-Ruault, M.C.; Katzke, V.; et al. Circulating copper and zinc levels and risk of hepatobiliary cancers in Europeans. Br. J. Cancer 2017, 116, 688–696. [Google Scholar] [CrossRef]
- Nagasue, N.; Kohno, H.; Chang, Y.C.; Nakamura, T. Iron, copper and zinc levels in serum and cirrhotic liver of patients with and without hepatocellular carcinoma. Oncology 1989, 46, 293–296. [Google Scholar] [CrossRef]
- Moriyama, M.; Matsumura, H.; Fukushima, A.; Ohkido, K.; Arakawa, Y.; Nirei, K.; Yamagami, H.; Kaneko, M.; Tanaka, N.; Arakawa, Y. Clinical significance of evaluation of serum zinc concentrations in C-viral chronic liver disease. Dig. Dis. Sci. 2006, 51, 1967–1977. [Google Scholar] [CrossRef]
- Himoto, T.; Masaki, T. Current trends of essential trace elements in patients with chronic liver diseases. Nutrients 2020, 12, 2084. [Google Scholar] [CrossRef] [PubMed]
- Shigefuku, R.; Iwasa, M.; Katayama, K.; Eguchi, A.; Kawaguchi, T.; Shiraishi, K.; Ito, T.; Suzuki, K.; Koreeda, C.; Ohtake, T.; et al. Hypozincemia is associated with human hepatocarcinogenesis in hepatitis C virus-related liver cirrhosis. Hepatol. Res. 2019, 49, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Fang, A.; Chen, P.; Wang, X.; Liu, Z.; Zhang, D.; Luo, Y.; Liao, G.; Long, J.; Zhong, R.; Zhou, Z.; et al. Serum copper and zinc levels at diagnosis and hepatocellular carcinoma survival in the Guangdong Liver Cancer Cohort. Int. J. Cancer 2019, 144, 2823–2832. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Fan, X.; Gao, Y.; Zuo, Y.; Hong, M.; Xu, Y. The relationship between zinc deficiency and hepatocellular carcinoma associated with hepatitis B liver cirrhosis: A 10-year follow-up study. Biol. Trace Elem. Res. 2023, 20, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Yang, Y.; Li, H.L.; Zheng, W.; Gao, J.; Zhang, W.; Yang, G.; Shu, X.O.; Xiang, Y.B. Dietary trace element intake and liver cancer risk: Results from two population-based cohorts in China. Int. J. Cancer 2017, 140, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, G.; Tan, X.; Ke, K.; Zhao, B.; Cheng, N.; Dang, Y.; Liao, N.; Wang, F.; Zheng, X.; et al. MT1G serves as a tumor suppressor in hepatocellular carcinoma by interacting with p53. Oncogenesis 2019, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Udali, S.; De Santis, D.; Mazzi, F.; Moruzzi, S.; Ruzzenente, A.; Castagna, A.; Pattini, P.; Beschin, G.; Franceschi, A.; Guglielmi, A.; et al. Trace elements status and metallothioneins DNA methylation influence human hepatocellular carcinoma survival rate. Front. Oncol. 2021, 10, 596040. [Google Scholar] [CrossRef] [PubMed]
- Harimoto, N.; Araki, K.; Muranushi, R.; Hoshino, K.; Yamanaka, T.; Hagiwara, K.; Ishii, N.; Tsukagoshi, M.; Watanabe, A.; Shirabe, K. Significance of zinc deficiency in patients with hepatocellular carcinoma undergoing hepatic resection. Hepatol. Res. 2022, 52, 210–220. [Google Scholar] [CrossRef]
- Hiraoka, A.; Nagamatsu, K.; Izumoto, H.; Adachi, T.; Yoshino, T.; Tsuruta, M.; Aibiki, T.; Okudaira, T.; Yamago, H.; Iwasaki, R.; et al. Zinc deficiency as an independent prognostic factor for patients with early hepatocellular carcinoma due to hepatitis virus. Hepatol. Res. 2020, 50, 92–100. [Google Scholar] [CrossRef]
- Ozeki, I.; Nakajima, T.; Suii, H.; Tatsumi, R.; Yamaguchi, M.; Arakawa, T.; Kuwata, Y. Predictors of hepatocellular carcinoma after hepatitis C virus eradication following direct-acting antiviral treatment: Relationship with serum zinc. J. Clin. Biochem. Nutr. 2020, 66, 245–252. [Google Scholar] [CrossRef]
- Ebara, M.; Fukuda, H.; Hatano, R.; Yoshikawa, M.; Sugiura, N.; Saisho, H.; Kondo, F.; Yukawa, M. Metal contents in the liver of patients with chronic liver disease caused by hepatitis C virus. Reference to hepatocellular carcinoma. Oncology 2003, 65, 323–330. [Google Scholar] [CrossRef]
- Tashiro, H.; Kawamoto, T.; Okubo, T.; Koide, O. Variation in the distribution of trace elements in hepatoma. Biol. Trace Elem. Res. 2003, 95, 49–63. [Google Scholar] [CrossRef]
- Pan, Z.; Choi, S.; Ouadid-Ahidouch, H.; Yang, J.M.; Beattie, J.H.; Korichneva, I. Zinc transporters and dysregulated channels in cancers. Front. Biosci. 2017, 22, 623–643. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.B.; Levy, B.A.; Zou, J.; Hanna, N.; Desouki, M.M.; Bagasra, O.; Johnson, L.A.; Costello, L.C. ZIP 14 zinc transporter down-regulation and zinc depletion in the development and progression of hepatocellular cancer. J. Gastrointest. Cancer 2012, 43, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Weaver, B.P.; Zhang, Y.; Hiscox, S.; Guo, G.L.; Apte, U.; Taylor, K.M.; Sheline, C.T.; Wang, L.; Andrews, G.K. Zip4 (Slc39a4) expression is activated in hepatocellular carcinomas and functions to repress apoptosis, enhance cell cycle and increase migration. PLoS ONE 2010, 5, e13158. [Google Scholar] [CrossRef] [PubMed]
- Gartmann, L.; Wex, T.; Grüngreiff, K.; Reinhold, D.; Kakinski, T.; Malfertheiner, P.; Schütte, K. Expression of zinc transporter ZIP4, ZIP14 and ZnT9 in hepatic carcinogenesis-An immunohistochemical study. J. Trace Elem. Med. Biol. 2018, 49, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, R.; Yang, J.; Mo, S.; Quek, K.; Kok, C.H.; Cheng, X.D.; Tian, S.; Zhang, W.; Qin, J.J. Integrated bioinformatics analysis reveals key candidate genes and pathways associated with clinical outcomes in hepatocellular carcinoma. Front. Genet. 2020, 11, 814. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Li, K.J.; Mao, L.; Hu, X.; Zhao, W.J.; Hu, A.; Lian, H.Z.; Zheng, W.J. Effects of exogeneous zinc on cell cycle, apoptosis and viability of MDAMB231, HepG2 and 293 T cells. Biol. Trace Elem. Res. 2013, 154, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhang, Z.; Zhu, L.; Lu, M.; Li, Y.; Zhou, J.; Lu, X.; Du, Q. Polaprezinc inhibits liver fibrosis and proliferation in hepatocellular carcinoma. Mol. Med. Rep. 2017, 16, 5523–5528. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, R.; Himoto, T.; Yamada, M.; Mimura, S.; Fujita, K.; Tani, J.; Morishita, A.; Masaki, T. Antitumor effect of zinc acetate in hepatocellular carcinoma cell lines via the induction of apoptosis. J. Nutr. Sci. Vitaminol. 2022, 68, 303–311. [Google Scholar] [CrossRef]
- El-Desouky, M.A.; Fahmi, A.A.; Abdelkader, I.Y.; Nasraldin, K.M. Anticancer effect of amygdalin (vitamin B17) on hepatocellular carcinoma cell line (HepG2) in the presence and absence of zinc. Anticancer Agents Med. Chem. 2020, 20, 486–494. [Google Scholar] [CrossRef]
- Wu, R.; Mei, X.; Ye, Y.; Xue, T.; Wang, J.; Sun, W.; Lin, C.; Xue, R.; Zhang, J.; Xu, D. Zn(II)-curcumin solid dispersion impairs hepatocellular carcinoma growth and enhances chemotherapy by modulating gut microbiota-mediated zinc homeostasis. Pharmacol. Res. 2019, 150, 104454. [Google Scholar] [CrossRef]
- Prasad, S.; Lall, R. Zinc-curcumin based complexes in health and diseases: An approach in chemopreventive and therapeutic improvement. J. Trace Elem. Med. Biol. 2022, 73, 127023. [Google Scholar] [CrossRef]
- Hassan, H.F.H.; Mansour, A.M.; Abo-Youssef, A.M.H.; Elsadek, B.E.M.; Messiha, B.A.S. Zinc oxide nanoparticles as a novel anticancer approach; in vitro and in vivo evidence. Clin. Exp. Pharmacol. Physiol. 2017, 44, 235–243. [Google Scholar] [CrossRef]
- Matsuoka, S.; Matsumura, H.; Nakamura, H.; Oshiro, S.; Arakawa, Y.; Hayashi, J.; Sekine, N.; Nirei, K.; Yamagami, H.; Ogawa, M.; et al. Zinc supplementation improves the outcome of chronic hepatitis C and liver cirrhosis. J. Clin. Biochem. Nutr. 2009, 45, 292–303. [Google Scholar] [CrossRef]
- Matsumura, H.; Nirei, K.; Nakamura, H.; Arakawa, Y.; Higuchi, T.; Hayashi, J.; Yamagami, H.; Matsuoka, S.; Ogawa, M.; Nakajima, N.; et al. Zinc supplementation therapy improves the outcome of patients with chronic hepatitis, C. J. Clin. Biochem. Nutr. 2012, 51, 178–184. [Google Scholar]
- Hosui, A.; Tanimoto, T.; Okahara, T.; Ashida, M.; Ohnishi, K.; Wakahara, Y.; Kusumoto, Y.; Yamaguchi, T.; Sueyoshi, Y.; Hirao, M.; et al. Oral zinc supplementation decreases the risk of HCC development in patients with HCV eradication by, D.A.A. Hepatol. Commun. 2021, 5, 2001–2008. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Sandstead, H.H. Zinc requirements and the risks and benefits of zinc supplementation. J. Trace Elem. Med. Biol. 2006, 20, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Tsang, T.; Davis, C.I.; Brady, D.C. Copper biology. Curr. Biol. 2021, 31, R421–R427. [Google Scholar] [CrossRef] [PubMed]
- Lalioti, V.; Muruais, G.; Tsuchiya, Y.; Pulido, D.; Sandoval, I.V. Molecular mechanisms of copper homeostasis. Front. Biosci. 2009, 14, 4878–4903. [Google Scholar] [CrossRef]
- Wang, Z.; Jin, D. Regulatory roles of copper metabolism and cuproptosis in human cancers. Front. Oncol. 2023, 13, 1123420. [Google Scholar] [CrossRef]
- Chen, L.; Min, J.; Wang, F. Copper homeostasis and cuproptosis in human health and disease. Signal Transduct. Target. Ther. 2022, 7, 378. [Google Scholar] [CrossRef]
- Tang, X.; Yan, Z. Copper in cancer: From limiting nutrient to therapeutic target. Front. Oncol. 2023, 13, 1209156. [Google Scholar] [CrossRef]
- Hellman, N.E.; Gitlin, J.D. Ceruloplasmin metabolism and function. Ann. Rev. Nutr. 2002, 22, 439–458. [Google Scholar] [CrossRef] [PubMed]
- Arredondo, M.; Gonzalez, M.; Olivares, M.; Pizarro, F.; Araya, M. Ceruloplasmin, an indicator of copper status. Biol. Trace Elem. Res. 2008, 123, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Ward, D.M.; Kaplan, J. Ferroportin-mediated iron transport: Expression and regulation. Biochem. Biophys. Acta 2012, 1823, 1426–1433. [Google Scholar] [CrossRef] [PubMed]
- Gulec, S.; Collins, J.F. Molecular mediators govering iron-copper interactions. Ann. Rev. Nutr. 2014, 34, 95–116. [Google Scholar] [CrossRef] [PubMed]
- Michalczyk, K.; Cymbaluk-Ploska, A. The role of zinc and copper in gynecological malignancies. Nutrients 2020, 12, 3732. [Google Scholar] [CrossRef] [PubMed]
- Gupte, A.; Mumper, R.J. Elevated copper and oxidative stress in cancer cells as a target for cancer treatment. Cancer Treat. Rev. 2009, 35, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Denoyer, D.; Masaldan, S. Targeting copper in cancer therapy: Copper that cancer. Metallomics 2015, 7, 1459–1476. [Google Scholar] [CrossRef]
- De Luca, A.; Barile, A.; Arciello, M.; Rossi, L. Copper homeostasis as target of both consolidated and innovative strategies of anti-tumor therapy. J. Trace Elem. Med. Biol. 2019, 55, 2040213. [Google Scholar] [CrossRef]
- Han, I.W.; Jang, J.-Y.; Kwon, W.; Park, T.; Kim, Y.; Lee, K.B.; Kim, S.-W. Ceruloplasmin as a prognostic marker in patients with bile duct cancer. Oncotarget 2017, 8, 29028–29037. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Z.; Chen, J.G.; Chen, X.F.; Gu, D.H.; Liu, Z.M.; Gao, Y.D.; Zheng, B. Ceruloplasmin overexpression is associated with oncogenic pathways and poorer survival rates in clear-cell renal cell carcinoma. FEBS Open Bio 2021, 11, 2988–3004. [Google Scholar] [CrossRef] [PubMed]
- Voli, F.; Valli, E.; Lerra, L.; Kimpton, K.; Saletta, F.; Giorgi, F.M.; Mercatelli, D.; Rouaen, J.R.; Shen, S.; Murray, J.E.; et al. Intratumoral copper modulates PD-L1 expression and influences tumor immune evasion. Cancer Res. 2020, 80, 4129–4144. [Google Scholar] [CrossRef]
- Tsvetkov, P.; Coy, S.; Petrova, B.; Dreishpoon, M.; Verma, A.; Abdusamad, M.; Rossen, J.; Joesch-Cohen, L.; Humeidi, R.; Spangler, R.D.; et al. Copper indices cell death by targeting lipoylated TCA cycle proteins. Science 2022, 375, 1254–1261. [Google Scholar] [CrossRef]
- Yang, W.; Wang, Y.; Huang, Y.; Yu, J.; Wang, T.; Li, C.; Yang, L.; Zhang, P.; Shi, L.; Yin, Y.; et al. 4-octyl itaconate inhibits aerobic glycolysis by targeting GAPDH to promote cuproputosis in colorectal cancer. Bio Med. Pharmacother. 2023, 159, 114301. [Google Scholar] [CrossRef]
- Ishida, S.; McCormick, F.; Smith-McCune, K.; Schlatter, E. Enhancing tumor-specific uptake of anticancer drug cisplatin with a copper chelator. Cancer Cell 2010, 17, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-Y.; Choi, C.H.; Do, I.-G.; Song, S.Y.; Lee, W.; Park, H.S.; Song, T.J.; Kim, M.K.; Kim, T.-J.; Lee, J.-W.; et al. Prognostic value of the copper transporters, CTR1 and CTR2, in patients with ovarian carcinoma receiving platinum-based chemotherapy. Gynecol. Oncol. 2011, 122, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Shanbhag, V.C.; Gudekar, N.; Jasmer, K.; Papageorgiou, C.; Singh, K.; Petris, M.J. Copper metabolism as a unique vulnerability in cancer. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118893. [Google Scholar] [CrossRef]
- Komatsu, M.; Sumizawa, T.; Mutoh, M.; Chen, Z.S.; Terada, K.; Furukawa, T.; Yang, X.L.; Gao, H.; Miura, N.; Sugiyama, T.; et al. Copper-transporting P-type adenosine triphosphatase (ATP7B) is associated with cisplatin resistance. Cancer Res. 2000, 60, 1312–1316. [Google Scholar] [PubMed]
- Sugeno, H.; Takebayashi, Y.; Higashimoto, M.; Ogura, Y.; Shibukawa, G.; Kanzaki, A.; Terada, K.; Sugiyama, T.; Watanabe, K.; Katoh, R.; et al. Expression of copper-transporting P-type adenosine triphosphatase (ATP7B) in human hepatocellular carcinoma. Anticancer Res. 2004, 24, 1045–1048. [Google Scholar] [PubMed]
- Porcu, C.; Antonucci, L.; Barbaro, B.; Illi, B.; Nasi, S.; Martini, M.; Licata, A.; Miele, L.; Grieco, A.; Balsano, C. Copper/MYC/CTR1 interplay: A dangerous relationship in hepatocellular carcinoma. Oncotarget 2018, 9, 9325–9343. [Google Scholar] [CrossRef] [PubMed]
- Iwadate, H.; Ohira, H.; Suzuki, T.; Abe, K.; Yokokawa, J.; Takiguchi, J.; Rai, T.; Orikasa, H.; Irisawa, A.; Obara, K.; et al. Hepatocellular carcinoma associated with Wilson’s disease. Intern. Med. 2004, 43, 1042–1045. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffenberger, J.; Mogler, C.; Gotthardt, D.N.; Schulze-Bergkamen, H.; Litwin, T.; Reuner, U.; Hefter, H.; Huster, D.; Schemmer, P.; Członkowska, A.; et al. Hepatobiliary malignancies in Wilson disease. Liver Int. 2015, 35, 1615–1622. [Google Scholar] [CrossRef] [PubMed]
- Poo, J.L.; Rosas-Romero, R.; Montemayor, A.C.; Isoard, F.; Uribe, M. Diagnostic value of copper/zinc ratio in hepatocellular carcinoma: A case control study. J. Gastroenterol. 2003, 38, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zeng, J.W.; Ma, Q.; Zhang, S.; Tang, J.; Feng, J.F. Serum copper and zinc levels in breast cancer: A meta-analysis. J. Trace Elem. Med. Biol. 2020, 62, 126629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shao, J.; Tan, S.W.; Ye, H.P.; Shan, X.Y. Association between serum copper/zinc ratio and lung cancer: A systematic review with meta-analysis. J. Trace Elem. Med. Biol. 2022, 74, 127061. [Google Scholar] [CrossRef]
- Tamai, Y.; Iwasa, M.; Eguchi, A.; Shigefuku, R.; Sugimoto, K.; Hasegawa, H.; Takei, Y. Serum copper, zinc metallothionein serve as potential biomarker for hepatocellular carcinoma. PLoS ONE 2020, 15, e0237370. [Google Scholar] [CrossRef]
- Ebara, M.; Fukuda, H.; Hatano, R.; Saisho, H.; Nagato, Y.; Suzuki, K.; Nakajima, K.; Yukawa, M.; Kondo, F.; Nakayama, A.; et al. Relationship between copper, zinc and metallothionein in hepatocellular carcinoma and its surrounding liver parenchyma. J. Hepatol. 2000, 33, 415–422. [Google Scholar] [CrossRef]
- Casaril, M.; Capra, F.; Marchiori, L.; Gabrielli, G.B.; Nicoli, N.; Corso, F.; Baracchino, F.; Corrocher, R. Serum copper and ceruloplasmin in early and in advanced hepatocellular carcinoma: Diagnostic and prognostic relevance. Tumori 1989, 75, 498–502. [Google Scholar] [CrossRef]
- Ferrin, G.; Rodriguez-Perálvarez, M.; Aguilar-Melero, P.; Ranchal, I.; Liamoza, C.; Linares, C. Plasma protein biomarkers of hepatocellular carcinoma in HCV-infected alcoholic patients with cirrhosis. PLoS ONE 2015, 10, e0118527. [Google Scholar] [CrossRef]
- Yin, H.; Lin, Z.; Nie, S.; Wu, J.; Tan, Z.; Zhu, J.; Dai, J.; Feng, Z.; Marrero, J.; Lubman, D.M. Mass-selected site-specific core-fucosylation of ceruloplasmin in alcohol-related hepatocellular carcinoma. J. Proteome Res. 2014, 13, 2827–2896. [Google Scholar] [CrossRef]
- Gan, B. Mitochondrial regulation of ferroptosis. J. Cell Biol. 2021, 220, e202105043. [Google Scholar] [CrossRef]
- Shang, Y.; Luo, M.; Yao, F.; Wang, S.; Yuan, Z.; Yang, Y. Ceruloplasmin suppresses ferroptosis by regulating iron homeostasis in hepatocellular carcinoma cells. Cell Signal 2020, 72, 109663. [Google Scholar] [CrossRef]
- Wang, X.; Chen, D.; Shi, Y.; Luo, J.; Zhang, Y.; Yuan, X.; Zhang, C.; Shu, H.; Yu, W.; Tian, J. Copper and cuproptosis-related genes in hepatocellular carcinoma: Therapeutic biomarkers targeting tumor immune microenvironment and immune checkpoints. Front. Immunol. 2023, 14, 1123231. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Wu, J. Hypoxia inducible factor in hepatocellular carcinoma: A therapeutic target. World J. Gastroenterol. 2015, 21, 12171–12178. [Google Scholar] [CrossRef] [PubMed]
- Himoto, T.; Fujita, K.; Nomura, T.; Tani, J.; Miyoshi, H.; Morishita, A.; Yoneyama, H.; Kubota, S.; Haba, R.; Suzuki, Y.; et al. Roles of copoper in hepatocarcinogenesis via the activation of hypoxia-inducible factor-1a. Biol. Trace Elem. Res. 2016, 174, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Sone, K.; Maeda, M.; Wakabayashi, K.; Takeichi, N.; Mori, M.; Sugimura, T.; Nagao, M. Inhibition of hereditary hepatitis and liver tumor development in Long-Evans cinnamon rats by the copper-chelating agent trientine dihydrochloride. Hepatology 1996, 23, 764–770. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, J.; Yoshiji, H.; Kuriyama, S.; Ikenaka, Y.; Noguchi, R.; Okuda, H.; Tsujinoue, H.; Nakatani, T.; Kishida, H.; Nakae, D.; et al. The copper-chelating agent, trientine, suppresses tumor development and angiogenesis in the murine hepatocellular carcinoma cells. Int. J. Cancer 2000, 94, 768–773. [Google Scholar] [CrossRef]
- Moriguchi, M.; Nakajima, T.; Kimura, H.; Watanabe, T.; Takashima, H.; Mitsumoto, Y.; Katagishi, T.; Okanoue, T.; Kagawa, K. The copper chelator trientine has an antiangiogenic effect against hepatocellular carcinoma, possibly through inhibition of interleukin-8 production. Int. J. Cancer 2002, 102, 445–452. [Google Scholar] [CrossRef]
- Davis, C.I.; Gu, X.; Kiefer, R.M.; Ralle, M.; Gade, T.P.; Brady, D.C. Altered copper homeostasis underlies sensitivity of hepatocellular carcinoma to copper chelation. Metallomics 2020, 12, 1995–2008. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, S.Y.; Wang, L.H.; Wang, F.Y.; Wang, N.N.; Cao, Q.; Wang, Y.T.; Yang, J.Y.; Wu, C.F. Copper improves the anti-angiogenic activity of dusulfiram through the EGFR/Src/VEGF pathway in glioma. Cancer Lett. 2015, 369, 86–96. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.H.; Zhang, H.T.; Wang, Y.T.; Liu, S.; Zhou, W.L.; Yuan, X.Z.; Li, T.Y.; Wu, C.F.; Yang, J.Y. Disulfiram combined with copper inhibits metastasis and epithelial-mesenchymal transition in hepatocellular carcinoma through the NF-kB and TGF-b pathways. J. Cell Mol. Med. 2018, 22, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.F.; De Oliveira, A.C.; Santoro, A.; Raoul, J.L.; Forner, A.; et al. Sorafenib in advanced hepatocellular carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef]
- Ren, X.; Li, Y.; Zhou, Y.; Hu, W.; Yang, C.; Jing, Q.; Zhou, C.; Wang, X.; Hu, J.; Wang, L.; et al. Overcoming the compensatory elevation of NRF2 renders hepatocellular carcinoma cells more vulnerable to disulfiram/copper-induced ferroptosis. Redox Biol. 2021, 46, 102122. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Guo, L.; Zhang, B.; Liu, S.; Zhang, K.; Yan, J.; Zhang, W.; Yu, M.; Chen, Z.; Xu, Y.; et al. Disulfiram combined with copper induces immunosuppression via PD-L1 stabilization in hepatocellular carcinoma. Am. J. Cancer Res. 2019, 9, 2442–2455. [Google Scholar]
- Gao, F.; Yuan, Y.; Ding, Y.; Li, P.Y.; Chang, Y.; He, X.X. DLAT as a cuproptosis promotor and molecular target of elesclomol in hepatocellular carcinoma. Curr. Med. Sci. 2023, 43, 526–538. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Shi, Z.; Liu, X.; Jin, S.; Chen, P.; Zhang, Y.; Chen, G.; Fan, X.; Yang, J.; Lin, H. Identification and development of a novel risk model based on cuproptosis-associated RNA methylation regulators for predicting prognosis and characterizing immune status in hepatocellular carcinoma. Hepatol. Int. 2023, 17, 112–130. [Google Scholar] [CrossRef]
- Fu, Z.G.; Wang, L.; Cui, H.Y.; Peng, J.L.; Wang, S.J.; Geng, J.J.; Feng, F.; Song, F.; Li, L.; Zhu, P.; et al. A novel small-molecule compound targeting CD147 inhibits the motility and invasion of hepatocellular carcinoma cells. Oncotarget 2016, 7, 9429–9447. [Google Scholar] [CrossRef]
- Yan, L.; Zucker, S.; Toole, B.P. Roles of the multifunctional glycoprotein, emmprin (basigin; CD147), in tumour progression. Tromb. Haemost. 2005, 93, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Liu, H.; Chen, K.; Hu, X.; Ma, X.; Lan, X.; Zhang, Y.; Cheng, Z. Theranostics of malignant melanoma with 64CuCl2. J. Nucl. Med. 2014, 55, 812–817. [Google Scholar] [CrossRef] [PubMed]
- Wachsmann, J.; Peng, F. Molecular imaging and therapy targeting copper metabolism in hepatocellular carcinoma. World J. Gastroenterol. 2016, 22, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.A.; Alhadlaq, H.A.; Ahmad, J.; Al-Khedhairy, A.A.; Musarrat, J.; Ahamed, M. Copper oxide nanoparticles induced mitochondria mediated apoptosis in human hepatocarcinoma cells. PLoS ONE 2013, 8, e69534. [Google Scholar] [CrossRef] [PubMed]
- Brewer, G.J.; Dick, R.D.; Grover, D.K.; LeClaire, V.; Tseng, M.; Wicha, M.; Pienta, K.; Redman, B.G.; Jahan, T.; Sondak, V.K.; et al. Treatmentof metastatic cancer with tetrathiomolybdate, an anticopper, antiangiogenetic agent: Phase I study. Clin. Cancer Res. 2000, 6, 1–10. [Google Scholar] [PubMed]
- Redman, B.G.; Esper, P.; Pan, Q.; Dunn, R.L.; Hussain, H.K.; Chenevert, T.; Brewer, G.J.; Merajver, S.D. Phase II trial of trtrathiomolybdate in patients with advanced kidney cancer. Clin. Cancer Res. 2003, 9, 1666–1672. [Google Scholar] [PubMed]
- Ohkoshi-Yamada, M.; Kamimura, K.; Kamimura, H.; Terai, S. Rare complication of hepatocellular carcinoma in Wilson’s disease. JGH Open 2021, 5, 12201222. [Google Scholar] [CrossRef]
- Rayman, M.P. The importance of selenium to human health. Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef]
- Roman, M.; Jitaru, P.; Barbante, C. Selenium biochemistry and its role for human health. Metallomics 2014, 6, 25–54. [Google Scholar] [CrossRef]
- Razaghi, A.; Poorebrahim, M.; Sarhan, D.; Björnstedt, M. Selenium stimulates the antitumor immunity: Insights to future research. Eur. J. Cancer 2021, 155, 256–267. [Google Scholar] [CrossRef]
- Zeng, H.; Combs, G.F., Jr. Selenium as an antitumor nutrient: Roles in cell proliferation and tumor invasion. J. Nutr. Biochem. 2008, 19, 1–7. [Google Scholar] [CrossRef]
- Yildiz, A.; Kaya, Y.; Tanriverdi, D. Effect of interaction between selenium and zinc on DNA repair in association with cancer prevention. J. Cancer Prev. 2019, 24, 146–154. [Google Scholar] [CrossRef]
- Lee, K.H.; Jeong, D. Bimodal actions of selenium essential for antioxidant and toxic pro-oxidant activities: The selenium paradox(review). Mol. Med. Rep. 2012, 5, 299–304. [Google Scholar] [PubMed]
- Zeng, H. Selenite and selenomethionine promote HL-60 cell cycle progression. J. Nutr. 2002, 132, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Brozmanová, J.; Mániková, D.; Vlčková, V.; Chovanec, M. Selenium: A double-edged sword for defense and offence in cancer. Arch. Toxicol. 2010, 84, 919–938. [Google Scholar] [CrossRef] [PubMed]
- Wallenberg, M.; Misra, S.; Björnstedt, M. Selenium cytotoxicity in cancer. Basic. Clin. Pharmacol. Toxicol. 2014, 114, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.; Gandin, V. Selenium compounds as therapeutic agents in cancer. Biochim. Biophys. Acta 2015, 1850, 1642–1660. [Google Scholar] [CrossRef] [PubMed]
- Thirunavukkarasu, C.; Premkumar, K.; Sheriff, A.K.; Sakthisekaran, D. Sodium selenite enhances glutathione peroxidase activity and DNA single strand breaks in hepatoma induced by N-nitrosodiethylamine and promoted by Phenobarbital. Mol. Cell Biochem. 2008, 310, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, E.; Rudolf, K.; Červinka, M. Selenium activates p53 and p38 pathways and induces caspase-independent cell death in cervical cancer cells. Cell Biol. Toxicol. 2008, 24, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Meng, J.; Xu, T.J.; Qin, X.Y.; Zhou, X.D. Sodium selenite induces apoptosis in colon cancer cells via Bax-dependent mitochondrial pathway. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2166–2171. [Google Scholar]
- Corcoran, N.M.; Najdovska, M.; Costello, A.J. Inogranic selenium retards progression of experimental hormone refractory prostate cancer. J. Urol. 2004, 171, 907–910. [Google Scholar] [CrossRef]
- Kiremidjian-Schumacher, L.; Roy, M.; Glickman, R.; Schneider, K.; Rothstein, S.; Cooper, J.; Hochster, H.; Kim, M.; Newman, R. Selenium and immunocompetence in patients with head and neck cancer. Biol. Trace Elem. Res. 2000, 73, 97–111. [Google Scholar] [CrossRef]
- Avery, J.C.; Hoffmann, P.R. Selenium, selenoprotein and immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef]
- Lai, H.; Zeng, D.; Liu, C.; Zhang, Q.; Wang, X.; Chen, T. Selenium containing ruthenium complex synergizes with natural killer cells to enhance immunotherapy against prostate cancer via activating TRAIL/FasL signaling. Biomaterials 2019, 219, 119377. [Google Scholar] [CrossRef]
- Wang, J.; Chang, Y.; Luo, H.; Jiang, W.; Xu, L.; Chen, T.; Zhu, X. Designing immunogenic nanotherapeutic for photothermal-triggered immunotherapy involving reprogramming immunosuppression and activating systemic antitumor responses. Biomaterials 2020, 255, 120153. [Google Scholar] [CrossRef]
- Meyer, F.; Galan, P.; Douville, P.; Bairati, I.; Kegle, P.; Bertrais, S.; Estaquio, C.; Hercberg, S. Antioxidant vitamin and mineral supplementation and prostate cancer prevention in the SU.VI.MAX trial. Int. J. Cancer 2005, 116, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.A.; Lee, E.H.; Cho, H.; Kim, J.H. High dose of selenium induces ferroptotic cell death in ovarian cancer. Int. J. Mol. Sci. 2023, 24, 1918. [Google Scholar] [CrossRef] [PubMed]
- Kalimuthu, K.; Keerthana, C.K.; Mohan, M.; Arivalagan, J.; Christyraj, J.R.S.S.; Firer, M.A.; Choudry, M.H.A.; Anto, R.J.; Lee, Y.J. Emerging role of selenium metabolic pathways in cancer: New therapeutic target for cancer. J. Cell Biochem. 2022, 123, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-D.; Liu, Z.-Y.; Wang, M.-S.; Guo, Y.-X.; Wang, X.-K.; Luo, K.; Huang, S.; Li, R.-F. Mechanisms and regulations of ferroptosis. Front. Immunol. 2023, 14, 1269451. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yang, W.; Long, G.; Wei, C. Trace elements and chemotherapy sensitivity. Biol. Trace Elem. Res. 2016, 173, 283–290. [Google Scholar] [CrossRef]
- Abd-Rabou, A.A.; Shalby, A.B.; Ahmed, H.H. Selenium nanoparticle induce the chemosensitivity of fluorouracil nanoparticles in breast and colon cancer cells. Biol. Trace Elem. Res. 2019, 187, 80–91. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Khabatova, V.V.; Gudkov, S.V.; Turovsky, E.A. Ca2+-dependent effects of selenium-sorafenib nanocomplexs on glioblastoma cells and astrocytes of the cerebral cortex: Anticancer agent and cytoprotector. Int. J. Mol. Sci. 2023, 24, 2411. [Google Scholar] [CrossRef] [PubMed]
- Himoto, T.; Yoneyama, H.; Kurokohchi, K.; Inukai, M.; Masugata, H.; Goda, F.; Haba, R.; Watababe, S.; Kubota, S.; Senda, S.; et al. Selenium deficiency is associated with insulin resistance in patients with hepatitis C virus-related chronic liver disease. Nutr. Res. 2011, 31, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Bettinger, D.; Schultheiss, M.; Hennecke, N.; Panther, E.; Knüppel, E.; Blum, H.E.; Thimme, R.; Spangenberg, H.C. Selenium levels in patients with hepatitis C virus-related chronic hepatitis, liver cirrhosis, and hepatocellular carcinoma: A pilot study. Hepatology 2013, 57, 2543–2544. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.W.; Bae, S.M.; Kim, Y.W.; Liu, H.B.; Bae, S.H.; Choi, J.Y.; Yoon, S.K.; Chaturvedi, P.K.; Battogtokh, G.; Ahn, W.S. Serum selenium levels in Krean hepatoma patients. Biol. Trace Elem. 2012, 148, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.J.; Duarte-Salles, T.; Hybsier, S.; Trichopoulou, A.; Stepien, M.; Aleksandrova, K.; Overvad, K.; Tjønneland, A.; Olsen, A.; Affret, A.; et al. Prediagnostic selenium status and hepatobiliary cancer risk in the European prospective investigation into cancer and nutrition cohort. Am. J. Clin. Nutr. 2016, 104, 406–414. [Google Scholar] [CrossRef]
- Zhang, Z.; Bi, M.; Liu, Q.; Yang, J.; Xu, S. Meta-analysis of the correlation between selenium and incidence of hepatocellular carcinoma. Oncotarget 2016, 7, 77110–77117. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Dong, F.; Geng, Y.; Zhuang, H.; Ma, Z.; Zhou, Z.; Huang, B.; Sun, Z.; Hou, B. Selenium concentration, dietary intake and risk of hepatocellular carcinoma-A systemic review with meta-analysis. Nutr. Hosp. 2019, 36, 1430–1437. [Google Scholar]
- Rohr-Udilova, N.; Sieghart, W.; Eferl, R.; Stoiber, D.; Björkhem-Bergman, L.; Eriksson, L.C.; Stolze, K.; Hayden, H.; Keppler, B.; Sagmeister, S.; et al. Antagonistic effects of selenium and lipid peroxides on growth control in early hepatocellular carcinoma. Hepatology 2012, 55, 1112–1121. [Google Scholar] [CrossRef]
- Stasio, M.D.; Volpe, M.G.; Colonna, G.; Nazzaro, M.; Polimeno, M.; Scala, S.; Castello, G.; Costantini, S. A possible predictive marker of progression for hepatocellular carcinoma. Oncol. Lett. 2011, 2, 1247–1251. [Google Scholar] [CrossRef] [PubMed]
- Rohr-Udilova, N.; Bauer, E.; Timelthaler, G.; Eferl, R.; Stolze, K.; Pinter, M.; Seif, M.; Hayden, H.; Reiberger, T.; Schulte-Hermann, R.; et al. Impact of glutathione peroxidase 4 on cell proliferation, angiogenesis and cytokine production in hepatocellular carcinoma. Oncotarget 2018, 9, 10054–10068. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.D.; Moura, A.C.; Andreolla, H.F.; Veiga, A.B.; Fiegenbaum, M.; Giovenardi, M.; Almeida, S. Gene expression evaluation of antioxidant enzymes in patients with hepatocellular carcinoma: RT-qPCR and bioinformatic analyses. Genet. Mol. Biol. 2021, 44, 20190373. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, E.; Capone, F.; Accardo, M.; Sorice, A.; Costantini, M.; Colonna, G.; Castello, G.; Costantini, S. GPX4 and GPX7 over-expression in human hepatocellular carcinoma tissues. Eur. J. Histochem. 2015, 59, 283–288. [Google Scholar] [CrossRef]
- Radomska, D.; Czarnomysy, R.; Radomski, D.; Bielawska, A.; Bielawski, K. Selenium as a bioactive micronutrient in the human diet and its cancer chemopreventive activity. Nutrients 2021, 13, 1649. [Google Scholar] [CrossRef] [PubMed]
- Elhodaky, M.; Diamond, A.M. Selenium-binding protein 1 in human health and disease. Int. J. Mol. Sci. 2018, 19, 3437. [Google Scholar] [CrossRef] [PubMed]
- Corona, G.; De Lorenzo, E.; Elia, C.; Simula, M.P.; Avellini, C.; Baccarani, U.; Lupo, F.; Tiribelli, C.; Colombatti, A.; Toffoli, G. Differential proteomic analysis of hepatocellular carcinoma. Int. J. Oncol. 2010, 36, 93–99. [Google Scholar] [CrossRef]
- Huang, C.; Ding, G.; Gu, C.; Zhou, J.; Kuang, M.; Ji, Y.; He, Y.; Kondo, T.; Fan, J. Decreased selenium-binding protein 1 enhances glutathione peroxidase 1 activity and downregulates HIF-1a to promote hepatocellular carcinoma invasiveness. Clin. Cancer Res. 2012, 18, 3042–3053. [Google Scholar] [CrossRef]
- Gao, P.T.; Ding, G.Y.; Yang, X.; Dong, R.Z.; Hu, B.; Zhu, X.D.; Cai, J.B.; Ji, Y.; Shi, G.M.; Shen, Y.H.; et al. Invasive potential of hepatocellular carcinoma is enhanced by loss of selenium-binding protein 1 and subsequent upregulation of CXCR4. Am. J. Cancer Res. 2018, 8, 1040–1049. [Google Scholar]
- Sutton, A.; Nahon, P.; Pessayre, D.; Rufat, P.; Poiré, A.; Ziol, M.; Vidaud, D.; Barget, N.; Ganne-Carrié, N.; Charnaux, N.; et al. Genetic polymorphisms in antioxidant enzymes modulated hepatic iron accumulation and hepatocellular carcinoma development in patients with alcohol-induced cirrhosis. Cancer Res. 2006, 66, 2844–2852. [Google Scholar] [CrossRef]
- Lincoln, D.T.; Ali Emadi, E.M.; Torissen, K.F.; Clarke, F.M. The thioredoxin-thioredoxin reductase system: Over-expression in human cancer. Anticancer Res. 2003, 23, 2425–2433. [Google Scholar]
- Fu, B.; Meng, W.; Zeng, X.; Zhao, H.; Liu, W.; Zhang, T. TXNR1 is an unfavorable prognostic factor for patients with hepatocellular carcinoma. BioMed Res. Int. 2017, 2017, 4698167. [Google Scholar] [CrossRef]
- Lee, D.; Xu, I.M.; Chiu, D.K.; Leibold, J.; Tse, A.P.; Bao, M.H.; Yuen, V.W.; Chan, C.Y.; Lai, R.K.; Chin, D.W.; et al. Induction of oxidative stress via inhibition of thioredoxin reductase 1 is an effective therapeutic approach for hepatocellular carcinoma. Hepatology 2019, 69, 1768–1786. [Google Scholar] [CrossRef]
- Li, C.; Peng, Y.; Mao, B.; Qian, K. Thioredoxin reductase: A novel, independent prognostic marker in patients with hepatocellular carcinoma. Oncotarget 2015, 6, 17792–17804. [Google Scholar] [CrossRef]
- Hua, S.; Quan, Y.; Zhan, M.; Liao, H.; Li, Y.; Lu, L. miR-125-5p inhibits cell proliferation, migration, and invasion in hepatocellular carcinoma via targeting TXNRD1. Cancer Cell Int. 2019, 19, 203. [Google Scholar] [CrossRef]
- Celik, H.A.; Aydin, H.H.; Deveci, R.; Terzioglu, E.; Karacali, S.; Saydam, G.; Akarca, U.; Batur, Y. Biochemical and morphological characteristics of selenite-induced apoptosis in human hepatoma HepG2 cells. Biol. Trace Elem. Res. 2004, 99, 27–40. [Google Scholar] [CrossRef]
- Borda, L.J.; Perper, M.; Keri, J.E. Treatment of seborrheic dermatitis: A comprehensive review. J. Dermatol. Treat. 2019, 30, 158–169. [Google Scholar] [CrossRef]
- Yang, T.; Huo, J.; Xu, R.; Su, Q.; Tang, W.; Zhang, D.; Zhu, M.; Zhan, Y.; Dai, B.; Zhang, Y. Selenium sulfide disrupts the PLAGL2/C-MET/STAT3-induced resistance against mitochondrial apoptosis in hepatocellular carcinoma. Clin. Transl. Med. 2021, 11, e536. [Google Scholar] [CrossRef] [PubMed]
- Cuello, S.; Ramos, S.; Mateos, R.; Martín, M.A.; Madrid, Y.; Cámara, C.; Bravo, L.; Goya, L. Selenium methylselenocysteine protects human hepatoma HepG2 cells against oxidative stress induced by tert-butyl hydroperoxide. Anal. Bilanal. Chem. 2007, 389, 2167–2178. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.G.; Zhao, H.J.; Liu, Y.J.; Liu, Y.W.; Wang, X.L. Effect of two selenium sources on hepatocarcinogenesis and several angiogenic cytokines in diethylnitrosamine-induced hepatocarcinoma rats. J. Trace Elem. Med. Biol. 2012, 26, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Li, Q.; Deng, C.; Wang, Y.; Ding, Y.; Zhang, W.; Chen, Y.; Zhao, T.; Wei, F.; Yang, L.; et al. The synergism and attenuation effect of selenium (Se)-enriched Grifola frondose-polysaccharide on 5-fluorouracil (5-Fu) in Heps-bearing mice. Int. J. Biol. Macromol. 2018, 107, 2211–2216. [Google Scholar] [CrossRef]
- Khurana, A.; Tekula, S.; Saifi, M.A.; Venkatesh, P.; Godugu, C. Therapeutic application of selenium nanoparticles. Biomed. Pharmacother. 2019, 111, 802–812. [Google Scholar] [CrossRef]
- Singh, D.; Singh, M. Hepatocellular-targeted mRNA delivery using functionalized selenium nanoparticles in vitro. Pharmaceutics 2021, 13, 298. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Zhong, J.; Zhao, M.; Tang, Y.; Han, N.; Hua, L.; Xu, T.; Wang, C.; Zhu, B. Galactose-modified selenium nanoparticles for targeted delivery of doxorubicin to hepatocellular carcinoma. Drug Deliv. 2019, 26, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Al-Noshokaty, T.M.; Mesbah, N.M.; Abo-Elmatty, D.M.; Abulsoud, A.I.; Abdel-Hamed, A.R. Selenium nanoparticles overcome sorafenib resistance in thioacetamide induced hepatocellular carcinoma in rats by modulation of mTOR, NF-kB pathways and LncRNA-AF085935/GPC3 axis. Life Sci. 2022, 303, 120675. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Ma, W.; Sun, R.; Yin, H.; Lin, F.; Liu, Y.; Xu, W.; Zeng, H. Butaselen prevents hepatocarcinogenesis and progression through inhibiting thioredoxin reductase activity. Redox Biol. 2018, 14, 237–249. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, W.; Lv, X.; Weng, Q.; Chen, M.; Cui, R.; Liang, G.; Ji, J. Piperlongumine, a novel TrxR1 inhibitor, induces apoptosis in hepatocellular carcinoma cells by ROS-mediated ER stress. Front. Pharmacol. 2019, 10, 1180. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Yin, H.; Bai, M.; Liu, J.; Liu, R.; Zeng, H.; Wen, J. A novel TrxR1 inhibitor regulates NK and CD8+ T cell infiltration and cytotoxicity, enhancing the efficacy of anti-PD-1 immunotherapy against hepatocellular carcinoma. J. Immunol. 2023, 210, 681–695. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Reference | Study Design | Assigned Patients | Formulation | Dosage and Duration | Outcomes |
|---|---|---|---|---|---|
| Matsuoka et al. (2009) [66] | Prospective study (Zn responders vs. Zn non-responders) | HCV-related CLD (n = 32) | Polaprezinc | 150 mg, 5 years | Lower incidence of HCC in Zn responders |
| Matsumura et al. (2012) [67] | Randomized control study (Zn group vs. untreated group) | HCV-related CLD (n = 62) | Polaprezinc | 150 mg, 10 years | Lower incidence of HCC in Zn group |
| Hosui et al. (2021) [68] | Retrospective study (Zn group vs. untreated group) | HCV-related CLD who achieved SVR (n = 599) | Zinc sulfate or zinc acetate | Zinc 50–150 mg, 3 years | Lower incidence of HCC in Zn group |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Himoto, T.; Masaki, T. Current Trends on the Involvement of Zinc, Copper, and Selenium in the Process of Hepatocarcinogenesis. Nutrients 2024, 16, 472. https://doi.org/10.3390/nu16040472
Himoto T, Masaki T. Current Trends on the Involvement of Zinc, Copper, and Selenium in the Process of Hepatocarcinogenesis. Nutrients. 2024; 16(4):472. https://doi.org/10.3390/nu16040472
Chicago/Turabian StyleHimoto, Takashi, and Tsutomu Masaki. 2024. "Current Trends on the Involvement of Zinc, Copper, and Selenium in the Process of Hepatocarcinogenesis" Nutrients 16, no. 4: 472. https://doi.org/10.3390/nu16040472
APA StyleHimoto, T., & Masaki, T. (2024). Current Trends on the Involvement of Zinc, Copper, and Selenium in the Process of Hepatocarcinogenesis. Nutrients, 16(4), 472. https://doi.org/10.3390/nu16040472