An Extract from Wax Apple (Syzygium samarangense (Blume) Merrill and Perry) Effects Glycogenesis and Glycolysis Pathways in Tumor Necrosis Factor-α-Treated FL83B Mouse Hepatocytes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Abbreviations
| TNF-α | tumor necrosis factor-α |
| WAE | wax apple aqueous extract |
| 2-NBDG | 2-[N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino]-2-deoxyglucose |
| KRB buffer | Krebs-Ringer Bicarbonate buffer |
| GS | glycogen synthase |
| HXK | hexokinase |
| G6PD | glucose-6-phosphate dehydrogenase |
| PFK | phosphofructokinase |
| DM | diabetes mellitus |
| F12K | F12 Ham Kaighn’s |
| FBS | Fetal bovine serum |
| PBS | phosphate buffered saline |
| SDS | sodium dodecyl sulfate |
| EDTA | ethylenediamine tetraacetic acid |
| PMSF | phenylmethanesulfonyl fluoride |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| PBST | phosphate buffer saline and Tween 20 |
| HRP | horseradish peroxidase |
| ECL | enhanced chemiluminescence |
| G6P | glucose-6-phosphate |
| PP pathway | pentose phosphate pathway |
| NADPH | nicotinamide adenine dinucleotide phosphate reduced |
| ROS | reactive oxygen species |
| PI3K | phosphatidylinositol-3 kinase |
| DHAP | dihydroxyacetone phosphate. |
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and the Extraction, Isolation and Purification of WAE
2.3. Cell Culture

2.4. Induction of Insulin Resistance Using TNF-α and Cell Preparation
2.5. Determination of Glycogen
2.6. Protein Extraction from Cells
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
3.1. Effect of WAE on Glycogen Content in Insulin Resistant FL83B Mouse Hepatocytes

3.2. Glycogen Synthase Expression

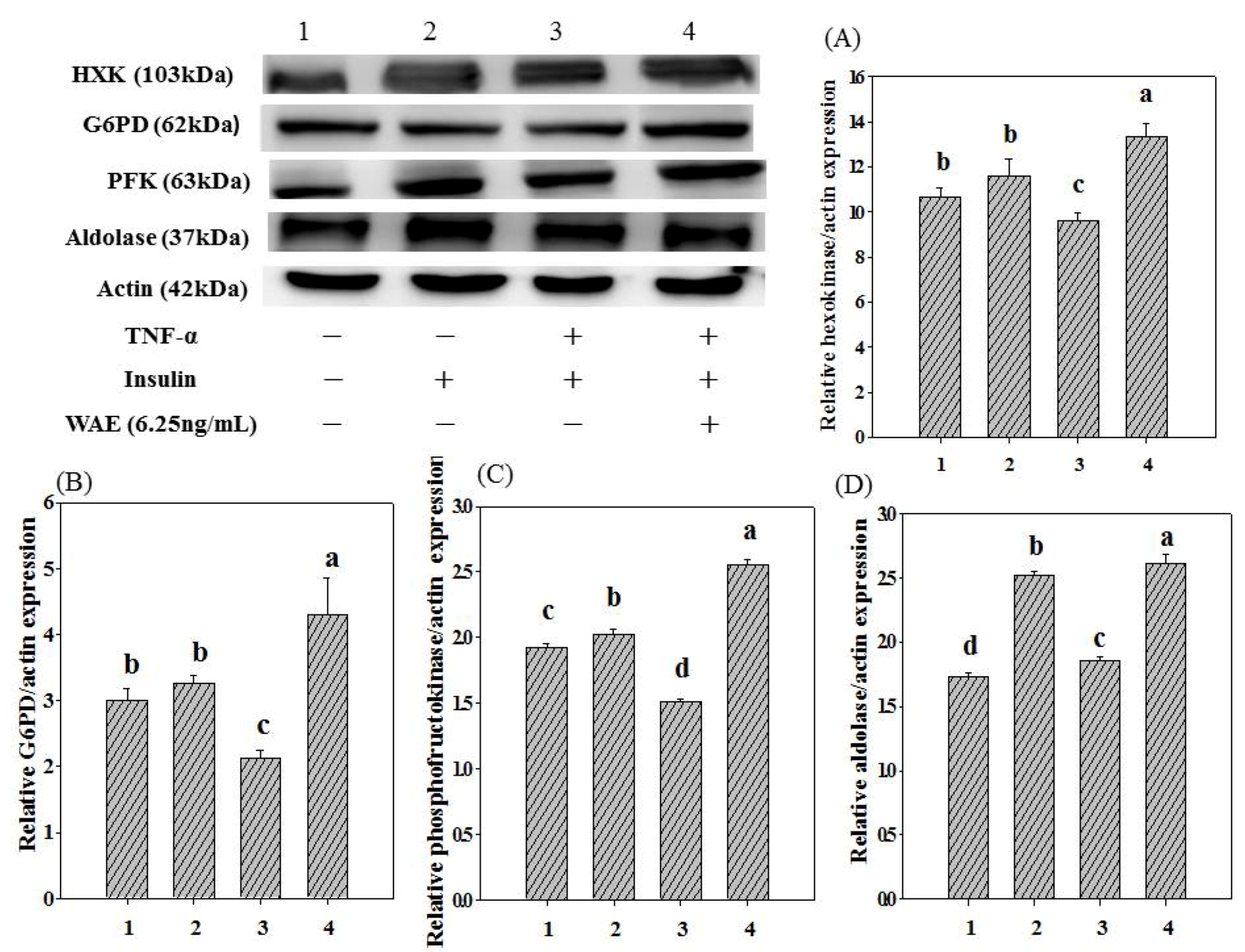
3.3. Glycolysis-Related Enzyme Expression

4. Discussion
5. Conclusions
Acknowledgements
Conflict of Interest
References
- Roa, B.K.; Sudarshan, P.R.; Rajasekhar, M.D.; Nagaraju, N.; Roa, C.A. Antidiabetic activity of Terminalia pallida fruit in alloxan induced diabetic rats. J. Ethnopharmacol. 2003, 85, 169–172. [Google Scholar] [CrossRef]
- Cheng, H.L.; Huang, H.K.; Chang, C.I.; Tsai, C.P.; Chou, C.H. A cell-based screening identifies compounds from the stem of Momordica charantia that overcome insulin resistance and activate AMP activated protein kinase. J. Agric. Food Chem. 2008, 56, 6835–6843. [Google Scholar] [CrossRef]
- Hengstler, J.G.; Utesch, D.; Steinberg, P.; Platt, K.L.; Diener, B.; Ringel, M.; Swales, N.; Fischer, T.; Biefang, K.; Gerl, M.; Böttger, T.; Oesch, F. Cryopreserved primary hepatocytes as a constantly available in vitro model for the evaluation of human and animal drug metabolism and enzyme induction. Drug Metab. Rev. 2000, 32, 81–118. [Google Scholar] [CrossRef]
- Rates, S.M. Plants as source of drugs. Toxicon 2001, 39, 603–613. [Google Scholar]
- Gurib-Fakim, A. Phytochemical screening of 38 Mauritian medicinal plants. Rev. Agric. Sucr. Ile Maurice 1996, 69, 42–50. [Google Scholar]
- Neergheen, V.; Soobrattee, M.; Bahorun, T.; Aruoma, O. Characterization of the phenolic constituents in Mauritian endemic plants as determinants of their antioxidant activities in vitro. J. Plant Physiol. 2006, 163, 787–799. [Google Scholar] [CrossRef]
- Resurreccion-Magno, M.; Villasenor, I.; Harada, N.; Monde, K. Antihyperglycaemic flavonoids from Syzygium samarangense (Blume) Merr. and Perry. Phytother. Res. 2005, 19, 246–251. [Google Scholar] [CrossRef]
- Shen, S.C.; Chang, W.C.; Chang, C.L. Fraction from wax apple [Syzygium samarangense (Blume) Merrill and Perry] fruit extract ameliorates insulin resistance via modulating insulin signaling and inflammation pathway in tumor necrosis factor α-treated FL83B mouse hepatocytes. Int. J. Mol. Sci. 2012, 13, 8562–8577. [Google Scholar] [CrossRef]
- Huang, D.W.; Shen, S.C.; Wu, J.S.B. Effects of caffeic acid and cinnamic acid on glucose uptake in insulin-resistant mouse hepatocytes. J. Agric. Food Chem. 2009, 57, 7687–7692. [Google Scholar] [CrossRef]
- Kim, H.P.; Son, K.H.; Chang, H.W.; Kang, S.S. Anti-inflammatory plant flavonoids and cellular action mechanism. J. Pharmacol. Sci. 2004, 96, 229–245. [Google Scholar] [CrossRef]
- Ferrer, J.C.; Favre, C.; Gomis, R.R.; Fernandez-Novell, J.M.; Garica-Rocha, M.; de la Iglesia, N.; Cid, E.; Guinovart, J.J. Control of glycogen deposition. FEBS Lett. 2003, 546, 127–132. [Google Scholar] [CrossRef]
- Iynedjian, P.B. Molecular physiology of mammalian glucokinase. Cell. Mol. Life Sci. 2009, 66, 27–42. [Google Scholar] [CrossRef]
- Panneerselvam, R.S.; Govindaswamy, S. Effect of sodium molybdate on carbohydrate metabolizing enzymes in alloxan-induced diabetic rat. J. Nutr. Biochem. 2002, 13, 21–26. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Kahn, C.R. Insulin signaling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef]
- Zick, Y. Insulin resistance: A phosphorylation-based uncoupling of insulin signaling. Trends Cell Biol. 2001, 11, 437–441. [Google Scholar]
- Cheng, J.T.; Liu, I.M. Stimulatory effect of caffeic acid on α1A-adrenoceptors to increase glucose uptake into cultured C2C12 cells. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2000, 362, 122–127. [Google Scholar] [CrossRef]
- Cichy, S.B.; Uddin, S.; Danilkovich, A.; Guo, S.; Klippel, A.; Unterman, T.G. Protein kinase B/Akt mediates effect of insulin on hepatic insulin-like growth factor-binding protein-1 gene expression through a conserved insulin response sequence. J. Biol. Chem. 1998, 273, 6482–6487. [Google Scholar]
- González-Espinosa, C.; Romero-Ávila, M.T.; Mora-Rodríguez, D.M.; González-Espinosa, D.; García-Sáinz, J.A. Molecular cloning and functional expression of the guinea pig α1A-adrenoceptor. Eur. J. Pharmacol. 2001, 426, 147–155. [Google Scholar] [CrossRef]
- Gropper, S.S.; Smith, J.L.; Groff, J.L. Advanced Nutrition and Human Metabolism, 5th ed; Wadsworth Publishing: Belmont, CA, USA, 2009; p. 72. [Google Scholar]
- Clore, J.N.; Stillman, J.; Sugerman, H. Glucose-6-phosphate flux in vitro is increased in type 2 diabetes. Diabetes 2000, 49, 969–974. [Google Scholar] [CrossRef]
- Ivy, J.L.; Sherman, W.M.; Cuyler, C.L.; Katz, A.L. Exercise and diet reduce muscle insulin resistance in obese Zucker rat. Am. J. Physiol. 1986, 251, E299–E305. [Google Scholar]
- Abdel-Rahim, E.A.; El-Saadany, S.S.; Abo-Eytta, A.M.; Wasif, M.M. The effect of sammo administration on some fundamental enzymes of pentose phosphate pathway and energy metabolities of alloxanized rats. Nahrung 1992, 36, 8–14. [Google Scholar] [CrossRef]
- Nikolaidis, M.G.; Jamurtas, A.Z.; Paschalis, V.; Kostaropoulos, I.A.; Kladi-Skandali, A.; Balamitsi, V.; Koutedakis, Y.; Kouretas, D. Exercise-induced oxidative stress in G6PD-deficient individuals. Med. Sci. Sports Exerc. 2006, 38, 1443–1450. [Google Scholar] [CrossRef]
- Ho, H.Y.; Cheng, M.L.; Chiu, D.T. Glucose-6-phosphate dehydrogenase-from oxidative stress to cellular functions and degenerative diseases. Redox. Rep. 2007, 12, 109–118. [Google Scholar] [CrossRef]
- Wagle, A.; Jivraj, S.; Garlock, G.L.; Stapleton, S.R. Insulin regulation of glucose-6-phosphate dehydrogenase gene expression is rapamycin-sensitive and requires phosphatidylinositol 3-kinase. J. Biol. Chem. 1998, 273, 14968–14974. [Google Scholar]
- Strack, T. Genetics and molecular biology protein kinase C-[zeta] as an AMP-activated protein kinase kinase kinase: The protein kinase C-[zeta]-LKB1-AMP-activated protein kinase pathway. Drugs Today (Barc.) 2008, 44, 303–314. [Google Scholar] [CrossRef]
- Silva, D.D.; Zancan, P.; Coelho, W.S.; Gomez, L.S.; Sola-Penna, M. Metformin reverses hexokinase and 6-phosphofructo-1-kinase inhibition in skeletal muscle, liver and adipose tissue from streptozotocin-induced diabetic mouse. Arch. Biochem. Biophys. 2010, 496, 53–60. [Google Scholar] [CrossRef]
- Deprez, J.; Vertommen, D.; Alessi, D.R.; Hue, L.; Rider, M.H. Phosphorylation and activation of heart 6-phosphofructo-2-kinase by protein kinase B and other protein kinases of the insulin signaling cascades. J. Biol. Chem. 1997, 272, 17269–17275. [Google Scholar]
- Spitz, G.A.; Furtado, C.M.; Sola-Penna, M.; Zancan, P. Acetylsalicylic acid and salicylic acid decrease tumor cell viability and glucose metabolism modulating 6-phosphofructo-1-kinase structure and activity. Biochem. Pharmacol. 2009, 77, 46–53. [Google Scholar]
- Yamakoshi, Y.; Nagano, T.; Hu, J.C.; Yamakoshi, F.; Simmer, J.P. Porcine dentin sialoprotein glycosylation and glycosaminoglycan attachments. BMC Biochem. 2011, 3, 1–13. [Google Scholar]
- Huang, Y.; Shinzawa, H.; Togashi, H.; Takahashi, T.; Kuzumaki, T.; Otsu, K.; Ishikawa, K. Interleukin-6 down regulates expression of the aldolase B and albumin genes through a pathway involving the activation of tyrosine kinase. Arch. Biochem. Biophys. 1995, 320, 203–209. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shen, S.-C.; Chang, W.-C.; Chang, C.-L. An Extract from Wax Apple (Syzygium samarangense (Blume) Merrill and Perry) Effects Glycogenesis and Glycolysis Pathways in Tumor Necrosis Factor-α-Treated FL83B Mouse Hepatocytes. Nutrients 2013, 5, 455-467. https://doi.org/10.3390/nu5020455
Shen S-C, Chang W-C, Chang C-L. An Extract from Wax Apple (Syzygium samarangense (Blume) Merrill and Perry) Effects Glycogenesis and Glycolysis Pathways in Tumor Necrosis Factor-α-Treated FL83B Mouse Hepatocytes. Nutrients. 2013; 5(2):455-467. https://doi.org/10.3390/nu5020455
Chicago/Turabian StyleShen, Szu-Chuan, Wen-Chang Chang, and Chiao-Li Chang. 2013. "An Extract from Wax Apple (Syzygium samarangense (Blume) Merrill and Perry) Effects Glycogenesis and Glycolysis Pathways in Tumor Necrosis Factor-α-Treated FL83B Mouse Hepatocytes" Nutrients 5, no. 2: 455-467. https://doi.org/10.3390/nu5020455
APA StyleShen, S. -C., Chang, W. -C., & Chang, C. -L. (2013). An Extract from Wax Apple (Syzygium samarangense (Blume) Merrill and Perry) Effects Glycogenesis and Glycolysis Pathways in Tumor Necrosis Factor-α-Treated FL83B Mouse Hepatocytes. Nutrients, 5(2), 455-467. https://doi.org/10.3390/nu5020455