
Selection of Fermentation Supernatant from Probiotic Strains Exhibiting Intestinal Epithelial Barrier Protective Ability and Evaluation of Their Effects on Colitis Mouse and Weaned Piglet Models

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Fermentation Supernatant Preparation
2.2. Cell Culture
2.3. CFS Treatment on Intestinal Cell Viability and Proliferation In Vitro
2.4. Caco-2 Intestinal Epithelial Monolayer Preparation and Transepithelial Electrical Resistance (TEER) Measurement
2.5. DSS-Induced Colitis Mice
2.6. Disease Activity Index
2.7. Histological Evaluation of the Colon
2.8. Determination of Serum Cytokines
2.9. In Vivo Intestinal Permeability Assay
2.10. Effects of CFS on Weaned Piglets
2.11. Preparation of CFS Powder for Feedstuffs
2.12. Piglet Body Weight and Growth Performance
2.13. Measurement of Cytokine Production and Endotoxin Levels in Serum
2.14. Statistical Analysis
3. Results
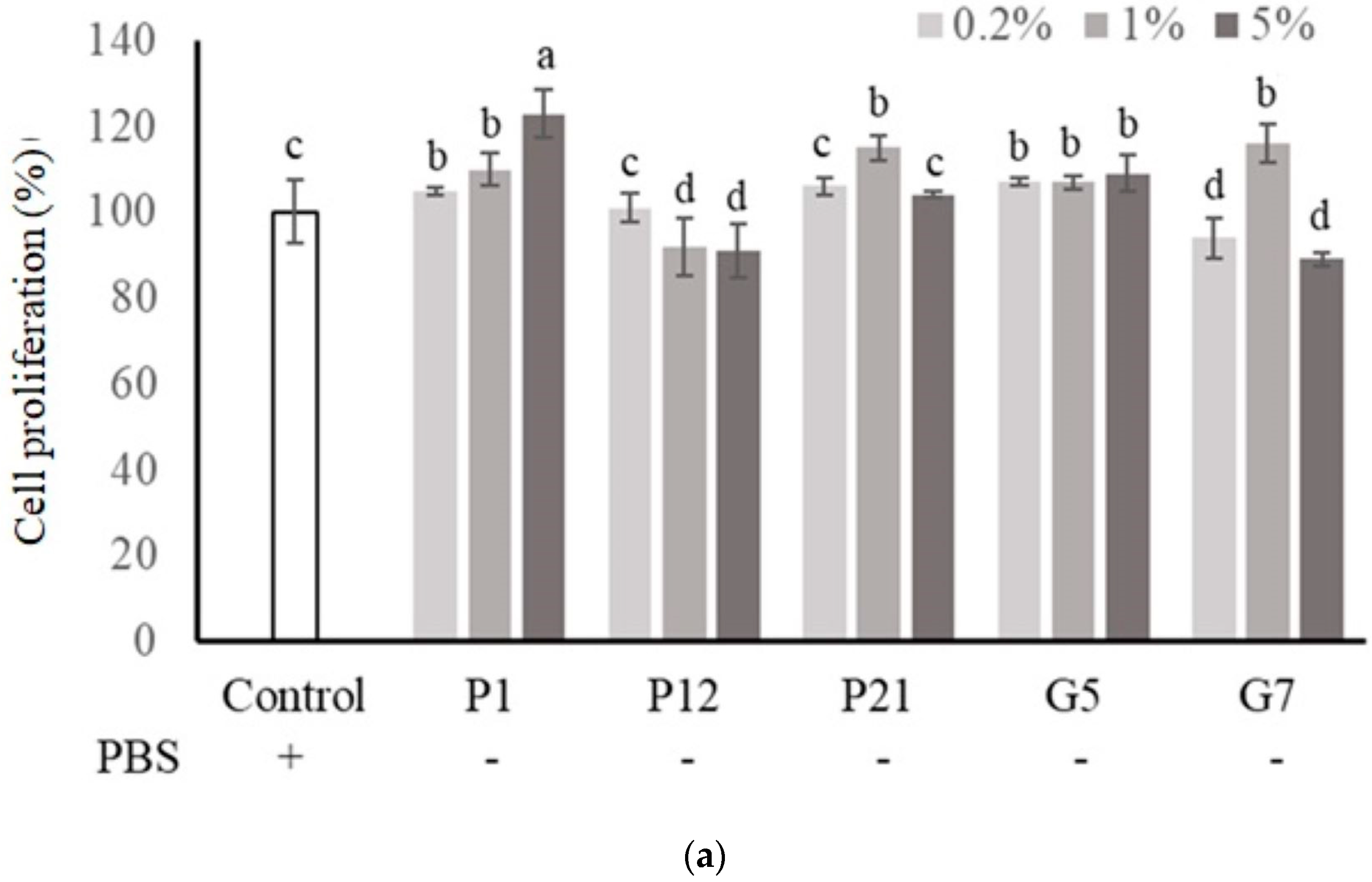
3.1. Preliminary Screening of Potential CFSs That Could Enhance Intestinal Epithelial Cell Viability
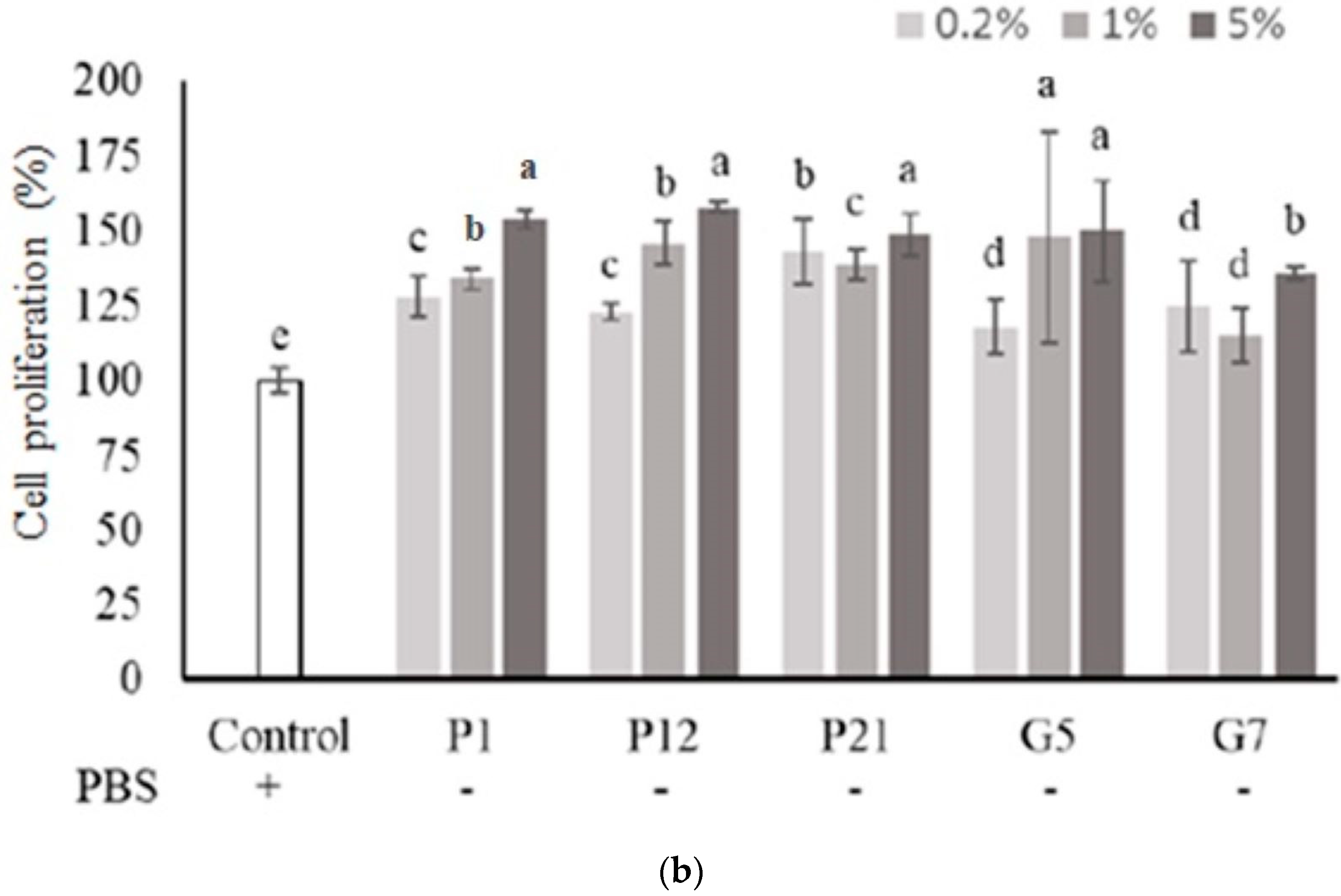
3.2. Effect of Selected CFSs on Intestinal Epithelial Cell Proliferation
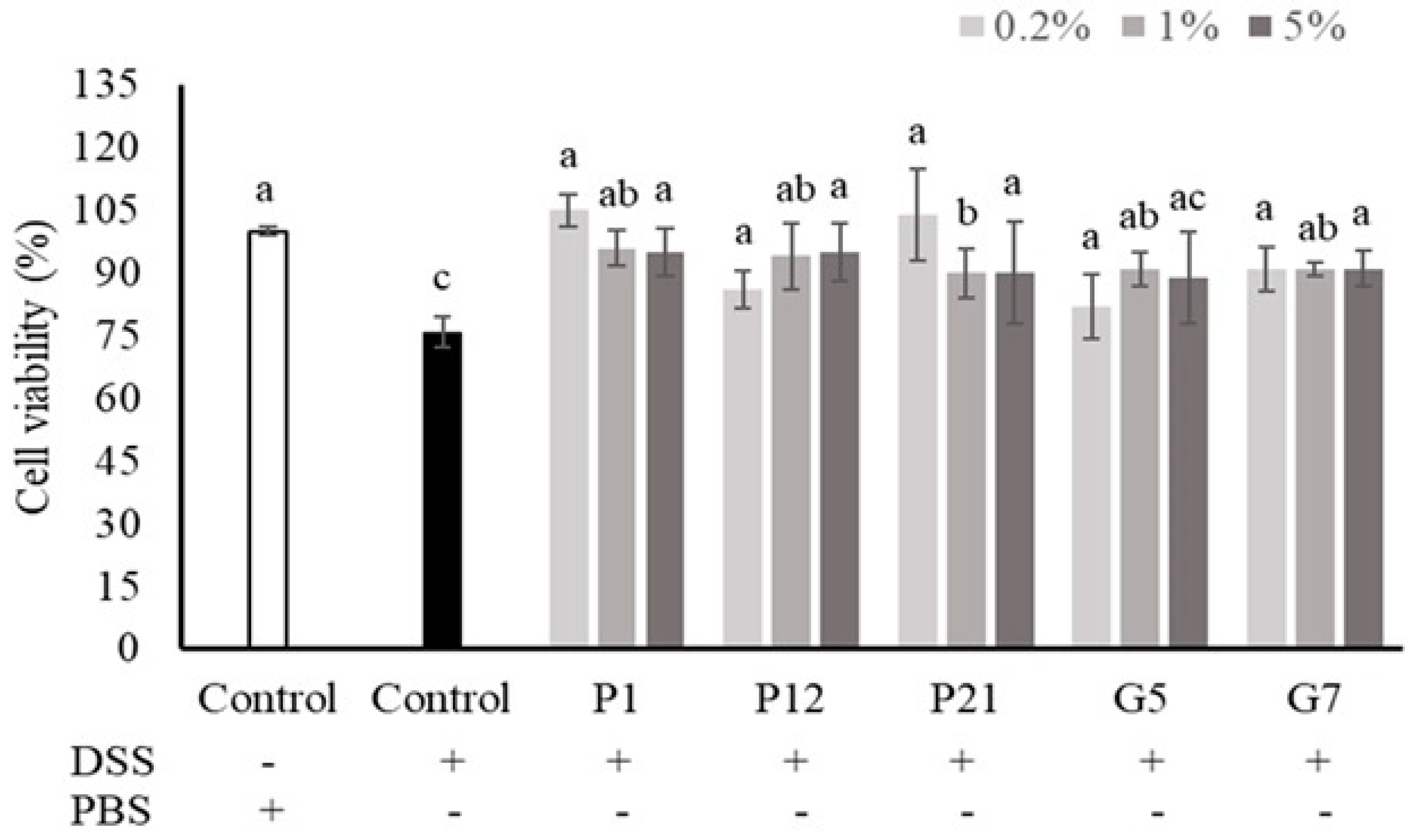
3.3. Effect of Selected CFSs on the Viability of DSS-Damaged Intestinal Epithelial Cells
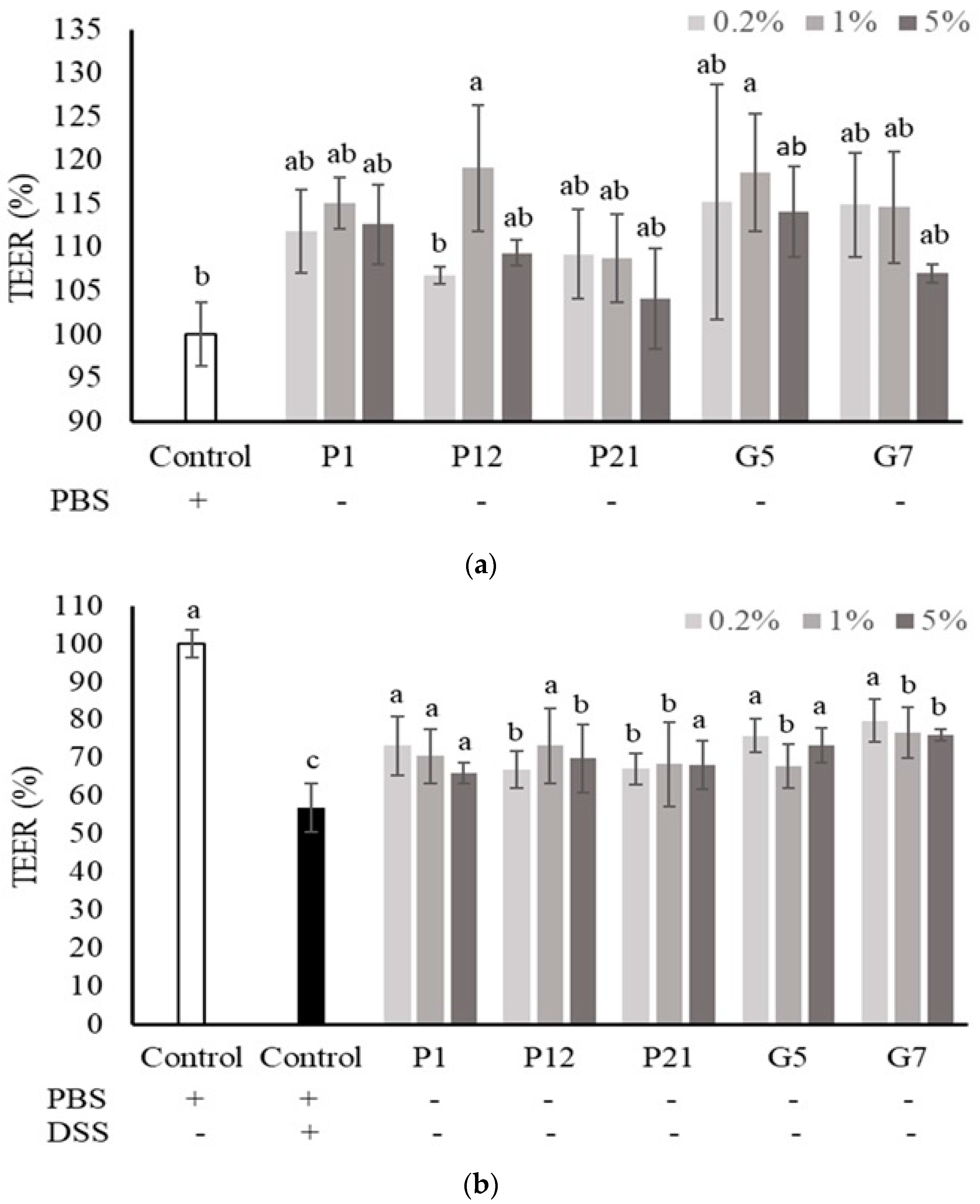
3.4. Effect of Selected CFSs on Transepithelial Electrical Resistance (TEER) of Caco-2 Cell Monolayers
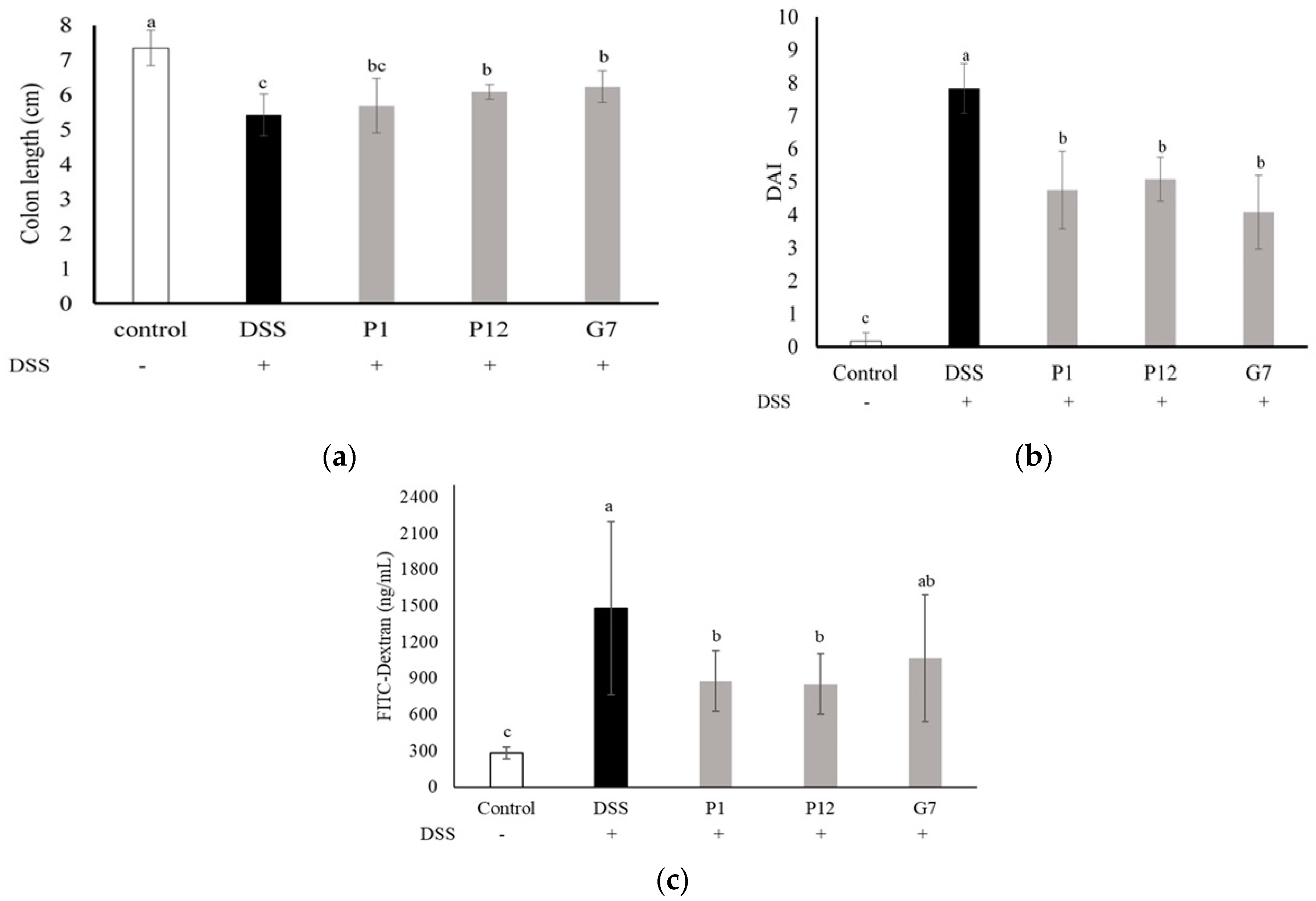
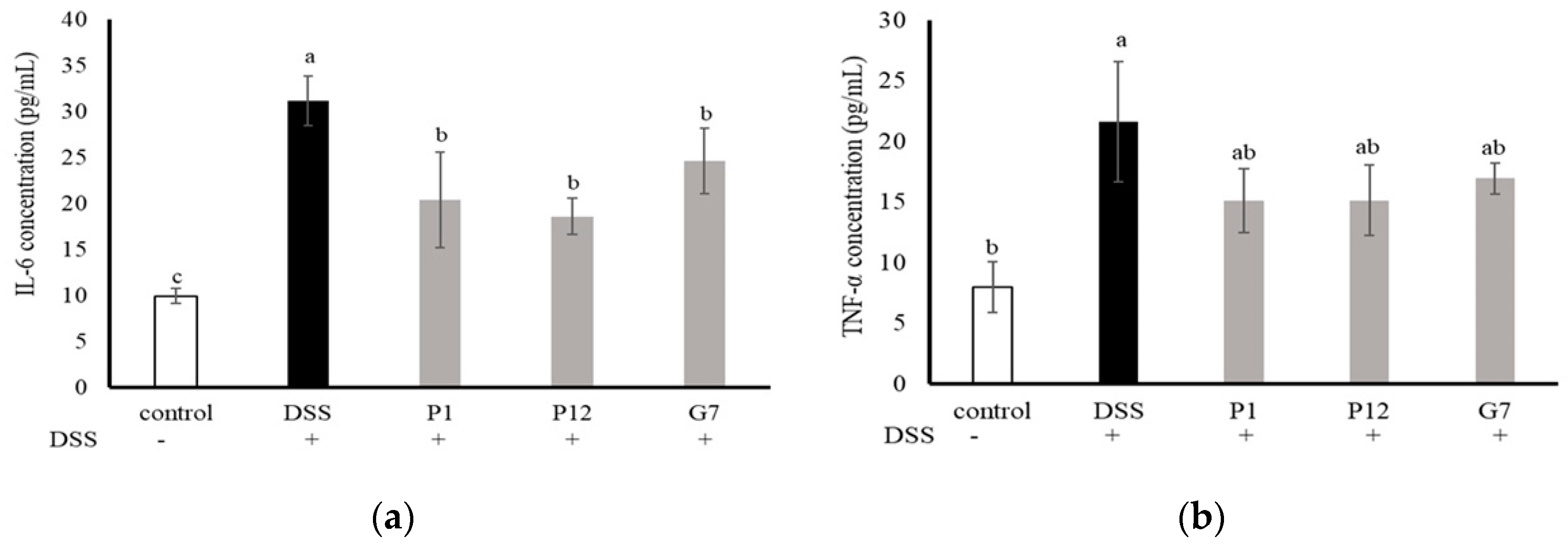
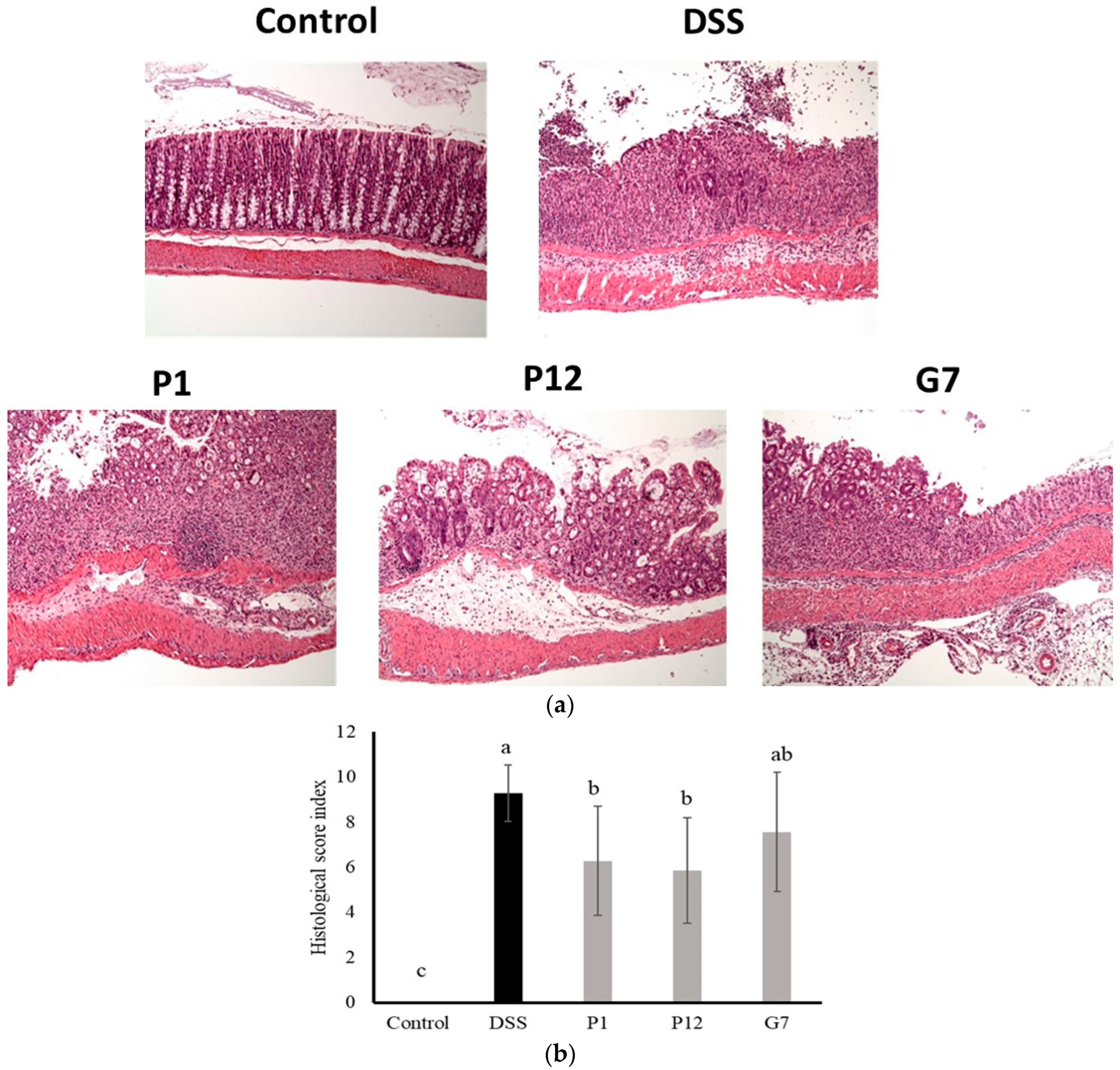
3.5. Effects of Selected CFSs on DSS-Induced Colitis in Mice
3.6. Effects of Selected CFSs on Weaned Piglets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schoultz, I.; Keita, Å.V. The Intestinal Barrier and Current Techniques for the Assessment of Gut Permeability. Cells 2020, 9, 1909. [Google Scholar] [CrossRef] [PubMed]
- Laukoetter, M.G.; Nava, P.; Nusrat, A. Role of the intestinal barrier in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Rescigno, M. The intestinal epithelial barrier in the control of homeostasis and immunity. Trends Immunol. 2011, 32, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Elamin, E.E.; Masclee, A.A.; Dekker, J.; Jonkers, D.M. Ethanol metabolism and its effects on the intestinal epithelial barrier. Nutr. Rev. 2013, 71, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.V.; Taylor, W.R. Intestinal barrier dysfunction as a therapeutic target for cardiovascular disease. Am. J. Physiol. Hear. Circ. Physiol. 2020, 319, H1227–H1233. [Google Scholar] [CrossRef]
- Han, X.; Lee, A.; Huang, S.; Gao, J.; Spence, J.R.; Owyang, C. Lactobacillus rhamnosus GG prevents epithelial barrier dysfunction induced by interferon-gamma and fecal supernatants from irritable bowel syndrome patients in human intestinal enteroids and colonoids. Gut Microbes 2019, 10, 59–76. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, P.; Zhang, X. Probiotics regulate gut microbiota: An effective method to improve immunity. Molecules 2021, 26, 6076. [Google Scholar] [CrossRef]
- Marteau, P.; Shanahan, F. Basic aspects and pharmacology of probiotics: An overview of pharmacokinetics, mechanisms of action and side-effects. Bailliere’s Best Pract. Res. Clin. Gastroenterol. 2003, 17, 725–740. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Vinderola, G.; Sanders, M.E.; Salminen, S. The Concept of Postbiotics. Foods 2022, 11, 1077. [Google Scholar] [CrossRef] [PubMed]
- Ozma, M.A.; Abbasi, A.; Akrami, S.; Lahouty, M.; Shahbazi, N.; Ganbarov, K.; Pagliano, P.; Sabahi, S.; Köse, Ş.; Yousefi, M.; et al. Postbiotics as the key mediators of the gut microbiota-host interactions. Infez. Med. 2022, 30, 180–193. [Google Scholar] [PubMed]
- Malagón-rojas, J.N.; Mantziari, A.; Salminen, S.; Szajewska, H. Postbiotics for preventing and treating common infectious diseases in children: A systematic review. Nutrients 2020, 12, 389. [Google Scholar] [CrossRef] [PubMed]
- Thorakkattu, P.; Khanashyam, A.C.; Shah, K.; Babu, K.S.; Mundanat, A.S.; Deliephan, A.; Deokar, G.S.; Santivarangkna, C.; Nirmal, N.P. Postbiotics: Current Trends in Food and Pharmaceutical Industry. Foods 2022, 11, 3094. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Wei, H.; Yuan, S.; Kong, Y.; Yang, H.; Zhang, Y.; Cui, X.; Chen, W.; Liu, J.; Zhang, Y. Probiotic Pediococcus pentosaceus ameliorates MPTP-induced oxidative stress via regulating the gut microbiota–gut–brain axis. Front. Cell. Infect. Microbiol. 2022, 12, 1022879. [Google Scholar] [CrossRef]
- Cicenia, A.; Santangelo, F.; Gambardella, L.; Pallotta, L.; Iebba, V.; Scirocco, A.; Marignani, M.; Tellan, G.; Carabotti, M.; Corazziari, E.S.; et al. Protective Role of Postbiotic Mediators Secreted by Lactobacillus rhamnosus GG Versus Lipopolysaccharide-induced Damage in Human Colonic Smooth Muscle Cells. J. Clin. Gastroenterol. 2016, 50, S140–S144. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Hsu, C.A.; Hung, W.T.; Chen, M.J. Effects of Lactobacillus paracasei 01 fermented milk beverage on protection of intestinal epithelial cell in vitro. J. Sci. Food Agric. 2016, 96, 2154–2160. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of cell viability by the MTT assay. Cold Spring Harb. Protoc. 2018, 2018, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Adnan, H.; Quach, H.; MacIntosh, K.; Antenos, M.; Kirby, G.M. Low Levels of GSTA1 Expression Are Required for Caco-2 Cell Proliferation. PLoS ONE 2012, 7, e51739. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. Biochem. Mol. Genet. Mech. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Sann, H.; VonErichsen, J.; Hessmann, M.; Pahl, A.; Hoffmeyer, A. Efficacy of drugs used in the treatment of IBD and combinations thereof in acute DSS-induced colitis in mice. Life Sci. 2013, 92, 708–718. [Google Scholar] [CrossRef]
- Wirtz, S.; Neufert, C.; Weigmann, B.; Neurath, M.F. Chemically induced mouse models of intestinal inflammation. Nat. Protoc. 2007, 2, 541–546. [Google Scholar] [CrossRef]
- Wirtz, S.; Popp, V.; Kindermann, M.; Gerlach, K.; Weigmann, B.; Fichtner-Feigl, S.; Neurath, M.F. Chemically induced mouse models of acute and chronic intestinal inflammation. Nat. Protoc. 2017, 12, 1295–1309. [Google Scholar] [CrossRef]
- Erben, U.; Christoph Loddenkemper, K.D.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kühl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar]
- Dieleman, L.A.; Palmen, M.J.H.J.; Akol, H.; Bloemena, E.; Peña, A.S.; Meuwissen, S.G.M.; Van Rees, E.P. Chronic experimental colitis induced by dextran sulphate sodium (DSS) is characterized by Th1 and Th2 cytokines. Clin. Exp. Immunol. 1998, 114, 385–391. [Google Scholar] [CrossRef]
- Vlantis, K.; Wullaert, A.; Polykratis, A.; Kondylis, V.; Dannappel, M.; Schwarzer, R.; Welz, P.; Corona, T.; Walczak, H.; Weih, F.; et al. NEMO prevents RIP kinase 1-mediated epithelial cell death and chronic intestinal inflammation by NF-κB-dependent and -independent functions. Immunity 2016, 44, 553–567. [Google Scholar] [CrossRef]
- Baxter, M.F.A.; Merino-Guzman, R.; Latorre, J.D.; Mahaffey, B.D.; Yang, Y.; Teague, K.D.; Graham, L.E.; Wolfenden, A.D.; Hernandez-Velasco, X.; Bielke, L.R.; et al. Optimizing fluorescein isothiocyanate dextran measurement as a biomarker in a 24-h feed restriction model to induce gut permeability in broiler chickens. Front. Vet. Sci. 2017, 4, 56. [Google Scholar] [CrossRef]
- Patience, J.F.; Rossoni-Serão, M.C.; Gutiérrez, N.A. A review of feed efficiency in swine: Biology and application. J. Anim. Sci. Biotechnol. 2015, 6, 33. [Google Scholar] [CrossRef]
- Hilgendorf, C.; Spahn-Langguth, H.; Regårdh, C.G.; Lipka, E.; Amidon, G.L.; Langguth, P. Caco-2 versus Caco-2/HT29-MTX co-cultured cell lines: Permeabilities via diffusion, inside- and outside-directed carrier-mediated transport. J. Pharm. Sci. 2000, 89, 63–75. [Google Scholar] [CrossRef]
- Jumarie, C.; Malo, C.M. Caco-2 cells cultured in serum-free medium as a model for the study of enterocytic differentiation in vitro. J. Cell. Physiol. 1991, 149, 24–33. [Google Scholar] [CrossRef]
- Klein, S.; Mueller, D.; Schevchenko, V.; Noor, F. Long-term maintenance of HepaRG cells in serum-free conditions and application in a repeated dose study. J. Appl. Toxicol. 2014, 34, 1078–1086. [Google Scholar] [CrossRef]
- van derValk, J.; Brunner, D.; DeSmet, K.; Svenningsen, Å.F.; Honegger, P.; Knudsen, L.E.; Lindl, T.; Noraberg, J.; Price, A.; Scarino, M.L.; et al. Optimization of chemically defined cell culture media—Replacing fetal bovine serum in mammalian in vitro methods. Toxicol. Vitr. 2010, 24, 1053–1063. [Google Scholar] [CrossRef]
- Chang, K.W.; Kuo, C.Y. 6-Gingerol modulates proinflammatory responses in dextran sodium sulfate (DSS)-treated Caco-2 cells and experimental colitis in mice through adenosine monophosphate-activated protein kinase (AMPK) activation. Food Funct. 2015, 6, 3334–3341. [Google Scholar] [CrossRef]
- Chen, S.; Einspanier, R.; Schoen, J. Transepithelial electrical resistance (TEER): A functional parameter to monitor the quality of oviduct epithelial cells cultured on filter supports. Histochem. Cell Biol. 2015, 144, 509–515. [Google Scholar] [CrossRef]
- Melgar, S.; Karlsson, A.; Michaëlsson, E. Acute colitis induced by dextran sulfate sodium progresses to chronicity in C57BL/6 but not in BALB/c mice: Correlation between symptoms and inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, 1328–1338. [Google Scholar] [CrossRef]
- Liu, X.C.; Mei, Q.; Xu, J.M.; Hu, J. Balsalazine decreases intestinal mucosal permeability of dextran sulfate sodium-induced colitis in mice. Acta Pharmacol. Sin. 2009, 30, 987–993. [Google Scholar] [CrossRef]
- Adnan, H.; Quach, H.; MacIntosh, K.; Antenos, M.; Kirby, G.M. Faecalibacterium prausnitzii supernatant improves intestinal barrier function in mice DSS colitis. Scand. J. Gastroenterol. 2013, 48, 1136–1144. [Google Scholar]
- Ueno, N.; Fujiya, M.; Segawa, S.; Nata, T.; Moriichi, K.; Tanabe, H.; Mizukami, Y.; Kobayashi, N.; Ito, K.; Kohgo, Y. Heat-killed body of lactobacillus brevis SBC8803 ameliorates intestinal injury in a murine model of colitis by enhancing the intestinal barrier function. Inflamm. Bowel Dis. 2011, 17, 2235–2250. [Google Scholar] [CrossRef]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef]
- Dowarah, R.; Verma, A.K.; Agarwal, N. The use of Lactobacillus as an alternative of antibiotic growth promoters in pigs: A review. Anim. Nutr. 2017, 3, 1–7. [Google Scholar] [CrossRef]
- Thu, T.V.; Loh, T.C.; Foo, H.L.; Yaakub, H.; Bejo, M.H. Effects of liquid metabolite combinations produced by Lactobacillus plantarum on growth performance, faeces characteristics, intestinal morphology and diarrhoea incidence in postweaning piglets. Trop. Anim. Health Prod. 2011, 43, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Blikslager, A.T.; Moeser, A.J.; Gookin, J.L.; Jones, S.L.; Odle, J. Restoration of barrier function in injured intestinal mucosa. Physiol. Rev. 2007, 87, 545–564. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Interleukin-18. Methods 1999, 19, 121–132. [Google Scholar] [CrossRef]
- Ganner, A.; Nitsch, S.; Erlacher, K.; Klimitsch, A.; Schatzmayr, G. Ex vivo effect of yeast beta-glucan on lymphocyte viability and plasma IL-18 in weaning piglets. Livest. Sci. 2010, 133, 246–248. [Google Scholar] [CrossRef]
- Tong, L.C.; Wang, Y.; BinWang, Z.; Liu, W.Y.; Sun, S.; Li, L.; Su, D.F.; Zhang, L.C. Propionate ameliorates dextran sodium sulfate-induced colitis by improving intestinal barrier function and reducing inflammation and oxidative stress. Front. Pharmacol. 2016, 7, 253. [Google Scholar] [CrossRef] [PubMed]
- Harmon, B.G. Avian Heterophils in Inflammation and Disease Resistance. Poult. Sci. 1998, 77, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Kansagra, K.; Stoll, B.; Rognerud, C.; Niinikoski, H.; Ou, C.N.; Harvey, R.; Burrin, D. Total parenteral nutrition adversely affects gut barrier function in neonatal piglets. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Young, D.; Ibuki, M.; Nakamori, T.; Fan, M.; Mine, Y. Soy-derived di-and tripeptides alleviate colon and ileum inflammation in pigs with dextran sodium sulfate-induced colitis. J. Nutr. 2012, 142, 363–368. [Google Scholar] [CrossRef]
- Rocha, B.S.; Correia, M.G.; Pereira, A.; Henriques, I.; DaSilva, G.J.; Laranjinha, J. Inorganic nitrate prevents the loss of tight junction proteins and modulates inflammatory events induced by broad-spectrum antibiotics: A role for intestinal microbiota? Nitric Oxide Biol. Chem. 2019, 88, 27–34. [Google Scholar] [CrossRef]
- Hackam, D.J.; Upperman, J.S.; Grishin, A.; Ford, H.R. Disordered enterocyte signaling and intestinal barrier dysfunction in the pathogenesis of necrotizing enterocolitis. Semin. Pediatr. Surg. 2005, 14, 49–57. [Google Scholar] [CrossRef]
- Wang, J.Q.; Yin, F.G.; Zhu, C.; Yu, H.; Niven, S.J.; de Lange, C.F.M.; Gong, J. Evaluation of probiotic bacteria for their effects on the growth performance and intestinal microbiota of newly-weaned pigs fed fermented high-moisture maize. Livest. Sci. 2012, 145, 79–86. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Antibiotic | P1 | P12 | G7 (L) | G7 (H) | ||
|---|---|---|---|---|---|---|---|
| Feed-to-gain ratio (F:G) | W1 | 0.78 ± 0.12 | 0.83 ± 0.08 | 0.64 ± 0.03 | 0.67 ± 0.05 | 0.70 ± 0.02 | 0.75 ± 0.05 |
| W2 | 0.65 ± 0.03 | 0.72 ± 0.01 | 0.65 ± 0.04 | 0.53 ± 0.16 | 0.64 ± 0.03 | 0.66 ± 0.05 | |
| W3 | 0.64 ± 0.01 a | 0.52 ± 0.01 b | 0.52 ± 0.04 b | 0.54 ± 0.02 b | 0.58 ± 0.03 ab | 0.59 ± 0.03 ab | |
| W4 | 0.60 ± 0.03 | 0.61 ± 0.02 | 0.60 ± 0.02 | 0.60 ± 0.01 | 0.60 ± 0.02 | 0.64 ± 0.02 | |
| W5 | 0.57 ± 0.02 | 0.53 ± 0.01 | 0.52 ± 0.01 | 0.55 ± 0.01 | 0.57 ± 0.03 | 0.53 ± 0.01 | |
| W6 | 0.49 ± 0.04 b | 0.63 ± 0.02 a | 0.55 ± 0.03 ab | 0.58 ± 0.05 ab | 0.62 ± 0.03 a | 0.57 ± 0.02 ab | |
| Feed conversion rate (FCR) | W1 | 1.36 ± 0.16 ab | 1.19 ± 0.13 b | 1.57 ± 0.06 a | 1.53 ± 0.12 ab | 1.42 ± 0.04 ab | 1.31 ± 0.10 ab |
| W2 | 1.55 ± 0.06 | 1.40 ± 0.02 | 1.56 ± 0.09 | 1.53 ± 0.09 | 1.57 ± 0.08 | 1.55 ± 0.12 | |
| W3 | 1.56 ± 0.04 b | 1.93 ± 0.03 a | 1.96 ± 0.14 a | 1.86 ± 0.07 a | 1.74 ± 0.09 ab | 1.72 ± 0.10 ab | |
| W4 | 1.67 ± 0.08 | 1.65 ± 0.07 | 1.68 ± 0.05 | 1.67 ± 0.04 | 1.68 ± 0.07 | 1.57 ± 0.06 | |
| W5 | 1.76 ± 0.06 | 1.91 ± 0.05 | 1.93 ± 0.05 | 1.81 ± 0.04 | 1.78 ± 0.08 | 1.88 ± 0.02 | |
| W6 | 2.10 ± 0.17 a | 1.57 ± 0.03 b | 1.84 ± 0.11 ab | 1.76 ± 0.15 ab | 1.62 ± 0.08 b | 1.76 ± 0.07 ab |
| Control | Antibiotic | P1 | P12 | G7 (L) | G7 (H) | ||
|---|---|---|---|---|---|---|---|
| Endotoxin (EU/mL) | Week 0 | 2.710 | 2.553 | 2.502 | 3.893 | 2.474 | 2.539 |
| Week 6 | 6.070 a | 1.715 b | 3.170 ab | 2.243 b | 1.974 b | 1.819 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abrehame, S.; Hung, M.-Y.; Chen, Y.-Y.; Liu, Y.-T.; Chen, Y.-T.; Liu, F.-C.; Lin, Y.-C.; Chen, Y.-P. Selection of Fermentation Supernatant from Probiotic Strains Exhibiting Intestinal Epithelial Barrier Protective Ability and Evaluation of Their Effects on Colitis Mouse and Weaned Piglet Models. Nutrients 2024, 16, 1138. https://doi.org/10.3390/nu16081138
Abrehame S, Hung M-Y, Chen Y-Y, Liu Y-T, Chen Y-T, Liu F-C, Lin Y-C, Chen Y-P. Selection of Fermentation Supernatant from Probiotic Strains Exhibiting Intestinal Epithelial Barrier Protective Ability and Evaluation of Their Effects on Colitis Mouse and Weaned Piglet Models. Nutrients. 2024; 16(8):1138. https://doi.org/10.3390/nu16081138
Chicago/Turabian StyleAbrehame, Solomon, Man-Yun Hung, Yu-Yi Chen, Yu-Tse Liu, Yung-Tsung Chen, Fang-Chueh Liu, Yu-Chun Lin, and Yen-Po Chen. 2024. "Selection of Fermentation Supernatant from Probiotic Strains Exhibiting Intestinal Epithelial Barrier Protective Ability and Evaluation of Their Effects on Colitis Mouse and Weaned Piglet Models" Nutrients 16, no. 8: 1138. https://doi.org/10.3390/nu16081138
APA StyleAbrehame, S., Hung, M. -Y., Chen, Y. -Y., Liu, Y. -T., Chen, Y. -T., Liu, F. -C., Lin, Y. -C., & Chen, Y. -P. (2024). Selection of Fermentation Supernatant from Probiotic Strains Exhibiting Intestinal Epithelial Barrier Protective Ability and Evaluation of Their Effects on Colitis Mouse and Weaned Piglet Models. Nutrients, 16(8), 1138. https://doi.org/10.3390/nu16081138