In Vitro Inhibition of Colorectal Cancer Gene Targets by Withania somnifera L. Methanolic Extracts: A Focus on Specific Genome Regulation

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
1.1. The Epidemiological Burden of Colorectal Cancer and Related Risk Indicators
1.2. An Overview of Targeted Genes in the Present Study
1.3. Phytotherapeutic Approaches to Treatment
1.4. The Botanical Description and Global Distribution of W. somnifera (L.)
2. Methodology
2.1. Procurement of Caco-2 Cell Lines
2.2. Plant Organ Acquisition
2.3. Extraction of WS Extracts Using Methanol and Acquisition of the Plant Organs
2.4. Reconstitution of Plant Extracts
2.5. Passaging Cancer Cell Lines (Caco-2)
2.6. Treatment of Cancer Cell Lines with WS Extracts
2.7. RNA Isolation
2.8. RNA Concentration and Purity Assessment Using UV Spectroscopy
2.9. Equipment and Procedure for a Quantitative Real-Time PCR (SYBR Green Protocol)
2.10. Analysis of Quantitative Real-Time PCR
2.11. Data Analysis
3. Results
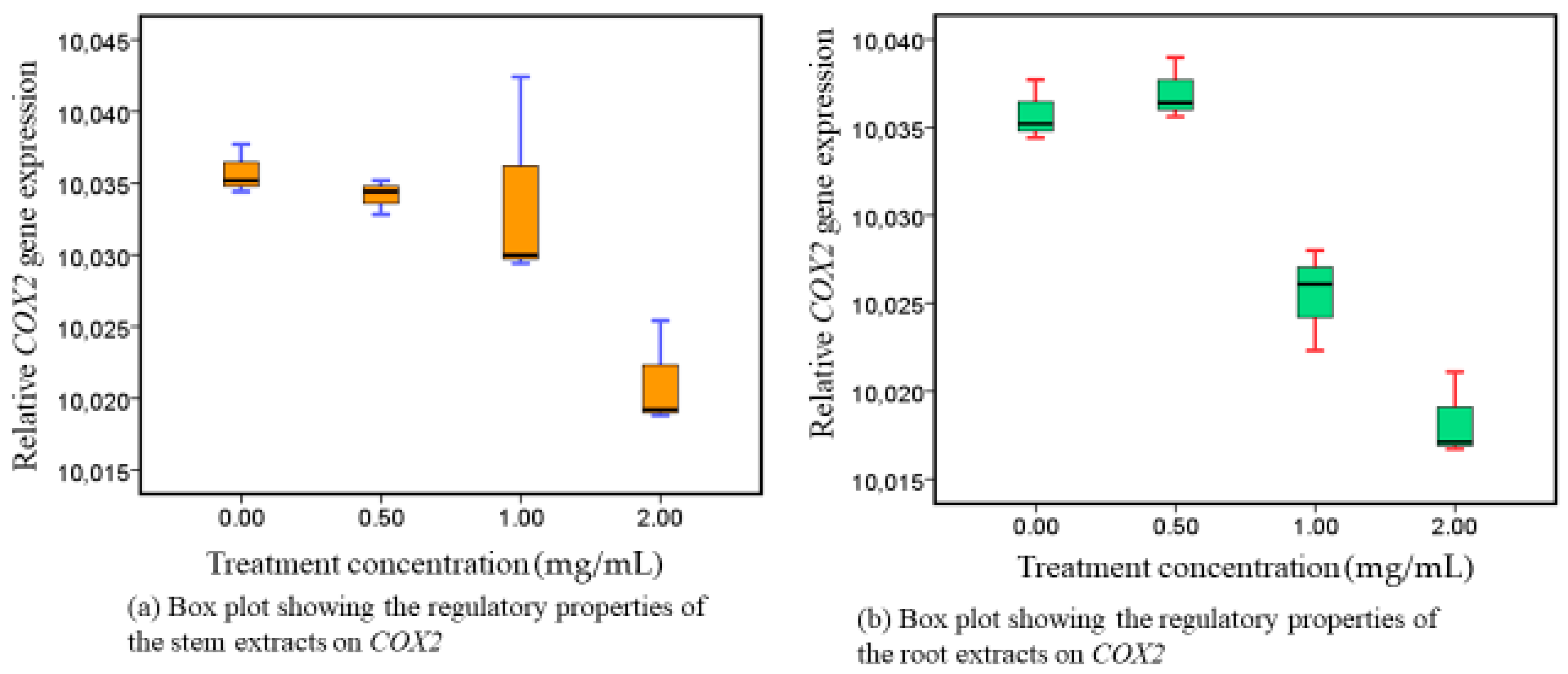
3.1. Responses of COX-2 following Administration of Methanolic Stem and Root Extracts at Progressive Dose Concentrations
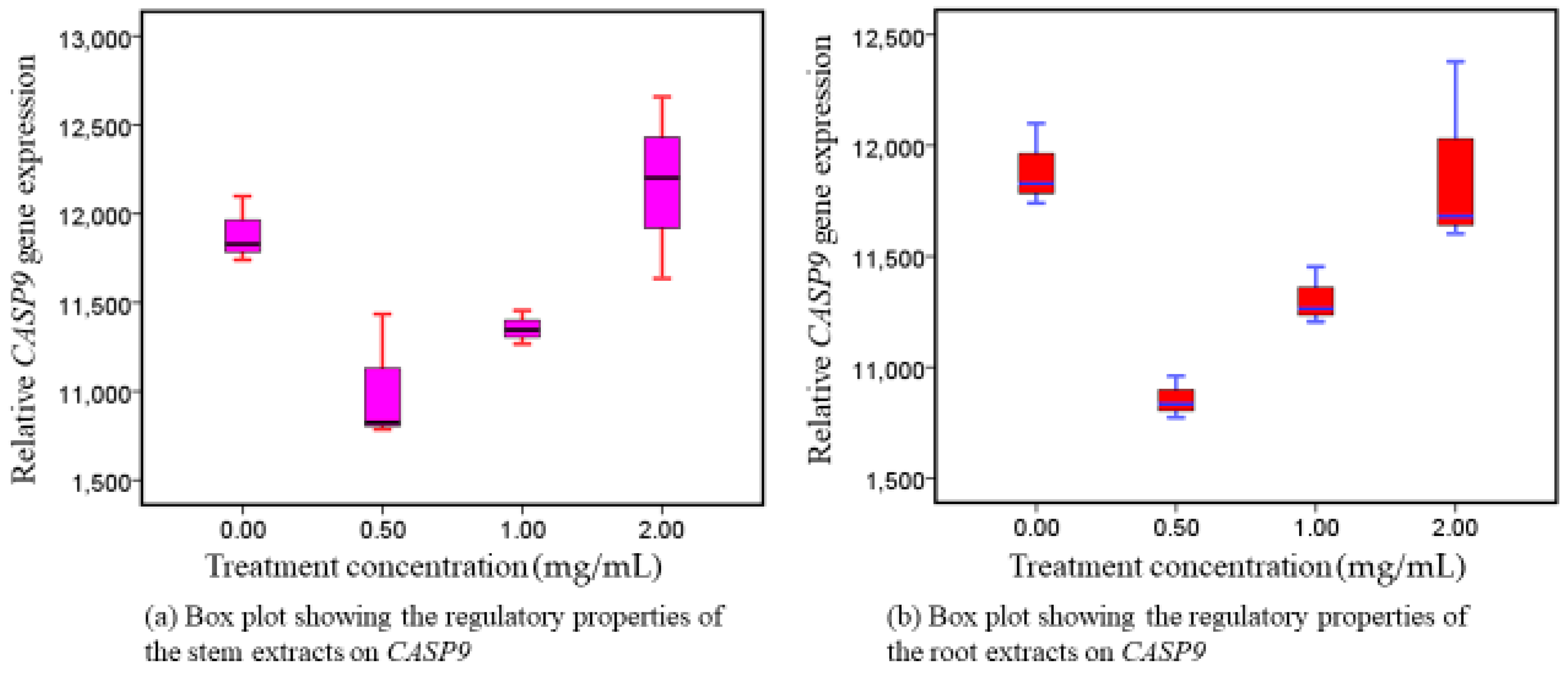
3.2. CASP9 Reactions following Exposure to Intervention Therapy
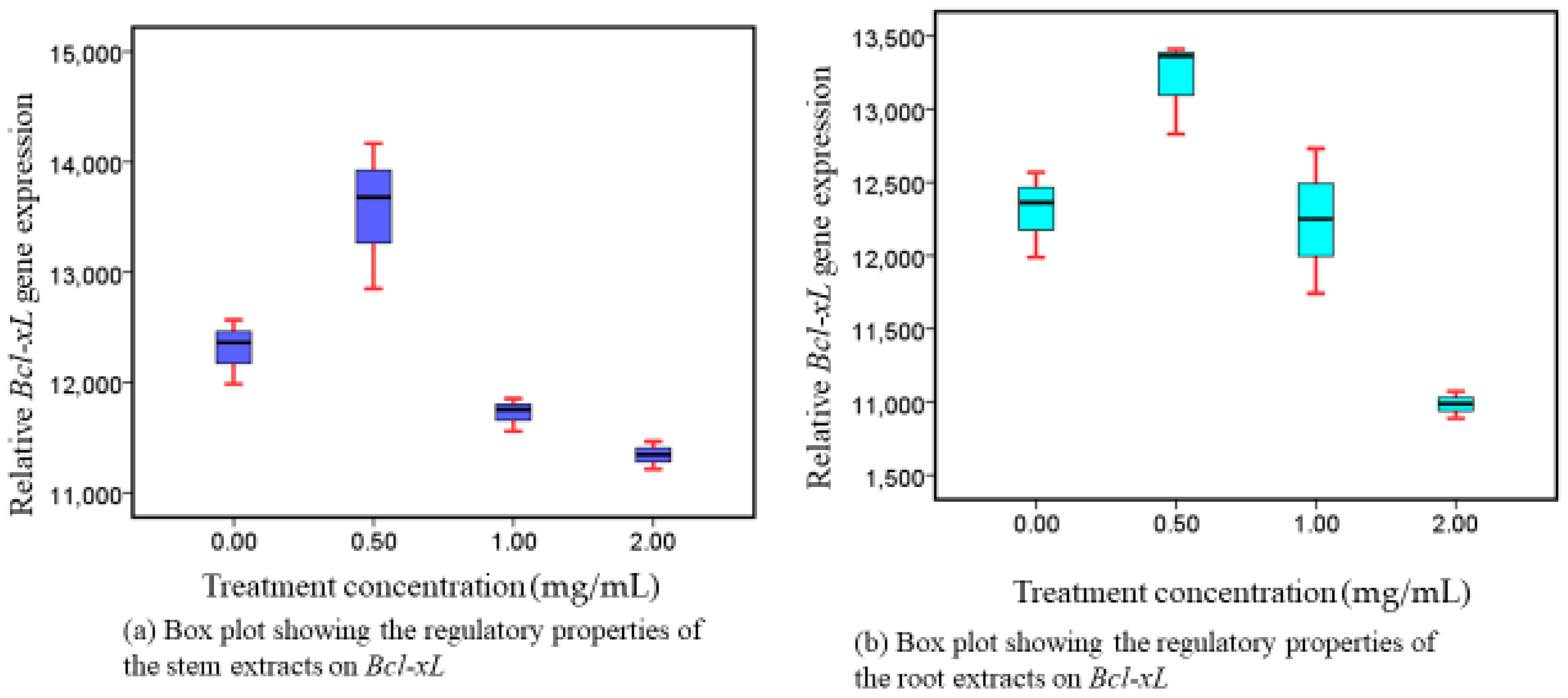
3.3. Responses of Bcl-xL following Administration of Methanolic Stem and Root Extracts at Progressive Dose Concentrations
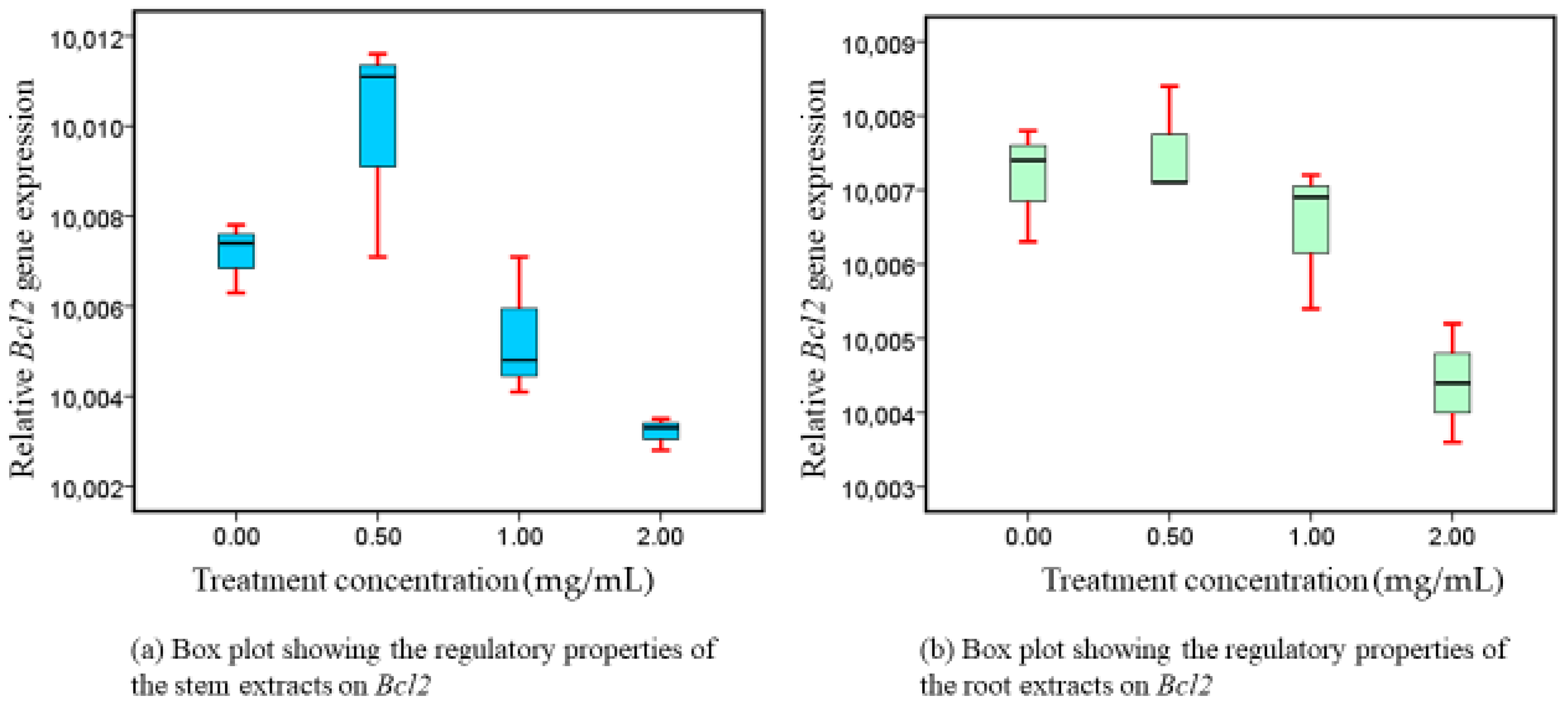
3.4. Bcl2 Expressions following Exposure to Intervention
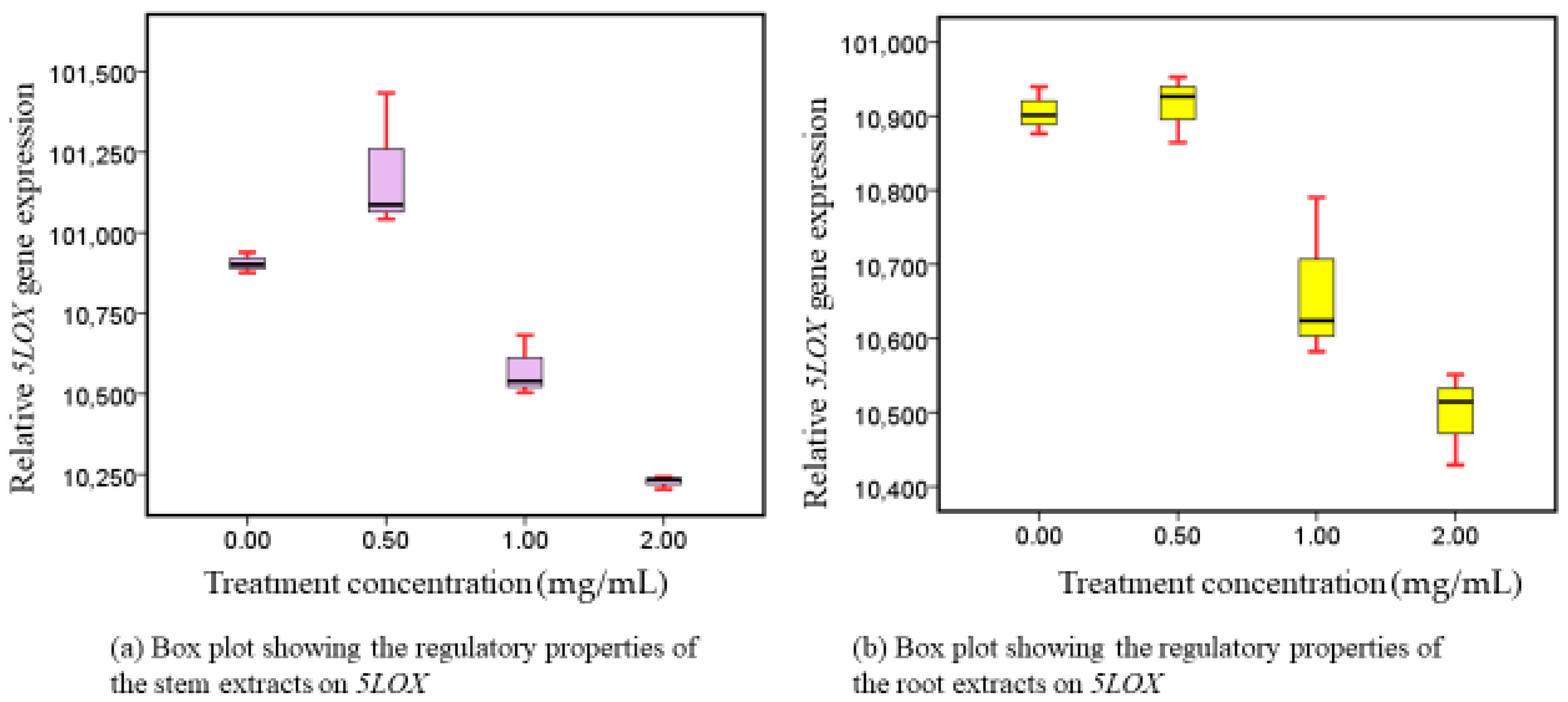
3.5. 5-LOX Responses following Administration of Methanolic Stem and Root Extracts at Progressive Dose Concentrations
4. Discussion
4.1. Phytotherapeutic Effects of Both (Roots and Stem) Extracts on Cyclooxigenase-2 Modulation
4.2. Phytotherapeutic Effects of Both Extracts on CASP9 Regulation
4.3. Phytotherapeutic Effects of Both Extracts on Bcl-xL and Bcl2 Regulation
4.4. Phytotherapeutic Effects of Both Extracts on 5-LOX Regulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef]
- Macharia, J.M.; Zhang, L.; Mwangi, R.W.; Rozmann, N.; Kaposztas, Z.; Varjas, T.; Sugár, M.; Alfatafta, H.; Pintér, M.; Bence, R.L. Are chemical compounds in medical mushrooms potent against colorectal cancer carcinogenesis and antimicrobial growth? Cancer Cell Int. 2022, 22, 379. [Google Scholar] [CrossRef]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global Cancer Statistics. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018, GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Macharia, J.M.; Mwangi, R.W.; Rozmann, N.; Wagara, I.N.; Kaposztas, Z.; Varjas, T.; Mathenge, J.; Bence, R.L. A systematic review of selected plants and their metabolites with anticolorectal cancer effects. Phytomed. Plus 2022, 2, 100332. [Google Scholar] [CrossRef]
- Macharia, J.M.; Mwangi, R.W.; Szabó, I.; Zand, A.; Kaposztas, Z.; Varjas, T.; Rozmann, N.; Raposa, B.L. Regulatory activities of Warbugia ugandensis ethanolic extracts on colorectal cancer-specific genome expression dose-dependently. Biomed. Pharmacother. 2023, 166, 115325. [Google Scholar] [CrossRef]
- Thornberry, N.A.; Lazebnik, Y. Caspases: Enemies within. Science 1998, 281, 1312–1316. [Google Scholar] [CrossRef]
- Raff, M. Cell suicide for beginners. Nature 1998, 396, 119–122. [Google Scholar] [CrossRef]
- Morimoto, Y.; Takada, K.; Takeuchi, O.; Watanabe, K.; Hirohara, M.; Hamamoto, T.; Masuda, Y. Bcl-2/Bcl-xL inhibitor navitoclax increases the antitumor effect of Chk1 inhibitor prexasertib by inducing apoptosis in pancreatic cancer cells via inhibition of Bcl-xL but not Bcl-2. Mol. Cell. Biochem. 2020, 472, 187–198. [Google Scholar] [CrossRef]
- Sjöström, J.; Bergh, J. How apoptosis is regulated, and what goes wrong in cancer. Br. Med. J. 2001, 322, 1538–1539. [Google Scholar] [CrossRef] [PubMed]
- Wong, B.C.Y.; Zhu, G.H.; Lam, S.K. Aspirin induced apoptosis in gastric cancer cells. Biomed. Pharmacother. 1999, 53, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Connelly, A.; Keku, T.O.; Mountcastle, S.B.; Galanko, J.; Woosley, J.T.; Schliebe, B.; Lund, P.; Sandler, R.S. Nonsteroidal anti-inflammatory drugs, apoptosis, and colorectal adenomas. Gastroenterology 2002, 123, 1770–1777. [Google Scholar] [CrossRef] [PubMed]
- Ezziyyani, M. Advanced Intelligent Systems for Sustainable Development (AI2SD’2019). Volume 2—Advanced Intelligent Systems for Sustainable Development Applied to Agriculture and Health; Advances in Intelligent Systems and Computing; Springer: Cham, Switzerland, 2019; Volume 1103. [Google Scholar]
- Pettersson, J.; Karlsson, P.C.; Göransson, U.; Rafter, J.J.; Bohlin, L. The flavouring phytochemical 2-pentanone reduces prostaglandin production and COX-2 expression in colon cancer cells. Biol. Pharm. Bull. 2008, 31, 534–537. [Google Scholar] [CrossRef] [PubMed]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 family proteins: Changing partners in the dance towards death. Cell Death Differ. 2018, 25, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-T.; Wolter, K.G.; Youle, R.J. Cytosol-to-membrane redistribution of Bax and Bcl-XL during apoptosis (programmed cell death Bcl-2 Bax Diphtheria toxin). Biochemistry 1997, 94, 3668–3672. [Google Scholar]
- Fiebig, A.A.; Zhu, W.; Hollerbach, C.; Leber, B.; Andrews, D.W. Bcl-XL is qualitatively different from and ten times more effective than Bcl-2 when expressed in a breast cancer cell line. BMC Cancer 2006, 6, 213. [Google Scholar] [CrossRef] [PubMed]
- Madhusudan, M.; Zameer, F.; Naidu, A.; Nagendra Prasad, M.N.; Dhananjaya, B.L.; Hegdekatte, R. Evaluating the inhibitory potential of Withania somnifera on platelet aggregation and inflammation enzymes: An in vitro and in silico study. Pharm. Biol. 2016, 54, 1936–1941. [Google Scholar] [CrossRef]
- Raghavendra, R.H.; Naidu, K.A. Eugenol and n-3 rich garden cress seed oil as modulators of platelet aggregation and eicosanoids in Wistar Albino rats. Open Nutraceuticals J. 2011, 4, 144–150. [Google Scholar] [CrossRef]
- Macharia, J.M.; Káposztás, Z.; Bence, R.L. Medicinal Characteristics of Withania somnifera L. in Colorectal Cancer Management. Pharmaceuticals 2023, 16, 915. [Google Scholar] [CrossRef]
- Macharia, J.M.; Varjas, T.; Mwangi, R.W.; Káposztás, Z.; Rozmann, N.; Pintér, M.; Wagara, I.N.; Raposa, B.L. Modulatory Properties of Aloe secundiflora’s Methanolic Extracts on Targeted Genes in Colorectal Cancer Management. Cancers 2023, 15, 5002. [Google Scholar] [CrossRef]
- Parmar, S.; Gangwal, A. The Antimicrobial activity of essential oil and plant extracts of Woodfordia fruticosa. Sch. Res. Libr. 2011, 2, 373–383. [Google Scholar]
- Mirjalili, M.H.; Moyano, E.; Bonfill, M.; Cusido, R.M.; Palazón, J. Steroidal lactones from Withania somnifera, an ancient plant for novel medicine. Molecules 2009, 14, 2373–2393. [Google Scholar] [CrossRef] [PubMed]
- Gaurav, H.; Yadav, D.; Maurya, A.; Yadav, H.; Yadav, R.; Shukla, A.C.; Sharma, M.; Gupta, V.K.; Palazon, J. Biodiversity, Biochemical Profiling, and Pharmaco-Commercial Applications of Withania somnifera: A Review. Molecules 2023, 28, 1208. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Maqsood, M.; Saeed, R.A.; Alam, A.; Sahar, A.; Kieliszek, M.; Miecznikowski, A.; Muzammil, H.S.; Aadil, R.M. Phytochemistry, food application, and therapeutic potential of the medicinal plant (Withania coagulans): A review. Molecules 2021, 26, 6881. [Google Scholar] [CrossRef] [PubMed]
- Rajeswara Rao, B.R.; Rajput, D.K.; Nagaraju, G.; Adinarayana, G. Opportunities and challenges in the cultivation of ashwagandha {Withania somnifera (L.) DUNAL}. J. Pharmacogn. 2012, 3, 88–91, ISSN: 0976-884X & E-ISSN: 0976-8858. [Google Scholar]
- Desai, A.; Qazi, G.; Ganju, R.; El-Tamer, M.; Singh, J.; Saxena, A.; Bedi, Y.; Taneja, S.; Bhat, H. Medicinal Plants and Cancer Chemoprevention. Curr. Drug Metab. 2008, 9, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Tewari, D.; Chander, V.; Dhyani, A.; Sahu, S.; Gupta, P.; Patni, P.; Kalick, L.S.; Bishayee, A. Withania somnifera (L.) Dunal: Phytochemistry, structure-activity relationship, and anticancer potential. Phytomedicine 2022, 98, 153949. [Google Scholar] [CrossRef] [PubMed]
- American Type Culture Collection. Caco-2 [Caco2]-HTB-37 | ATCC 2022. Available online: https://www.atcc.org/products/htb-37 (accessed on 15 February 2023).
- National Irrigation Authority. Perkerra Irrigation Scheme 2023. Available online: https://irrigation.go.ke/projects/perkerra-irrigation-scheme/ (accessed on 13 April 2022).
- Nelson, V.K.; Sahoo, N.K.; Sahu, M.; Sudhan, H.H.; Pullaiah, C.P.; Muralikrishna, K.S. In vitro anticancer activity of Eclipta alba whole plant extract on colon cancer cell HCT-116. BMC Complement. Med. Ther. 2020, 20, 1–8. [Google Scholar] [CrossRef]
- Fernand, V.E. Initial Characterization of Crude Extracts from Phyllanthus amarus Schum. and Thonn. and Quassia amara L. Using Normal Phase Thin Layer Chromatography. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2003. [Google Scholar]
- Chaves, J.O.; de Souza, M.C.; da Silva, L.C.; Lachos-Perez, D.; Torres-Mayanga, P.C.; Machado, A.P.D.F.; Forster-Carneiro, T.; Vázquez-Espinosa, M.; González-De-Peredo, A.V.; Barbero, G.F. Extraction of Flavonoids from Natural Sources Using Modern Techniques. Front. Chem. 2020, 8, 507887. [Google Scholar] [CrossRef]
- Toledo, A.G.; de Souza, J.G.D.L.; Santana, C.B.; Mallmann, A.P.; dos Santos, C.V.; Correa, J.M.; Pinto, F.G.D.S. Antimicrobial, antioxidant activity and phytochemical prospection of Eugenia involucrata DC. leaf extracts. Braz. J. Biol. 2023, 83, 1–9. [Google Scholar] [CrossRef]
- Tzanova, M.; Atanasov, V.; Yaneva, Z.; Ivanova, D.; Dinev, T. Selectivity of current extraction techniques for flavonoids from plant materials. Processes 2020, 8, 1222. [Google Scholar] [CrossRef]
- Tessema Desta, G.; Andargie Ferede, Y.; Sisay Zewdu, W.; Adela Alemu, M. Evaluation of Antidiarrheal Activity of 80% Methanol Extract and Solvent Fractions of the Leaves of Withania somnifera (L.) Dunal in Swiss Albino Mice. Evid.-Based Complement. Altern. Med. 2022, 2022, 7968973. [Google Scholar] [CrossRef] [PubMed]
- Duraisamy Saravanakumar, D.S.; Loganathan Karthiba, L.K.; Rajendran Ramjegathesh, R.R.; Kuppusami Prabakar, K.P.; Thiruvengadam Raguchander, T.R. Characterization of bioactive compounds from botanicals for the management of plant diseases. In Sustainable Crop Disease Management Using Natural Products; CABI: Wallingford, UK, 2015; pp. 1–18. [Google Scholar] [CrossRef]
- Sangweni, N.F.; Dludla, P.V.; Chellan, N.; Mabasa, L.; Sharma, J.R.; Johnson, R. The implication of low dose dimethyl sulfoxide on mitochondrial function and oxidative damage in cultured cardiac and cancer cells. Molecules 2021, 26, 7305. [Google Scholar] [CrossRef] [PubMed]
- Kus, M.; Ibragimow, I.; Piotrowska-Kempisty, H. Caco-2 Cell Line Standardization with Pharmaceutical Requirements and In Vitro Model Suitability for Permeability Assays. Pharmaceutics 2023, 15, 2523. [Google Scholar] [CrossRef]
- Doak, S.H.; Zaïr, Z.M. Real-time reverse-transcription polymerase chain reaction: Technical considerations for gene expression analysis. Methods Mol. Biol. 2012, 817, 251–270. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Soyoola, E.; Chanmugam, P.; Hart, S.; Sun, W.; Zhong, H.; Liou, S.; Simmons, D.; Hwang, D. Selective expression of mitogen-inducible cyclooxygenase in macrophages stimulated with lipopolysaccharide. J. Biol. Chem. 1992, 267, 25934–25938. [Google Scholar] [CrossRef] [PubMed]
- Macharia, J.M.; Mwangi, R.W.; Rozmann, N.; Zsolt, K.; Varjas, T.; Uchechukwu, P.O.; Wagara, I.N.; Raposa, B.L. Biomedicine & Pharmacotherapy Medicinal plants with anti-colorectal cancer bioactive compounds: Potential game-changers in colorectal cancer management. Biomed. Pharmacother. 2022, 153, 113383. [Google Scholar] [CrossRef]
- Jayaprakasam, B.; Zhang, Y.; Seeram, N.P.; Nair, M.G. Growth inhibition of human tumor cell lines by withanolides from Withania somnifera leaves. Life Sci. 2003, 74, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, B.; Pal, D.; Kolluru, V.; Tyagi, A.; Baby, B.; Dahiya, N.R.; Youssef, K.; Alatassi, H.; Ankem, M.K.; Sharma, A.K. The chemopreventive effect of withaferin A on spontaneous and inflammation-associated colon carcinogenesis models. Carcinogenesis 2018, 39, 1537–1547. [Google Scholar] [CrossRef]
- Jang, M.H.; Piao, X.L.; Kim, J.M.; Kwon, S.W.; Park, J.H. Inhibition of cholinesterase and amyloid-beta aggregation by resveratrol oligomers from Vitis amurensis. Phyther. Res. 2008, 22, 544–549. [Google Scholar] [CrossRef]
- Shen, X.G.; Wang, C.; Li, Y.; Wang, L.; Zhou, B.; Xu, B.; Jiang, X.; Zhou, Z.; Sun, X. Downregulation of caspase-9 is a frequent event in patients with stage II colorectal cancer and correlates with poor clinical outcome. Color. Dis. 2010, 12, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Zhou, Z.G.; Li, Y.; Wang, L.; Yang, L.; Zhou, B.; Liu, H.-Y.; Song, J.-M.; Zeng, Y.-J.; Wang, R.; et al. Clinicopathological significance of caspase-8 and caspase-10 expression in rectal cancer. Oncology 2008, 74, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, H.; Takada, Y.; Shishodia, S.; Jayaprakasam, B.; Nair, M.G.; Aggarwal, B.B. Withanolides potentiate apoptosis, inhibit invasion, and abolish osteoclastogenesis through suppression of nuclear factor-ΚB (NF-ΚB) activation and NF-ΚB-regulated gene expression. Mol. Cancer Ther. 2006, 5, 1434–1445. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Chakraborty, S.; Anand, U.; Dey, S.; Nandy, S.; Ghorai, M.; Saha, S.C.; Patil, M.T.; Kandimalla, R.; Proćków, J.; et al. Withania somnifera (L.) Dunal (Ashwagandha): A comprehensive review on ethnopharmacology, pharmacotherapeutics, biomedicinal and toxicological aspects. Biomed. Pharmacother. 2021, 143, 112175. [Google Scholar] [CrossRef] [PubMed]
- Dar, P.A.; Mir, S.A.; Bhat, J.A.; Hamid, A.; Singh, L.R.; Malik, F.; Dar, T.A. An anti-cancerous protein fraction from Withania somnifera induces ROS-dependent mitochondria-mediated apoptosis in human MDA-MB-231 breast cancer cells. Int. J. Biol. Macromol. 2019, 135, 77–87. [Google Scholar] [CrossRef]
- Das, T.; Roy, K.S.; Chakrabarti, T.; Mukhopadhyay, S.; Roychoudhury, S. Withaferin A modulates the Spindle Assembly Checkpoint by degradation of Mad2-Cdc20 complex in colorectal cancer cell lines. Biochem. Pharmacol. 2014, 91, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Koduru, S.; Kumar, R.; Srinivasan, S.; Evers, M.B.; Damodaran, C. Notch-1 inhibition by withaferin-A: A therapeutic target against colon carcinogenesis. Mol. Cancer Ther. 2010, 9, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Mofed, D.; Zekri, A.R.; El-Sayed, N.; Rahouma, M.; Sabet, S. Antioxidant activity and apoptotic induction as mechanisms of action of Withania somnifera (Ashwagandha) against a hepatocellular carcinoma cell line. J. Int. Med. Res. 2018, 46, 1358–1369. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The Bcl-2 apoptotic switch in cancer development and therapy. Oncogene 2007, 26, 1324–1337. [Google Scholar] [CrossRef]
- Shureiqi, I.; Jiang, W.; Zuo, X.; Wu, Y.; Stimmel, J.B.; Leesnitzer, L.M.; Morris, J.S.; Fan, H.-Z.; Fischer, S.M.; Lippman, S.M. The 15-lipoxygenase-1 product 13-S-hydroxyoctadecadienoic acid down-regulates PPAR-δ to induce apoptosis in colorectal cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 9968–9973. [Google Scholar] [CrossRef]
- Ye, Y.N.; Wu, W.K.K.; Shin, V.Y.; Bruce, I.C.; Wong, B.C.Y.; Cho, C.H. Dual inhibition of 5-LOX and COX-2 suppresses colon cancer formation promoted by cigarette smoke. Carcinogenesis 2005, 26, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Zhu, C.; Qiang, H.; Zhou, X.; Zhou, G. Enhancing antitumor effects in pancreatic cancer cells by combined use of COX-2 and 5-LOX inhibitors. Biomed. Pharmacother. 2011, 65, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Rådmark, O.; Shimizu, T.; Jörnvall, H.; Samuelsson, B. Leukotriene A4 hydrolase in human leukocytes. Purification and properties. J. Biol. Chem. 1984, 259, 12339–12345. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer ID | Forward Primer | Reverse Primer |
|---|---|---|
| Cyclooxygenase-2 | CGGTGAAACTCTGGCTAGACAG | GCAAACCGTAGATGCTCAGGGA |
| 5-Lipoxygenase | GGAGAACCTGTTCATCAACCGC | CAGGTCTTCCTGCCAGTGATTC |
| B-cell lymphoma 2 | ATCGCCCTGTGGATGACTGAGT | GCCAGGAGAAATCAAACAGAGGC |
| B-cell lymphoma-extra-large | GCCACTTACCTGAATGACCACC | AACCAGCGGTTGAAGCGTTCCT |
| Caspase-9 | GTTTGAGGACCTTCGACCAGCT | CAACGTACCAGGAGCCACTCTT |
| HPRT1 | TGCTTCTCCTCAGCTTCA | CTCAGGAGGAGGAAGCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macharia, J.M.; Pande, D.O.; Zand, A.; Budán, F.; Káposztás, Z.; Kövesdi, O.; Varjas, T.; Raposa, B.L. In Vitro Inhibition of Colorectal Cancer Gene Targets by Withania somnifera L. Methanolic Extracts: A Focus on Specific Genome Regulation. Nutrients 2024, 16, 1140. https://doi.org/10.3390/nu16081140
Macharia JM, Pande DO, Zand A, Budán F, Káposztás Z, Kövesdi O, Varjas T, Raposa BL. In Vitro Inhibition of Colorectal Cancer Gene Targets by Withania somnifera L. Methanolic Extracts: A Focus on Specific Genome Regulation. Nutrients. 2024; 16(8):1140. https://doi.org/10.3390/nu16081140
Chicago/Turabian StyleMacharia, John M., Daniel O. Pande, Afshin Zand, Ferenc Budán, Zsolt Káposztás, Orsolya Kövesdi, Tímea Varjas, and Bence L. Raposa. 2024. "In Vitro Inhibition of Colorectal Cancer Gene Targets by Withania somnifera L. Methanolic Extracts: A Focus on Specific Genome Regulation" Nutrients 16, no. 8: 1140. https://doi.org/10.3390/nu16081140
APA StyleMacharia, J. M., Pande, D. O., Zand, A., Budán, F., Káposztás, Z., Kövesdi, O., Varjas, T., & Raposa, B. L. (2024). In Vitro Inhibition of Colorectal Cancer Gene Targets by Withania somnifera L. Methanolic Extracts: A Focus on Specific Genome Regulation. Nutrients, 16(8), 1140. https://doi.org/10.3390/nu16081140