Bioactive Peptides from Muscle Sources: Meat and Fish


Abstract
:1. Introduction
History of Bioactive Peptide Discovery
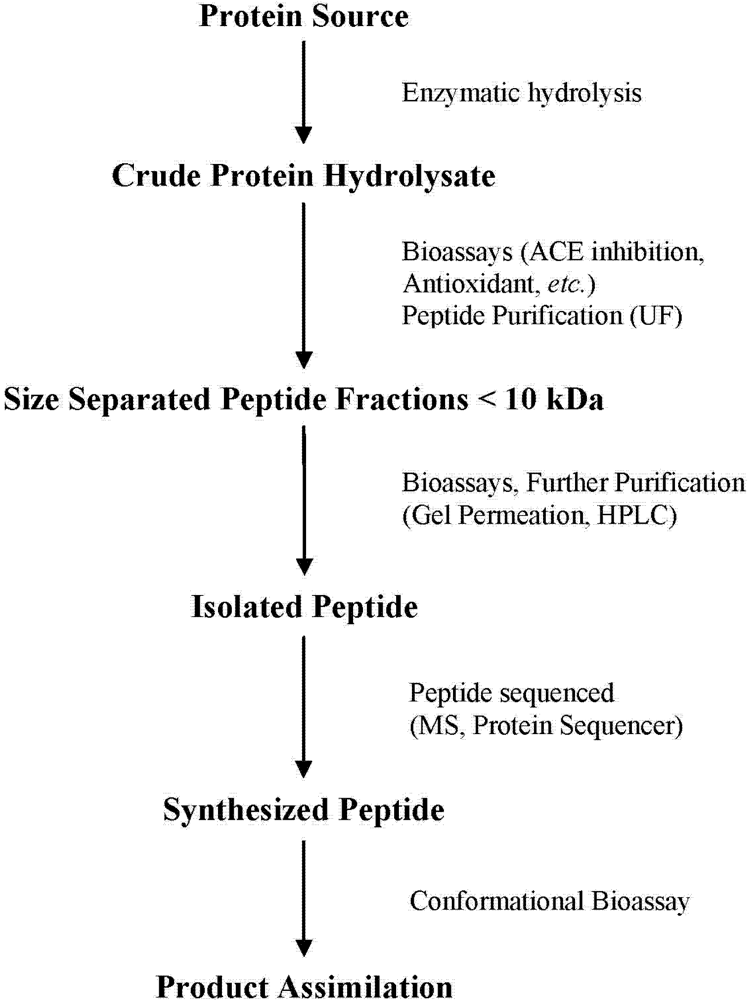
2. Production of Bioactive Peptides
Identification of Bioactive Peptides

3. Antihypertensive Peptides
3.1. Mechanism of ACE Inhibition
3.1.1. ACE Inhibitor Type Peptides
3.1.2. Absorption of ACE Inhibitory Peptides
3.1.3. Structure Correlation of ACE Inhibitory Peptides
3.2. Antihypertensive Peptides from Animal Sources
3.2.1. Antihypertensive Peptides from Myosin Sources
3.2.2. Antihypertensive Peptides from Troponin Sources
3.2.3. Antihypertensive Peptides from Collagen
3.2.4. Antihypertensive Peptides from Miscellaneous Sources

{kind=link}
| Source | Amino Acid Sequence | Parent Protein | Enzyme | IC50 (µM) | SHR a | Ref. |
|---|---|---|---|---|---|---|
| pig | ITTNP | myosin | thermolysin | 549 | ∆ 21.0 mmHg | [23,66] |
| MNPPK | myosin | synthesized | 945.5 | ∆ 23.4 mmHg | [23,66] | |
| MNP | myosin | synthesized | 66.6 | ∆ 19.6 mmHg | [23,66] | |
| PPL | myosin | synthesized | >1000 | ∆ 24.7 mmHg | [23,66] | |
| NPP | myosin | synthesized | 290.5 | ∆ 17.6 mmHg | [23,66] | |
| TNP | myosin | synthesized | 207.4 | ∆ 11.1 mmHg | [23,66] | |
| RMLGQTPTK | troponin | pepsin | 34 | nt | [50,85] | |
| RMLGQTP | troponin | pepsin | 503 | nt | [50] | |
| VKKVLGNP | myosin | pepsin | 28.5 | ∆ 24.0 mmHg b | [84] | |
| KRVITY | myosin | pepsin | 6.1 | ∆ 23.0 mmHg b | [83] | |
| VKAGF | actin | pepsin | 20.3 | ∆ 17.0 mmHg b | [83] | |
| KRQKYDI | troponin T | pepsin | 26.2 | ∆ 9.9 mmHg b | [24] | |
| KAPVA | titin | pepsin + pancreatin | 46.56 | nt | [86] | |
| PTPVP | titin | pepsin + pancreatin | 256.41 | nt | [86] | |
| ER | muscle | pepsin + pancreatin | 667 | nt | [86] | |
| KLP | muscle | pepsin + pancreatin | 500 | nt | [86] | |
| RPR | muscle | pepsin + pancreatin | 382 | nt | [86] | |
| chicken | LKA | creatine kinase | thermolysin | 8.5 | nt | [89] |
| LKP | aldolase | thermolysin | 0.32 | ∆ 75 mmHg c | [89] | |
| LAP | muscle | thermolysin | 3.2 | ∆ 40.0 mmHg c | [89] | |
| FQKPKR | myosin | thermolysin | 14 | nt | [89] | |
| FKGRYYP | creatine kinase | thermolysin | 0.55 | ∆ 0 mmHg c | [89] | |
| IKW | muscle | thermolysin | 0.21 | ∆ 50.0 mmHg c | [89] | |
| GFXGTXGLXGF | muscle | Aspergillus protease + gastric proteases | 42.4 | ∆ 20 mmHg d | [36,79] | |
| GAXGLXGP | collagen | Aspergillus protease + protease FP, A, G, N | 29.4 | nt | [37] | |
| GAXGPAGPGGIXGERGLXG | collagen | Aspergillus protease + protease FP, A, G, N | 45.6 | nt | [37] | |
| GLXGSRGERGERGLXG | collagen | Aspergillus protease + protease FP, A, G, N | 60.8 | nt | [37] | |
| GIXGSRGERGPVGPSG | collagen | Aspergillus protease + protease FP, A, G, N | 43.4 | nt | [37] | |
| cow | VLAQYK | muscle | thermolysin + proteinase A | 32.06 | + | [25,88] |
| GFHI | muscle | proteinase K | 64.3 | nt | [10] | |
| DFHING | muscle | alcalase | 50.5 | nt | [10] | |
| FHG | muscle | thermolysin + proteinase A | 52.9 | nt | [10] | |
| GLSDGEWQ | muscle | thermolysin + proteinase A | 117 | nt | [10] | |
| GPV | skin gelatin | alcalase + pronase E + collagenase | 4.67 | nt | [34] | |
| GPL | skin gelatin | alcalase + pronase E + collagenase | 2.55 | nt | [34] |
4. Antihypertensive Peptides from Fish Sources
Antihypertensive Peptides from Fish By-Products
| Source | Amino acid sequence | Parent protein | Enzyme | IC50 (µM) | SHR a | Ref. |
|---|---|---|---|---|---|---|
| bonito | IKPLNY | muscle | thermolysin | 43 | nt | [75] |
| IVGRPRHQG | actin | thermolysin | 2.4 | nt | [75] | |
| IWHHT | actin | thermolysin | 5.8 | nt | [75] | |
| ALPHA | actin | thermolysin | 10 | nt | [75] | |
| FQP | actin | thermolysin | 12 | nt | [75] | |
| LKPNM | muscle | thermolysin | 2.4 | ∆ 23.0 mmHg b | [72,75] | |
| IY | actin | thermolysin | 2.31 | ∆ 19.0 mmHg b | [72,75] | |
| DYGLYP | muscle | thermolysin | 62 | nt | [75] | |
| LKP | muscle | thermolysin | 0.32 | ∆ 18.0 mmHg b | [72] | |
| IWHHT | actin | thermolysin | 3.5 | ∆ 26.0 mmHg b | [72] | |
| IKP | muscle | thermolysin | 6.9 | ∆ 20.0 mmHg b | [72] | |
| IVGRPR | actin | thermolysin | 300 | ∆ 25.0 mmHg b | [89] | |
| salmon | WA | muscle | thermolysin | 277.3 | nt | [74] |
| VW | muscle | thermolysin | 2.5 | nt | [74] | |
| WM | muscle | thermolysin | 96.6 | nt | [74] | |
| MW | muscle | thermolysin | 9.9 | nt | [74] | |
| IW | muscle | thermolysin | 4.7 | nt | [74] | |
| LW | muscle | thermolysin | 17.4 | nt | [74] | |
| sardine | MF | muscle | alcalase | 44.7 | nt | [73] |
| RY | muscle | alcalase | 51 | nt | [73] | |
| MY | muscle | alcalase | 193 | nt | [73] | |
| LY | muscle | alcalase | 38.5 | nt | [73] | |
| YL | muscle | alcalase | 82 | nt | [73] | |
| IY | muscle | alcalase | 10.5 | nt | [73] | |
| VF | muscle | alcalase | 43.7 | nt | [73] | |
| GRP | muscle | alcalase | 20 | nt | [73] | |
| RFP | muscle | alcalase | 330 | nt | [73] | |
| AKK | muscle | alcalase | 3.13 | nt | [73] | |
| RVY | muscle | alcalase | 205.6 | nt | [73] | |
| GWAP | muscle | alcalase | 3.86 | nt | [73] | |
| KY | muscle | alcalase | 1.63 | nt | [73] | |
| VY | muscle | alcalase | 10 | ∆ 7.0 mmHg | [99,100] | |
| tuna | PTHIKWGD | muscle | acid | nd | nt | [101] |
| GDLGKTTTVSNWSPPKYKDTP | frame protein | pepsin | 11.28 | ∆ 21.0 mmHg | [51] | |
| WPEAAELMMEVDP | dark muscle | pepsin | 21.6 | ∆ 18.0 mmHg | [49] | |
| alaska pollack | GPL | skin | alcalase + pronase + collagenase | 2.65 | nt | [102] |
| GPM | skin | alcalase + pronase + collagenase | 17.13 | nt | [102] | |
| FGASTRGA | frame protein | pepsin | 14.7 | nt | [95] | |
| yellowfin sole | MIFPGAGGPEL | frame protein | α chymotrypsin | 28.7 | ∆ 22.0 mmHg | [96] |
| shark | EY | muscle | protease SM98011 | 1.98 | nt | [98] |
| FE | muscle | protease SM98012 | 2.68 | nt | [98] | |
| CF | muscle | protease SM98013 | 1.45 | nt | [98] |
5. Antioxidant Peptides
5.1. Antioxidant Peptides from Fish Sources
5.2. Antioxidant Peptides from Meat Sources
6. Antimicrobial and Antiproliferative Peptides
7. Conclusion
Acknowledgements
References
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar]
- Friedman, M. Nutritional value of proteins from different food sources: A review. J. Agric. Food Chem. 1996, 44, 6–29. [Google Scholar]
- Vercruysse, L.; van Camp, J.; Smagghe, G. ACE inhibitory peptides derived from enzymatic hydrolysates of animal muscle protein: A review. J. Agric. Food Chem. 2005, 53, 8106–8115. [Google Scholar]
- Vermeirssen, V.; Camp, J.V.; Verstraete, W. Bioavailability of angiotensin I converting enzyme inhibitory peptides. Br. J. Nutr. 2007, 92, 357–366. [Google Scholar]
- Erdmann, K.; Cheung, B.W.Y.; Schroder, H. The possible roles of food-derived bioactive peptides in reducing the risk of cardiovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar]
- Arihara, K. Strategies for designing novel functional meat products. Meat Sci. 2006, 74, 219–229. [Google Scholar]
- Kim, E.K.; Lee, S.J.; Jeon, B.T.; Moon, S.H.; Kim, B.; Park, T.K.; Han, J.S.; Park, P.J. Purification and characterisation of antioxidative peptides from enzymatic hydrolysates of venison protein. Food Chem. 2009, 114, 1365–1370. [Google Scholar]
- Matsui, T.; Matsumoto, K.; Mahmud, T.H.K.; Arjumand, A. Antihypertensive peptides from natural resources. In Advances in Phytomedicine; Elsevier: Oxford, UK, 2006; Volume 2, pp. 255–271. [Google Scholar]
- Cinq-Mars, C.D.; Hu, C.; Kitts, D.D.; Li-Chan, E.C.Y. Investigations into inhibitor type and mode, simulated gastrointestinal digestion, and cell transport of the angiotensin I-Converting enzyme-inhibitory peptides in pacific hake (Merluccius productus) fillet hydrolysate. J. Agric. Food Chem. 2008, 56, 410–419. [Google Scholar] [PubMed]
- Jang, A.; Jo, C.; Kang, K.S.; Lee, M. Antimicrobial and human cancer cell cytotoxic effect of synthetic angiotensin-converting enzyme (ACE) inhibitory peptides. Food Chem. 2008, 107, 327–336. [Google Scholar]
- Mellander, O. The physiological importance of the casein phosphopeptide calcium salts. II. Peroral calcium dosage of infants. Acta Soc. Med. Ups. 1950, 55, 247–255. [Google Scholar] [PubMed]
- Rutherfurd-Markwick, K.J.; Moughan, P.J. Bioactive peptides derived from food. J. AOAC Int. 2005, 88, 955–966. [Google Scholar]
- Hayes, M.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Putting microbes to work: Dairy fermentation, cell factories and bioactive peptides. Part I: Overview. J. Biotechnol. 2007, 2, 426–434. [Google Scholar]
- Korhonen, H. Milk-derived bioactive peptides: From science to applications. J. Funct. Foods 2009, 1, 177–187. [Google Scholar]
- Hernandez-Ledesma, B.; Quiros, A.; Amigo, L.; Recio, I. Identification of bioactive peptides after digestion of human milk and infant formula with pepsin and pancreatin. Int. Dairy J. 2007, 17, 42–49. [Google Scholar]
- Zhu, K.X.; Zhou, H.M.; Qian, H.F. Antioxidant and free radical-scavenging activities of wheat germ protein hydrolysates (WGPH) prepared with alcalase. Process Biochem. 2006, 41, 1296–1302. [Google Scholar]
- You, L.J.; Zhao, M.M.; Cui, C.; Zhao, H.F.; Yang, B. Effect of degree of hydrolysis on the antioxidant activity of loach (Misgurnus anguillicaudatus) protein hydrolysates. Inn. Food Sci. Emerg. Technol. 2009, 10, 235–240. [Google Scholar]
- Rizzello, C.G.; Losito, I.; Gobbetti, M.; Carbonara, T.; de Bari, M.D.; Zambonin, P.G. Antibacterial activities of peptides from the water-soluble extracts of Italian cheese varieties. J. Dairy Sci. 2005, 88, 2348–2360. [Google Scholar]
- Hayes, M.; Ross, R.P.; Fitzgerald, G.F.; Hill, C.; Stanton, C. Casein-derived antimicrobial peptides generated by Lactobacillus acidophilus DPC6026. Appl. Environ. Microbiol. 2006, 72, 2260–2264. [Google Scholar] [PubMed]
- Tsai, J.S.; Chen, T.J.; Pan, B.S.; Gong, S.D.; Chung, M.Y. Antihypertensive effect of bioactive peptides produced by protease-facilitated lactic acid fermentation of milk. Food Chem. 2008, 106, 552–558. [Google Scholar]
- Li, G.H.; Le, G.W.; Shi, Y.H.; Shrestha, S. Angiotensin I-converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar]
- Pihlanto, A.; Akkanen, S.; Korhonen, H.J. ACE-inhibitory and antioxidant properties of potato (Solanum tuberosum). Food Chem. 2008, 109, 104–112. [Google Scholar] [CrossRef]
- Arihara, K.; Nakashima, Y.; Mukai, T.; Ishikawa, S.; Itoh, M. Peptide inhibitors for angiotensin I-converting enzyme from enzymatic hydrolysates of porcine skeletal muscle proteins. Meat Sci. 2001, 57, 319–324. [Google Scholar]
- Katayama, K.; Anggraeni, H.E.; Mori, T.; Ahhmed, A.A.; Kawahara, S.; Sugiyama, M.; Nakayama, T.; Maruyama, M.; Mugurumat, M. Porcine skeletal muscle troponin is a good source of peptides with angiotensin-I converting enzyme inhibitory activity and anti hypertensive effects in spontaneously hypertensive rats. J. Agric. Food Chem. 2008, 56, 355–360. [Google Scholar]
- Jang, A.; Lee, M. Purification and identification of angiotensin converting enzyme inhibitory peptides from beef hydrolysates. Meat Sci. 2005, 69, 653–661. [Google Scholar]
- Liu, Z.Y.; Dong, S.Y.; Xu, J.; Zeng, M.Y.; Song, H.X.; Zhao, Y.H. Production of cysteine-rich antimicrobial peptide by digestion of oyster (Crassostrea gigas) with alcalase and bromelin. Food Control 2008, 19, 231–235. [Google Scholar] [CrossRef]
- Hayes, M.; Stanton, C.; Slattery, H.; O’Sullivan, O.; Hill, C.; Fitzgerald, G.F.; Ross, R.P. Casein fermentate of Lactobacillus animalis DPC6134 contains a range of novel propeptide angiotensin-converting enzyme inhibitors. Appl. Environ. Microbiol. 2007, 73, 4658–4667. [Google Scholar]
- Kudoh, Y.; Matsuda, S.; Igoshi, K.; Oki, T. Antioxidative peptide from milk fermented with Lactobacillus delbrueckii subsp. bulgaricus IFO13953. J. Jpn. Soc. Food Sci. Technol. 2001, 48, 44–50. [Google Scholar] [CrossRef]
- Pihlanto, A.; Virtanen, T.; Korhonen, H. Angiotensin I converting enzyme (ACE) inhibitory activity and antihypertensive effect of fermented milk. Int. Dairy J. 2010, 20, 3–10. [Google Scholar]
- Arihara, K.; Ohata, M. Functional Properties of Bioactive Peptides Derived from meat Proteins. In Advanced Technologies for Meat Processing; Toldra, F., Ed.; Springer: New York, NY, USA, 2006; pp. 245–274. [Google Scholar]
- Hammes, W.P.; Haller, D.; Ganzle, M.G. Fermented Meat. In Handbook of Fermented Functional Foods; Farnworth, E.R., Ed.; CRC: New York, NY, USA, 2003; pp. 251–269. [Google Scholar]
- Ondetti, M.A.; Rubin, B.; Cushman, D.W. Design of specific inhibitors of angiotensin-converting enzyme: new class of orally active antihypertensive agents. Science 1977, 196, 441–444. [Google Scholar]
- Iroyukifujita, H.; Eiichiyokoyama, K.; Yoshikawa, M. Classification and antihypertensive activity of angiotensin I-converting enzyme inhibitory peptides derived from food proteins. J. Food Sci. 2000, 65, 564–569. [Google Scholar]
- Kim, S.-K.; Byun, H.-G.; Park, P.-J.; Shahidi, F. Angiotensin I converting enzyme inhibitory peptides purified from bovine skin gelatin hydrolysate. J. Agric. Food Chem. 2001, 49, 2992–2997. [Google Scholar]
- Qian, Z.J.; Jung, W.K.; Lee, S.H.; Byun, H.G.; Kim, S.K. Antihypertensive effect of an angiotensin I-converting enzyme inhibitory peptide from bullfrog (Rana catesbeiana Shaw) muscle protein in spontaneously hypertensive rats. Process Biochem. 2007, 42, 1443–1448. [Google Scholar]
- Saiga, A.; Okumura, T.; Makihara, T.; Katsuda, S.I.; Morimatsu, F.; Nishimura, T. Action mechanism of an angiotensin I-converting enzyme inhibitory peptide derived from chicken breast muscle. J. Agric. Food Chem. 2006, 54, 942–945. [Google Scholar]
- Saiga, A.; Iwai, K.; Hayakawa, T.; Takahata, Y.; Kitamura, S.; Nishimura, T.; Morimatsu, F. Angiotensin I-converting enzyme-inhibitory peptides obtained from chicken collagen hydrolysate. J. Agric. Food Chem. 2008, 56, 9586–9591. [Google Scholar]
- Ferreira, S.H.; Bartelt, D.C.; Greene, L.J. Isolation of bradykinin-potentiating peptides from Bothrops jararaca venom. Biochemistry 1970, 9, 2583–2593. [Google Scholar]
- Vermeirssen, V.; van Camp, J.; Verstraete, W. Optimisation and validation of an angiotensin-converting enzyme inhibition assay for the screening of bioactive peptides. J. Biochem. Biophys. Methods 2002, 51, 75–87. [Google Scholar]
- Shalaby, S.M.; Zakora, M.; Otte, J. Performance of two commonly used angiotensin-converting enzyme inhibition assays using FA-PGG and HHL as substrates. J. Dairy Res. 2006, 73, 178–186. [Google Scholar]
- Lopez-Fandino, R.; Otte, J.; van Camp, J. Physiological, chemical and technological aspects of milk-protein-derived peptides with antihypertensive and ACE-inhibitory activity. Int. Dairy J. 2006, 16, 1277–1293. [Google Scholar]
- Brown, N.J.; Vaughan, D.E. Angiotensin-converting enzyme inhibitors. Circulation 1998, 97, 1411–1420. [Google Scholar]
- Folkow, B.; Johansson, B.; Mellander, S. The comparative effects of angiotensin and noradrenaline on consecutive vascular sections. Acta Physiol. Scand. 1961, 53, 99–104. [Google Scholar]
- Biron, P.; Koiw, E.; Nowaczynski, W.; Brouillet, J.; Genest, J. The effects of intravenous infusions of valine-5 angiotensin II and other pressor agents on urinary electrolytes and corticosteroids, including aldosterone. J. Clin. Invest. 1961, 40, 338–347. [Google Scholar]
- Padfield, P.L.; Morton, J.J. Effects of angiotensin II on arginine-vasopressin in physiological and pathological situations in man. J. Endocrinol. 1977, 74, 251–259. [Google Scholar]
- Proud, D.; Kaplan, A.P. Kinin formation: Mechanisms and role in inflammatory disorders. Annu. Rev. Immunol. 1988, 6, 49–83. [Google Scholar]
- Vanhoutte, P.M. Endothelium and control of vascular function. State of the art lecture. Hypertension 1989, 13, 658–667. [Google Scholar] [PubMed]
- Ahhmed, A.M.; Muguruma, M. A review of meat protein hydrolysates and hypertension. Meat Sci. 2010, 86, 110–118. [Google Scholar]
- Qian, Z.-J.; Je, J.-Y.; Kim, S.-K. Antihypertensive effect of angiotensin I converting enzyme-inhibitory peptide from hydrolysates of bigeye tuna dark muscle, thunnus obesus. J. Agric. Food Chem. 2007, 55, 8398–8403. [Google Scholar]
- Katayama, K.; Tomatsu, M.; Kawahara, S.; Yamauchi, K.; Fuchu, H.; Kodama, Y.; Kawamura, Y.; Muguruma, M. Inhibitory profile of nonapeptide derived from porcine troponin C against angiotensin I-converting enzyme. J. Agric. Food Chem. 2004, 52, 771–775. [Google Scholar]
- Lee, S.-H.; Qian, Z.-J.; Kim, S.-K. A novel angiotensin I converting enzyme inhibitory peptide from tuna frame protein hydrolysate and its antihypertensive effect in spontaneously hypertensive rats. Food Chem. 2010, 118, 96–102. [Google Scholar]
- Wijesekara, I.; Kim, S.K. Angiotensin-I-Converting Enzyme (ACE) Inhibitors from Marine Resources: Prospects in the Pharmaceutical Industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar]
- Fuller, M.F.; Tomé, D. In vivo determination of amino acid bioavailability in humans and model animals. J. AOAC Int. 2005, 88, 923–934. [Google Scholar] [PubMed]
- Balimane, P.V.; Chong, S.; Morrison, R.A. Current methodologies used for evaluation of intestinal permeability and absorption. J. Pharmacol. Toxicol. Methods 2000, 44, 301–312. [Google Scholar]
- Ekmekcioglu, C. A physiological approach for preparing and conducting intestinal bioavailability studies using experimental systems. Food Chem. 2002, 76, 225–230. [Google Scholar]
- Pauletti, G.M.; Gangwar, S.; Siahaan, T.J.; Jeffrey, A.; Borchardt, R.T. Improvement of oral peptide bioavailability: Peptidomimetics and prodrug strategies. Adv. Drug Deliv. Rev. 1997, 27, 235–256. [Google Scholar]
- DeSesso, J.M.; Jacobson, C.F. Anatomical and physiological parameters affecting gastrointestinal absorption in humans and rats. Food Chem. Toxicol. 2001, 39, 209–228. [Google Scholar]
- Greger, J.L. Using animals to assess bioavailability of minerals: Implications for human nutrition. J. Nutr. 1992, 122, 2047–2052. [Google Scholar]
- Foltz, M.; Cerstiaens, A.; van Meensel, A.; Mols, R.; van der Pijl, P.C.; Duchateau, G.S.M.J.E.; Augustijns, P. The angiotensin converting enzyme inhibitory tripeptides Ile-Pro-Pro and Val-Pro-Pro show increasing permeabilities with increasing physiological relevance of absorption models. Peptides 2008, 29, 1312–1320. [Google Scholar]
- Sun, H.; Liu, D.; Li, S.; Qin, Z. Transepithelial transport characteristics of the antihypertensive peptide, Lys-Val-Leu-Pro-Val-Pro, in human intestinal Caco-2 cell monolayer. Biosci. Biotechnol. Biochem. 2009, 73, 293–298. [Google Scholar]
- Vermeirssen, V.; Camp, J.V.; Verstraete, W. Bioavailability of angiotensin I converting enzyme inhibitory peptides. Br. J. Nutr. 2004, 92, 357–366. [Google Scholar]
- Cinq-Mars, C.D.; Hu, C.; Kitts, D.D.; Li-Chan, E.C.Y. Investigations into inhibitor type and mode, simulated gastrointestinal digestion, and cell transport of the angiotensin I-converting enzyme-inhibitory peptides in pacific hake (Merluccius productus) fillet hydrolysate. J. Agric. Food Chem. 2007, 56, 410–419. [Google Scholar]
- Bailey, C.A.; Bryla, P.; Malick, A.W. The use of the intestinal epithelial cell culture model, Caco-2, in pharmaceutical development. Adv. Drug Deliv. Rev. 1996, 22, 85–103. [Google Scholar] [CrossRef]
- Hidalgo, I.J.; Raub, T.J.; Borchardt, R.T. Characterization of the human colon carcinoma cell line (Caco-2) as a model system for intestinal epithelial permeability. Gastroenterology 1989, 96, 736–749. [Google Scholar]
- Fujita, H.; Yokoyama, K.; Yasumoto, R.; Yoshikawa, M. Antihypertensive effect of thermolysin digest of dried bonito in spontaneously hypertensive rat. Clin. Exp. Pharmacol. Physiol. 1995, 22, S304–S305. [Google Scholar]
- Nakashima, Y.; Arihara, K.; Sasaki, A.; Mio, H.; Ishikawa, S.; Itoh, M. Antihypertensive activities of peptides derived from porcine skeletal muscle myosin in spontaneously hypertensive rats. J. Food Sci. 2002, 67, 434–437. [Google Scholar]
- Quirós, A.; Dávalos, A.; Lasunción, M.A.; Ramos, M.; Recio, I. Bioavailability of the antihypertensive peptide LHLPLP: Transepithelial flux of HLPLP. Int. Dairy J. 2008, 18, 279–286. [Google Scholar]
- Foltz, M.; Meynen, E.E.; Bianco, V.; van Platerink, C.; Koning, T.M.M.G.; Kloek, J. Angiotensin converting enzyme inhibitory peptides from a lactotripeptide-enriched milk beverage are absorbed intact into the circulation. J. Nutr. 2007, 137, 953–958. [Google Scholar]
- Hata, Y.; Yamamoto, M.; Ohni, M.; Nakajima, K.; Nakamura, Y.; Takano, T. A placebo-controlled study of the effect of sour milk on blood pressure in hypertensive subjects. Am. J. Clin. Nutr. 1996, 64, 767–771. [Google Scholar]
- Seppo, L.; Jauhiainen, T.; Poussa, T.; Korpela, R. A fermented milk high in bioactive peptides has a blood pressure-lowering effect in hypertensive subjects. Am. J. Clin. Nutr. 2003, 77, 326–330. [Google Scholar]
- Jauhiainen, T.; Vapaatalo, H.; Poussa, T.; Kyrönpalo, S.; Rasmussen, M.; Korpela, R. Lactobacillus helveticus fermented milk lowers blood pressure in hypertensive subjects in 24-h ambulatory blood pressure measurement. Am. J. Hypertens. 2005, 18, 1600–1605. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Yoshikawa, M. LKPNM: A prodrug-type ACE-inhibitory peptide derived from fish protein. Immunopharmacology 1999, 44, 123–127. [Google Scholar]
- Matsufuji, H.; Matsui, T.; Seki, E.; Osajima, K.; Nakashima, M.; Osajima, Y. Angiotensin I-converting enzyme inhibitory peptides in an alkaline protease hydrolyzate derived from sardine muscle. Biosci. Biotechnol. Biochem. 1994, 58, 2244–2245. [Google Scholar]
- Ono, S.; Hosokawa, M.; Miyashita, K.; Takahashi, K. Isolation of peptides with angiotensin I-converting enzyme inhibitory effect derived from hydrolysate of upstream chum salmon muscle. J. Food Sci. 2003, 68, 1611–1614. [Google Scholar]
- Yokoyama, K.; Chiba, H.; Yoshikawa, M. Peptide inhibitors for angiotensin I-converting enzyme from thermolysin digest of dried bonito. Biosci. Biotechnol. Biochem. 1992, 56, 1541–1545. [Google Scholar]
- Hayes, M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Putting microbes to work: Dairy fermentation, cell factories and bioactive peptides. Part II: Bioactive peptide functions. J. Biotechnol. 2007, 2, 435–449. [Google Scholar]
- Cushman, D.W.; Cheung, H.S. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar]
- Rho, S.J.; Lee, J.-S.; Chung, Y.I.; Kim, Y.-W.; Lee, H.G. Purification and identification of an angiotensin I-converting enzyme inhibitory peptide from fermented soybean extract. Process Biochem. 2009, 44, 490–493. [Google Scholar]
- Saiga, A.; Okumura, T.; Makihara, T.; Katsuta, S.; Shimizu, T.; Yamada, R.; Nishimura, T. Angiotensin I-converting enzyme inhibitory peptides in a hydrolyzed chicken breast muscle extract. J. Agric. Food Chem. 2003, 51, 1741–1745. [Google Scholar]
- Maruyama, S.; Mitachi, H.; Awaya, J.; Kurono, M.; Tomizuka, N.; Suzuki, H. Angiotensin I-converting enzyme inhibitory activity of the C-terminal hexapeptide of s1-casein. Agric. Biol. Chem. 1987, 51, 2557–2561. [Google Scholar] [CrossRef]
- Lawrie, R.A.; Ledward, D.A. Lawrie’s Meat Science. In Lawrie’s Meat Science, 7th; Lawrie, R.A., Ed.; Woodhead Publishing: Cambridge, UK, 2006; pp. 41–73. [Google Scholar]
- Spudich, J.A.; Watt, S. The regulation of rabbit skeletal muscle contraction. J. Biol. Chem. 1971, 246, 4866–4871. [Google Scholar]
- Muguruma, M.; Ahhmed, A.M.; Katayama, K.; Kawahara, S.; Maruyama, M.; Nakamura, T. Identification of pro-drug type ACE inhibitory peptide sourced from porcine myosin B: Evaluation of its antihypertensive effects in vivo. Food Chem. 2009, 114, 516–522. [Google Scholar] [CrossRef]
- Katayama, K.; Mori, T.; Kawahara, S.; Miake, K.; Kodama, Y.; Sugiyama, M.; Kawamura, Y.; Nakayama, T.; Maruyama, M.; Muguruma, M. Angiotensin-I converting enzyme inhibitory peptide derived from porcine skeletal muscle myosin and its antihypertensive activity in spontaneously hypertensive rats. J. Food Sci. 2007, 72, S702–S706. [Google Scholar]
- Katayama, K.; Tomatsu, M.; Fuchu, H.; Sugiyama, M.; Kawahara, S.; Yamauchi, K.; Kawamura, Y.; Muguruma, M. Purification and characterization of an angiotensin I converting enzyme inhibitory peptide derived from porcine troponin C. Anim. Sci. J. 2003, 74, 53–58. [Google Scholar]
- Escudero, E.; Sentandreu, M.A.; Arihara, K.; Toldrá, F. Angiotensin I-converting enzyme inhibitory peptides generated from in vitro gastrointestinal digestion of pork meat. J. Agric. Food Chem. 2010, 58, 2895–2901. [Google Scholar] [CrossRef] [PubMed]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens—structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef] [PubMed]
- Jang, A.; Cho, Y.J.; Lee, J.I.; Shin, J.H.; Kim, I.S.; Lee, M. The effect of beef peptide on blood pressure and serum lipid concentration of spontaneously hypertensive rat (SHR). J. Anim. Sci. Technol. 2004, 46, 107–114. [Google Scholar]
- Iroyukifujita, H.; Eiichiyokoyama, K.; Yoshikawa, M. Classification and antihypertensive activity of angiotensin I converting enzyme inhibitory peptides derived from food proteins. J. Food Sci. 2000, 65, 564–569. [Google Scholar]
- Suetsuna, K.; Osajima, K. The inhibitory activities against angiotensin I-converting enzyme of basic peptides originating from sardine and hair tail meat. Bull. Jpn. Soc. Sci. Fish. 1986, 52, 1981–1984. [Google Scholar]
- Hai-Lun, H.; Xiu-Lan, C.; Cai-Yun, S.; Yu-Zhong, M.; Bai-Cheng, Z. Analysis of novel angiotensin-I-converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis. J. Pept. Sci. 2006, 12, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Theodore, A.E.; Kristinsson, H.G. Angiotensin converting enzyme inhibition of fish protein hydrolysates prepared from alkaline-aided channel catfish protein isolate. J. Sci. Food Agric. 2007, 87, 2353–2357. [Google Scholar]
- Bougatef, A.; Nedjar-Arroume, N.; Ravallec-Plé, R.; Leroy, Y.; Guillochon, D.; Barkia, A.; Nasri, M. Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained by treatment with microbial and visceral fish serine proteases. Food Chem. 2008, 111, 350–356. [Google Scholar] [CrossRef]
- Samaranayaka, A.G.P.; Kitts, D.D.; Li-Chan, E.C.Y. Antioxidative and angiotensin-I-converting enzyme inhibitory potential of a Pacific hake (Merluccius productus) fish protein hydrolysate subjected to simulated gastrointestinal digestion and Caco-2 cell permeation. J. Agric. Food Chem. 2010, 58, 1535–1542. [Google Scholar] [PubMed]
- Je, J.-Y.; Park, P.-J.; Kwon, J.Y.; Kim, S.-K. A Novel Angiotensin I Converting Enzyme Inhibitory Peptide from Alaska Pollack (Theragra chalcogramma) Frame Protein Hydrolysate. J. Agric. Food Chem. 2004, 52, 7842–7845. [Google Scholar] [PubMed]
- Jung, W.-K.; Mendis, E.; Je, J.-Y.; Park, P.-J.; Son, B.W.; Kim, H.C.; Choi, Y.K.; Kim, S.-K. Angiotensin I-converting enzyme inhibitory peptide from yellowfin sole (Limanda aspera) frame protein and its antihypertensive effect in spontaneously hypertensive rats. Food Chem. 2006, 94, 26–32. [Google Scholar]
- Ono, S.; Hosokawa, M.; Miyashita, K.; Takahashi, K. Inhibition properties of dipeptides from salmon muscle hydrolysate on angiotensin I-converting enzyme. Int. J. Food Sci. Technol. 2006, 41, 383–386. [Google Scholar]
- Wu, H.; He, H.L.; Chen, X.L.; Sun, C.Y.; Zhang, Y.Z.; Zhou, B.C. Purification and identification of novel angiotensin-I-converting enzyme inhibitory peptides from shark meat hydrolysate. Process Biochem. 2008, 43, 457–461. [Google Scholar]
- Kawasaki, T.; Seki, E.; Osajima, K.; Yoshida, M.; Asada, K.; Matsui, T.; Osajima, Y. Antihypertensive effect of valyl-tyrosine, a short chain peptide derived from sardine muscle hydrolyzate, on mild hypertensive subjects. J. Hum. Hypertens. 2000, 14, 519–523. [Google Scholar] [PubMed]
- Matsufuji, H.; Matsui, T.; Ohshige, S.; Kawasaki, T.; Osajima, K.; Osajima, Y. Antihypertensive effects of angiotensin fragments in SHR. Biosci. Biotechnol. Biochem. 1995, 59, 1398–1401. [Google Scholar]
- Kohama, Y.; Matsumoto, S.; Oka, H.; Teramoto, T.; Okabe, M.; Mimura, T. Isolation of angiotensin-converting enzyme inhibitor from tuna muscle. Biochem. Biophys. Res. Commun. 1988, 155, 332–337. [Google Scholar]
- Byun, H.-G.; Kim, S.-K. Purification and characterization of angiotensin I converting enzyme (ACE) inhibitory peptides from Alaska pollack (Theragra chalcogramma) skin. Process Biochem. 2001, 36, 1155–1162. [Google Scholar] [CrossRef]
- Ames, B.N. Dietary carcinogens and anticarcinogens. Oxygen radicals and degenerative diseases. Science 1983, 221, 1256–1264. [Google Scholar] [PubMed]
- Gimenez, B.; Aleman, A.; Montero, P.; Gomez-Guillen, M.C. Antioxidant and functional properties of gelatin hydrolysates obtained from skin of sole and squid. Food Chem. 2009, 114, 976–983. [Google Scholar]
- Cai, W.; Gao, Q.D.; Zhu, L.; Peppa, M.; He, C.; Vlassara, H. Oxidative stress-inducing carbonyl compounds from common foods: Novel mediators of cellular dysfunction. Mol. Med. 2002, 8, 337–346. [Google Scholar]
- Esterbauer, H. Cytotoxicity and genotoxicity of lipid-oxidation products. Am. J. Clin. Nutr. 1993, 57, 779S–785S. [Google Scholar]
- Pihlanto, A. Antioxidative peptides derived from milk proteins. Int. Dairy J. 2006, 16, 1306–1314. [Google Scholar]
- Vercruysse, L.; Smagghe, G.; Beckers, T.; Camp, J.V. Antioxidative and ACE inhibitory activities in enzymatic hydrolysates of the cotton leafworm, Spodoptera littoralis. Food Chem. 2009, 114, 38–43. [Google Scholar] [CrossRef]
- Valenzuela, A.; Sanhueza, J.; Nieto, S. Natural antioxidants in functional foods: From food safety to health benefits. Grasas Aceites 2003, 54, 295–303. [Google Scholar]
- Moure, A.; Cruz, J.M.; Franco, D.; Domínguez, J.M.; Sineiro, J.; Domínguez, H.; José Núñez, M.; Parajó, J.C. Natural antioxidants from residual sources. Food Chem. 2001, 72, 145–171. [Google Scholar]
- Suetsuna, K.; Ukeda, H.; Ochi, H. Isolation and characterization of free radical scavenging activities peptides derived from casein. J. Nutr. Biochem. 2000, 11, 128–131. [Google Scholar]
- Sun, J.; He, H.; Xie, B.J. Novel antioxidant peptides from fermented mushroom Ganoderma lucidum. J. Agric. Food Chem. 2004, 52, 6646–6652. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Chen, H.M.; Shiau, C.Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Je, J.Y.; Park, P.J.; Kim, S.K. Antioxidant activity of a peptide isolated from Alaska pollack (Theragra chalcogramma) frame protein hydrolysate. Food Res. Int. 2005, 38, 45–50. [Google Scholar] [CrossRef]
- Raghavan, S.; Kristinsson, H.G.; Leeuwenburgh, C. Radical scavenging and reducing ability of tilapia (Oreochromis niloticus) protein hydrolysates. J. Agric. Food Chem. 2008, 56, 10359–10367. [Google Scholar] [PubMed]
- Dong, S.Y.; Zeng, M.Y.; Wang, D.F.; Liu, Z.Y.; Zhao, Y.H.; Yang, H.C. Antioxidant and biochemical properties of protein hydrolysates prepared from Silver carp (Hypophthalmichthys molitrix). Food Chem. 2008, 107, 1485–1493. [Google Scholar] [CrossRef]
- Klompong, V.; Benjakul, S.; Yachai, M.; Visessanguan, W.; Shahidi, F.; Hayes, K. Amino acid composition and antioxidative peptides from protein hydrolysates of yellow stripe trevally (Selaroides leptolepis). J. Food Sci. 2009, 74, C126–C133. [Google Scholar] [PubMed]
- Bougatef, A.; Nedjar-Arroume, N.; Manni, L.; Ravallec, R.; Barkia, A.; Guillochon, D.; Nasri, M. Purification and identification of novel antioxidant peptides from enzymatic hydrolysates of sardinelle (Sardinella aurita) by-products proteins. Food Chem. 2010, 118, 559–565. [Google Scholar] [CrossRef]
- Hsu, K.-C. Purification of antioxidative peptides prepared from enzymatic hydrolysates of tuna dark muscle by-product. Food Chem. 2010, 122, 42–48. [Google Scholar]
- Mendis, E.; Rajapakse, N.; Kim, S.-K. Antioxidant Properties of a Radical-Scavenging Peptide Purified from Enzymatically Prepared Fish Skin Gelatin Hydrolysate. J. Agric. Food Chem. 2004, 53, 581–587. [Google Scholar]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar]
- Li, B.; Chen, F.; Wang, X.; Ji, B.P.; Wu, Y.N. Isolation and identification of antioxidative peptides from porcine collagen hydrolysate by consecutive chromatography and electrospray ionization-mass spectrometry. Food Chem. 2007, 102, 1135–1143. [Google Scholar]
- Liu, Q.; Kong, B.; Xiong, Y.L.; Xia, X. Antioxidant activity and functional properties of porcine plasma protein hydrolysate as influenced by the degree of hydrolysis. Food Chem. 2010, 118, 403–410. [Google Scholar]
- Wu, H.C.; Pan, B.S.; Chang, C.L.; Shiau, C.Y. Low-molecular-weight peptides as related to antioxidant properties of chicken essence. J. Food Drug Anal. 2005, 13, 176–183. [Google Scholar]
- Hayes, M.; Ross, R.P.; Fitzgerald, G.F.; Hill, C.; Stanton, C. Casein-derived antimicrobial peptides generated by Lactobacillus acidophilus DPC6026. Appl. Environ. Microbiol. 2006, 72, 2260–2264. [Google Scholar] [PubMed]
- McCann, K.B.; Shiell, B.J.; Michalski, W.P.; Lee, A.; Wan, J.; Roginski, H.; Coventry, M.J. Isolation and characterisation of antibacterial peptides derived from the f (164-207) region of bovine [alpha] S2-casein. Int. Dairy J. 2005, 15, 133–143. [Google Scholar]
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identification of the bactericidal domain of lactoferrin. Biochim. Biophys. Acta 1992, 1121, 130–136. [Google Scholar] [CrossRef] [PubMed]
- van der Kraan, M.I.A.; Groenink, J.; Nazmi, K.; Veerman, E.C.I.; Bolscher, J.G.M.; Nieuw Amerongen, A.V. Lactoferrampin: A novel antimicrobial peptide in the N1-domain of bovine lactoferrin. Peptides 2004, 25, 177–183. [Google Scholar]
- Lee, Y.; Kim, J.I.Y.; Lee, K.; Kim, K.H.; Lee, H. Peptides from anchovy sauce induce apoptosis in a human lymphoma cell (U937) through the increase of caspase 3 and 8 activities. Ann. N. Y. Acad. Sci. 2003, 1010, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.G.; Lee, K.W.; Kim, J.Y.; Kim, K.H.; Lee, H.J. Induction of apoptosis in a human lymphoma cell line by hydrophobic peptide fraction separated from anchovy sauce. BioFactors 2004, 21, 63–67. [Google Scholar]
- Picot, L.; Bordenave, S.; Didelot, S.; Fruitier-Arnaudin, I.; Sannier, F.; Thorkelsson, G.; Berge, J.P.; Guerard, F.; Chabeaud, A.; Piot, J.M. Antiproliferative activity of fish protein hydrolysates on human breast cancer cell lines. Process Biochem. 2006, 41, 1217–1222. [Google Scholar]
- Hsu, K.-C.; Li-Chan, E.C.Y.; Jao, C.-L. Antiproliferative activity of peptides prepared from enzymatic hydrolysates of tuna dark muscle on human breast cancer cell line MCF-7. Food Chem. 2010, 126, 617–622. [Google Scholar]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar]
- Slim, M.-K. Cardiovascular actions of chicken-meat extract in normo- and hypertensive rats. Br. J. Nutr. 2001, 86, 97–103. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ryan, J.T.; Ross, R.P.; Bolton, D.; Fitzgerald, G.F.; Stanton, C. Bioactive Peptides from Muscle Sources: Meat and Fish. Nutrients 2011, 3, 765-791. https://doi.org/10.3390/nu3090765
Ryan JT, Ross RP, Bolton D, Fitzgerald GF, Stanton C. Bioactive Peptides from Muscle Sources: Meat and Fish. Nutrients. 2011; 3(9):765-791. https://doi.org/10.3390/nu3090765
Chicago/Turabian StyleRyan, Joseph Thomas, Reynolds Paul Ross, Declan Bolton, Gerald F. Fitzgerald, and Catherine Stanton. 2011. "Bioactive Peptides from Muscle Sources: Meat and Fish" Nutrients 3, no. 9: 765-791. https://doi.org/10.3390/nu3090765
APA StyleRyan, J. T., Ross, R. P., Bolton, D., Fitzgerald, G. F., & Stanton, C. (2011). Bioactive Peptides from Muscle Sources: Meat and Fish. Nutrients, 3(9), 765-791. https://doi.org/10.3390/nu3090765