Fermented Brown Rice and Rice Bran with Aspergillus oryzae (FBRA) Prevents Inflammation-Related Carcinogenesis in Mice, through Inhibition of Inflammatory Cell Infiltration


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
2.2. Mice
2.3. Inflammation-Related Carcinogenesis Model
2.4. Subcutaneous Tumor Growth
2.5. Determination of the Total Numbers and the Cell Types of the Peripheral Blood Leukocytes, Bone Marrow Cells and Gelatin Sponge-Infiltrated Cells
2.6. Immunohistochemistry
2.7. Quantification of Immunohistochemically Positive Cells
2.8. RNA Extraction, cDNA Preparation and Quantitative Real-Time PCR Analysis
2.9. Statistical Analysis
3. Results
3.1. Suppression of Inflammation-Related Carcinogenesis by Administration of FBRA

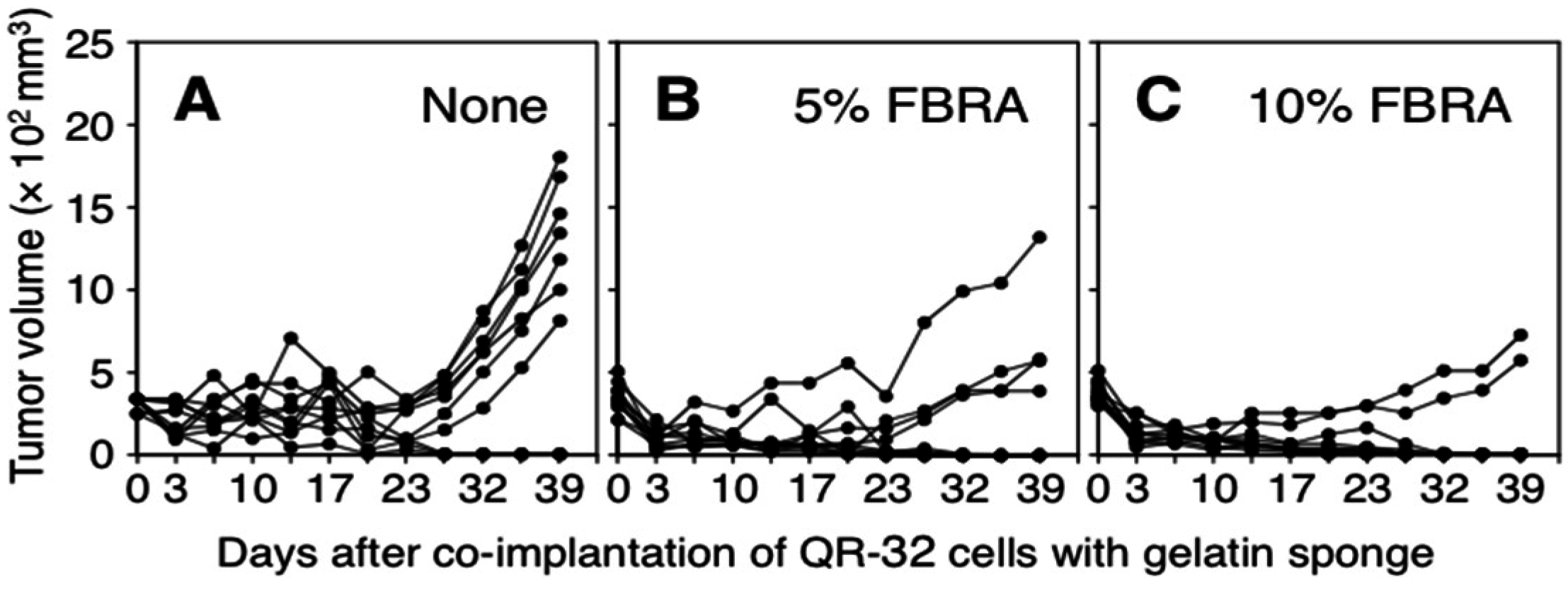{kind=link}
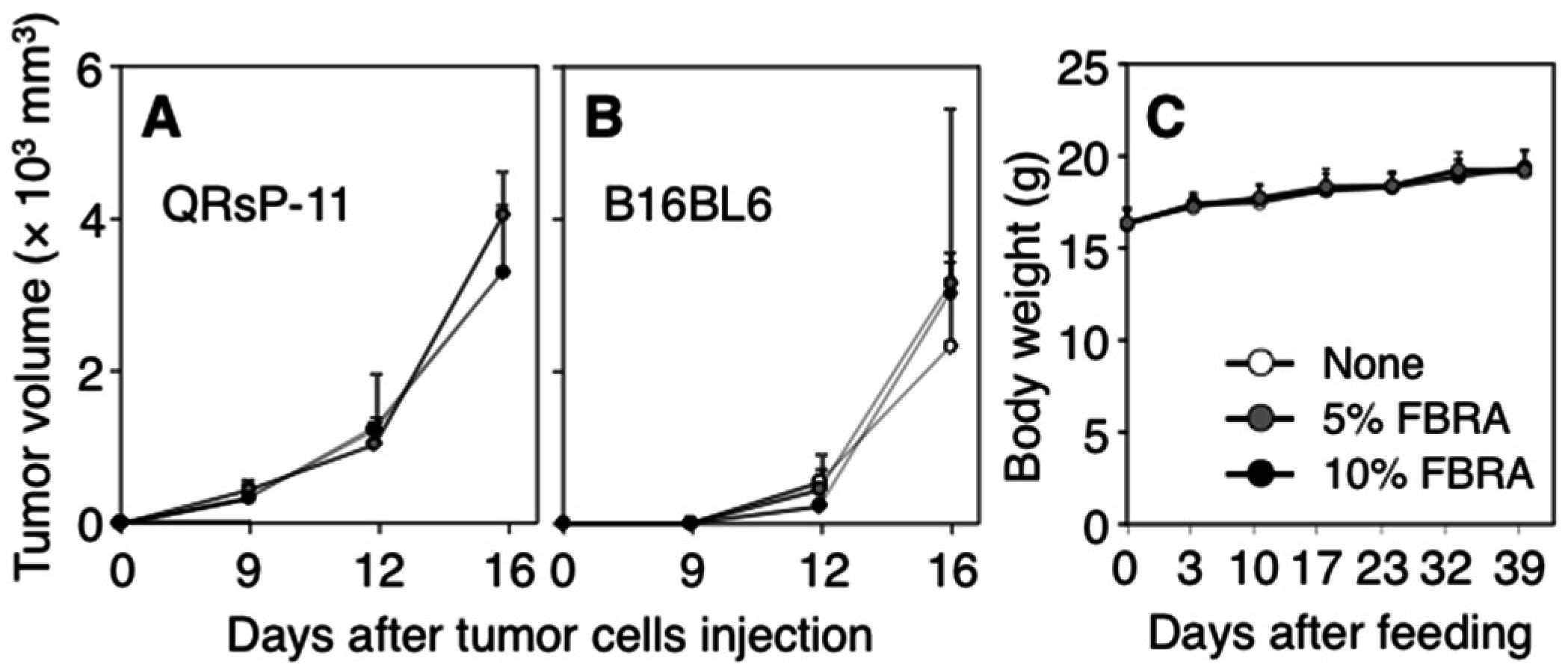{kind=link}
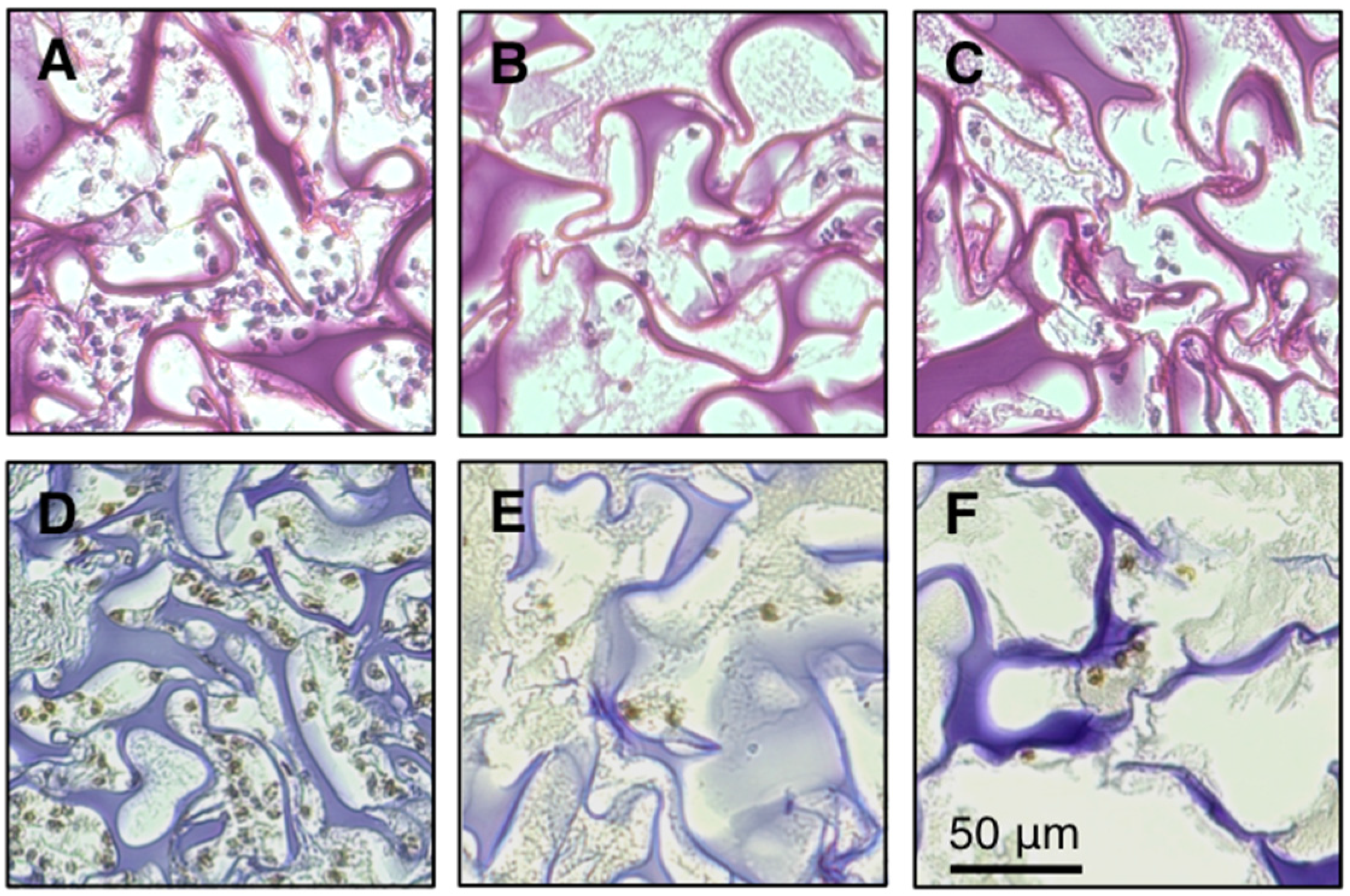{kind=link}
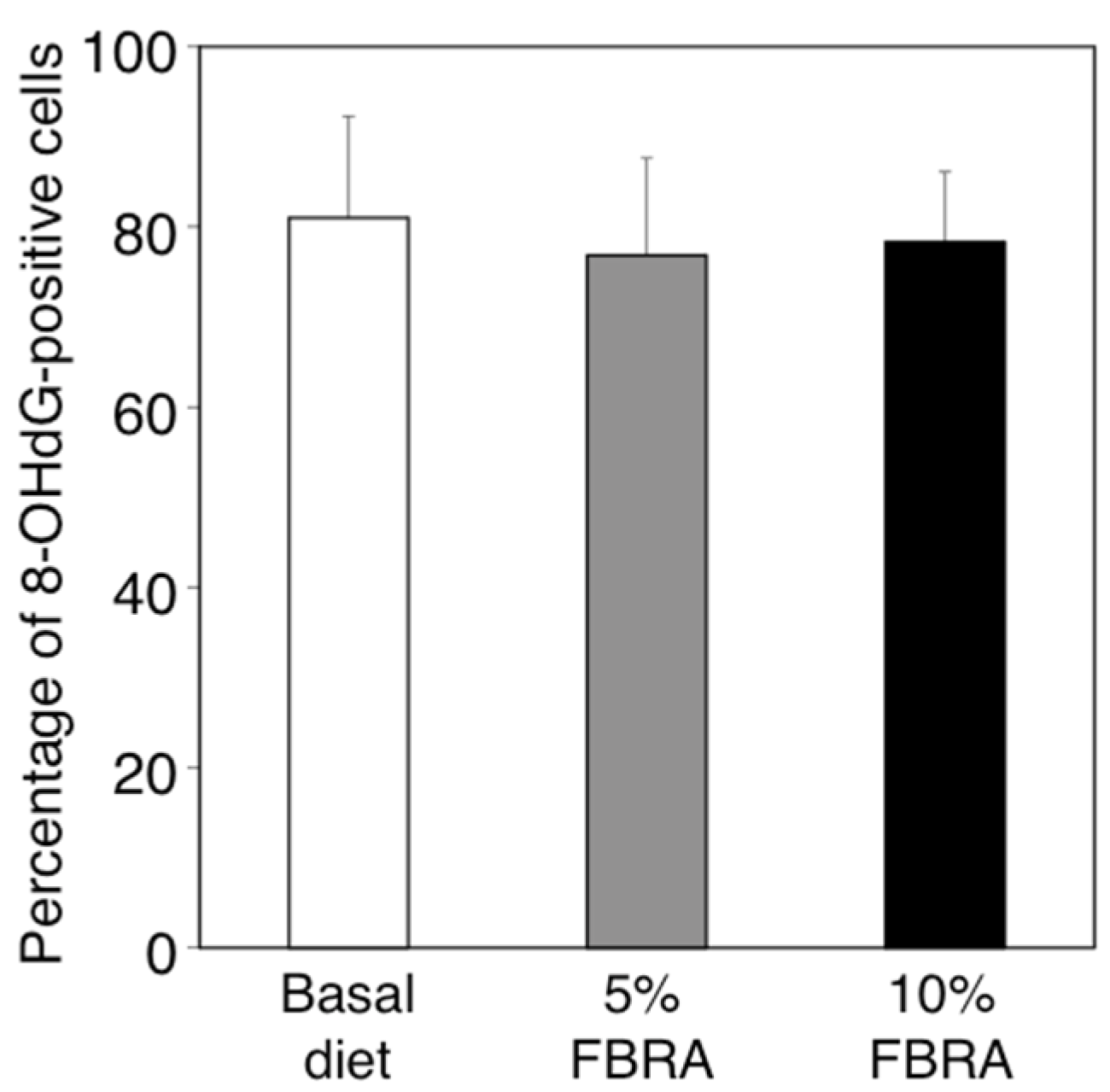{kind=link}
{kind=link}
| Treatment a | Gelatin Sponge Implantation | QR-32 Cells Injection b | No. of Mice with Tumor/No. of Mice Tested (%) | ||
|---|---|---|---|---|---|
| Experiment I | Experiment II | Total | |||
| None | - | + | 0/10 | 0/10 | 0/20 (0) |
| None | + | + | 7/10 | 7/10 | 14/20 (70) |
| 5% FBRA | + | + | 4/10 | 3/10 | 7/20 c (35) |
| 10% FBRA | + | + | 2/10c | 2/10c | 4/20 d (20) |

3.2. Effects of FBRA Administration on Proliferation of Transplantable Tumor Cells

| Cells | Subcutaneous Tumorigenicity a | Spontaneous Metastasis c | ||||
|---|---|---|---|---|---|---|
| Treatment b | No. of Mice with Tumor/No. of Mice Tested (%) | Mean Survival Time (Days) | No. of Mice with Lung Metastasis/No. of Mice Tested (%) | No. of Mice with Lymph Node (LN) Metastasis/No. of Mice Tested (%) | ||
| Inguinal LN | Axillary LN | |||||
| QRsP-11 | None | 5/5 (100) | 34 ± 6 | 0/5 (0) | 0/5 (0) | 0/5 (0) |
| 5% FBRA | 5/5 (100) | 33 ± 10 | 0/5 (0) | 0/5 (0) | 0/5 (0) | |
| 10% FBRA | 4/4 (100) | 25 ± 14 | 0/5 (0) | 0/5 (0) | 0/5 (0) | |
| B16BL6 | None | 5/5 (100) | 26 ± 4 | 2/5 (40) | 0/5 (0) | 0/5 (0) |
| 5% FBRA | 5/5 (100) | 27 ± 1 | 0/5 (0) | 1/5 (20) | 0/5 (0) | |
| 10% FBRA | 5/5 (100) | 28 ± 4 | 1/5 (20) | 1/5 (20) | 2/5 (40) | |
3.3. Inhibition of Infiltration of Inflammatory Cells into Gelatin Sponge by FBRA

| Treatment a | Co-Implantation of QR-32 Cells with Gelatin Sponge b | No. of Gelatin Sponge-Infiltrated Cells (× 104) c | No. of Peripheral Blood Leukocytes (× 102/μL) | No. of Bone Marrow Cells (× 106) |
|---|---|---|---|---|
| None | - | NA d | 18 ± 4 | 17 ± 2 |
| None | + | 143 ± 25 | 22 ± 6 | 20 ± 4 |
| 5% FBRA | + | 94 ± 24 e | 26 ± 4 | 16 ± 5 |
| 10% FBRA | + | 86 ± 22 e | 20 ± 5 | 18 ± 4 |
3.4. Timing of FBRA Administration for Suppressing the Infiltration of Inflammatory Cells
| Treatment Period | No. of Gelatin Sponge-Infiltrated Cells (× 104) a | ||
|---|---|---|---|
| None | 5% FBRA | 10% FBRA | |
| Day −2–5 | 215 ± 15 | 146 ± 21 b | 134 ± 21 b |
| Day 0–1 | 212 ± 35 | 200 ± 14 | 204 ± 19 |
| Day 0–3 | 261 ± 16 | 244 ± 31 | 245 ± 18 |
| Day 0–5 | 202 ± 30 | 215 ± 60 | 230 ± 30 |
| Day −1–0 | 233 ± 25 | 227 ± 26 | 218 ± 15 |
| Day −3–0 | 277 ± 15 | 145 ± 6 b | 140 ± 16 b |
| Day −5–0 | 238 ± 26 | 138 ± 32 b | 152 ± 27 b |
3.5. Types of Inflammatory Cells Affected by FBRA Administration
| Cell Source a | Treatment b | Co-Implantation of QR-32 Cells with Gelatin Sponge c | Differential Leukocyte Counts (%) d | ||
|---|---|---|---|---|---|
| Monocytes/Macrophages | Granulocytes | Lymphocytes | |||
| Gelatin sponge-Infiltrated | None | + | 11 ± 2 | 57 ± 3 | 32 ± 5 |
| 5% FBRA | + | 11 ± 3 | 59 ± 4 | 30 ± 1 | |
| 10% FBRA | + | 11 ± 1 | 59 ± 2 | 31 ± 3 | |
| Peripheral blood | None | - | 2 ± 1 | 18 ± 4 | 81 ± 4 |
| None | + | 2 ± 1 | 16 ± 4 | 82 ± 4 | |
| 5% FBRA | + | 3 ± 1 | 16 ± 6 | 81 ± 5 | |
| 10% FBRA | + | 3 ± 0 | 17 ± 7 | 81 ± 7 | |
| Bone marrow | None | - | 5 ± 2 | 72 ± 5 | 23 ± 3 |
| None | + | 5 ± 1 | 73 ± 2 | 22 ± 2 | |
| 5% FBRA | + | 5 ± 1 | 71 ± 3 | 24 ± 3 | |
| 10% FBRA | + | 5 ± 1 | 72 ± 3 | 23 ± 2 | |
3.6. Effects of FBRA Administration on the Ability to Induce Oxidative Stress

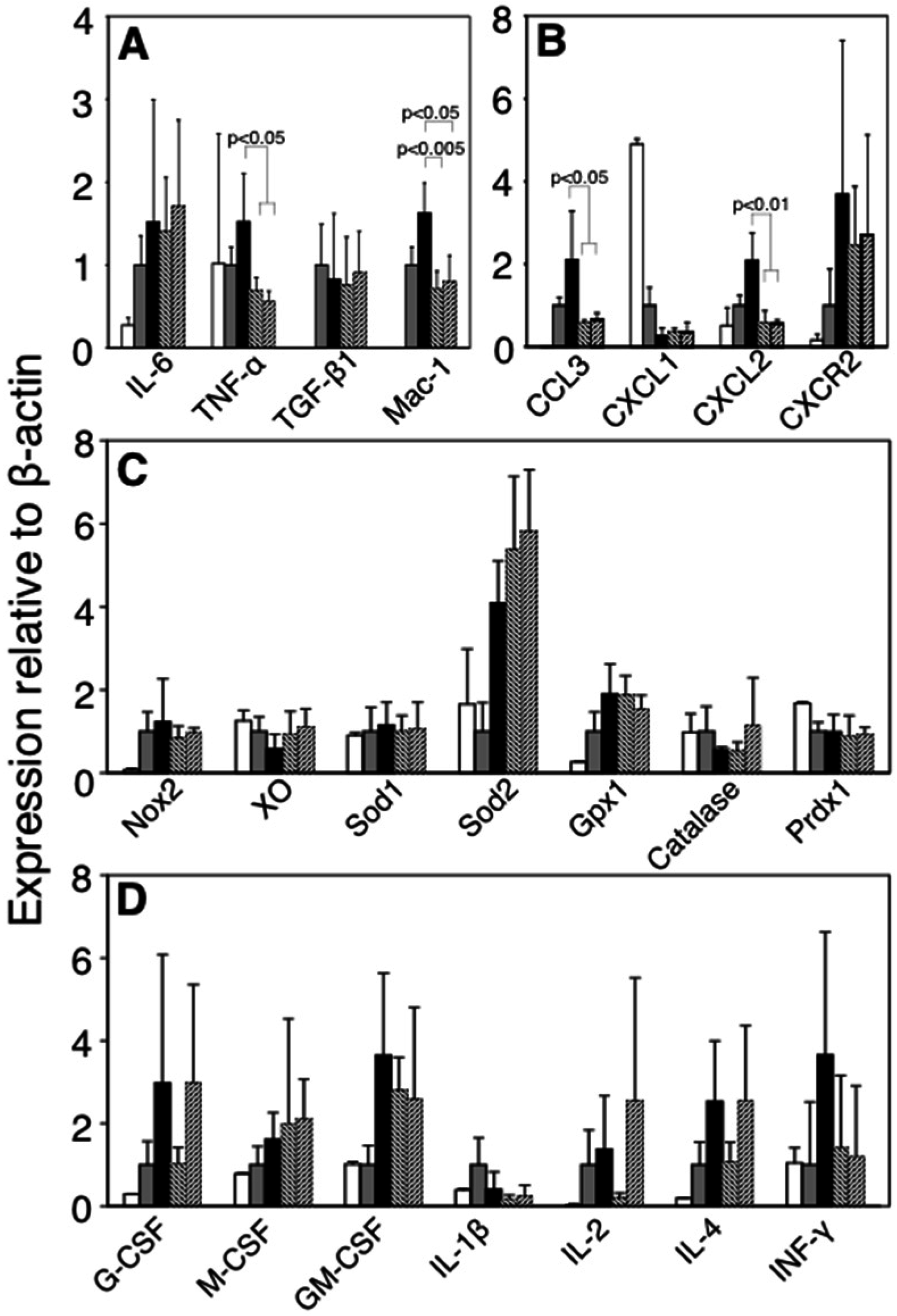
3.7. Changes in the Expressions of Inflammation-Related Genes at the Inflammatory Site by Administration of FBRA
3.8. No Change in Gene Expression of Bone Marrow Cells by FBRA

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Grivennikov, S.; Greten, F.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, A.; Jankowski, J. Acid reflux and oesophageal cancer. Recent Results Cancer Res. 2011, 185, 65–82. [Google Scholar] [PubMed]
- Zaridze, D.; Borisova, E.; Maximovitch, D.; Chkhikvadze, V. Aspirin protects against gastric cancer: Results of a case-control study from Moscow, Russia. Int. J. Cancer 1999, 82, 473–476. [Google Scholar] [CrossRef]
- Velayos, F.S.; Loftus, E.V., Jr.; Jess, T.; Harmsen, W.S.; Bida, J.; Zinsmeister, A.R.; Tremaine, W.J.; Sandborn, W.J. Predictive and protective factors associated with colorectal cancer in ulcerative colitis: A case-control study. Gastroenterol. 2006, 130, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- IARC Working Group on the Evaluation of Cancer Preventive Agents. Non-steroidal anti-inflammatory drugs. In IARC Handbook of Cancer Prevention; International Agency for Research on Cancer, Ed.; IARC Publications: Lyon, France, 1997; Volume 1, pp. 43–125. [Google Scholar]
- Suh, O.; Mettlin, C.; Petrelli, N.J. Aspirin use, cancer, and polyps of the large bowel. Cancer 1993, 72, 1171–1177. [Google Scholar] [CrossRef]
- Vonkeman, H.E.; van de Laar, M.A. Nonsteroidal anti-inflammatory drugs: Adverse effects and their prevention. Semin. Arthritis Rheum. 2010, 39, 294–312. [Google Scholar] [CrossRef] [PubMed]
- Katyama, M.; Yoshimi, N.; Yamada, Y.; Sakata, K.; Kuno, T.; Yoshida, K.; Qiao, Z.; Vihn, P.Q.; Iwasaki, T.; Kobayashi, H.; et al. Preventive effect of fermented brown rice and rice bran against colon carcinogenesis in male F344 rats. Oncol. Rep. 2002, 9, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Katayama, M.; Sugie, S.; Yoshimi, N.; Yamada, Y.; Sakata, K.; Qiao, Z.; Iwasaki, T.; Kobayashi, H.; Mori, H. Preventive effect of fermented brown rice and rice bran on diethylnitrosoamine and phenobarbital-induced hepatocarcinogenesis in male F344 rats. Oncol. Rep. 2003, 10, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Kuno, T.; Hirose, Y.; Hata, K.; Kato, K.; Qiang, S.H.; Kitaori, N.; Hara, A.; Iwasaki, T.; Yoshimura, T.; Wada, K.; et al. Preventive effect of fermented brown rice and rice bran on N-nitrosomethylbenzylamine-induced esophageal tumorigenesis in rats. Int. J. Oncol. 2004, 25, 1809–1815. [Google Scholar] [CrossRef] [PubMed]
- Kuno, T.; Hirose, Y.; Yamada, Y.; Hata, K.; Qiang, S.H.; Asano, N.; Oyama, T.; Zhi, H.; Iwasaki, T.; Kobayashi, H.; et al. Chemoprevention of mouse urinary bladder carcinogenesis by fermented brown rice and rice bran. Oncol. Rep. 2006, 15, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Tomita, H.; Kuno, T.; Yamada, Y.; Oyama, T.; Asano, N.; Miyazaki, Y.; Baba, S.; Taguchi, A.; Hara, A.; Iwasaki, T.; et al. Preventive effect of fermented brown rice and rice bran on N-methyl-N′-nitro-N-nitrosoguanidine-induced gastric carcinogenesis in rats. Oncol. Rep. 2008, 19, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Phutthaphadoong, S.; Yamada, Y.; Hirata, A.; Tomita, H.; Taguchi, A.; Hara, A.; Limtrakul, P.N.; Iwasaki, T.; Kobayashi, H.; Mori, H. Chemopreventive effects of fermented brown rice and rice bran against 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced lung tumorigenesis in female A/J mice. Oncol. Rep. 2009, 21, 321–327. [Google Scholar] [PubMed]
- Long, N.K.; Makita, H.; Yamashita, T.; Toida, M.; Kato, K.; Hatakeyama, D.; Shibata, T. Chemopreventive effect of fermented brown rice and rice bran on 4-nitroquinoline 1-oxide-induced oral carcinogenesis in rats. Oncol. Rep. 2007, 17, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Phutthaphadoong, S.; Yamada, Y.; Hirata, A.; Tomita, H.; Hara, A.; Limtrakul, P.; Iwasaki, T.; Kobayashi, H.; Mori, H. Chemopreventive effect of fermented brown rice and rice bran (FBRA) on the inflammation-related colorectal carcinogenesis in ApcMin/+ mice. Oncol. Rep. 2010, 23, 53–59. [Google Scholar] [PubMed]
- Tazawa, H.; Okada, F.; Kobayashi, T.; Tada, M.; Mori, Y.; Une, Y.; Sendo, F.; Kobayashi, M.; Hosokawa, M. Infiltration of neutrophils is required for acquisition of metastatic phenotype of benign murine fibrosarcoma cells: Implication of inflammation-associated carcinogenesis and tumor progression. Am. J. Pathol. 2003, 163, 2221–2232. [Google Scholar] [CrossRef]
- Okada, F.; Hosokawa, M.; Hamada, J.I.; Hasegawa, J.; Kato, M.; Mizutani, M.; Ren, J.; Takeichi, N.; Kobayashi, H. Malignant progression of a mouse fibrosarcoma by host cells reactive to a foreign body (gelatin sponge). Br. J. Cancer 1992, 66, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Okada, F.; Kobayashi, M.; Tanaka, H.; Kobayashi, T.; Tazawa, H.; Iuchi, Y.; Onuma, K.; Hosokawa, M.; Dinauer, M.C.; Hunt, N.H. The role of nicotinamide adenine dinucleotide phosphate oxidase-derived reactive oxygen species in the acquisition of metastatic ability of tumor cells. Am. J. Pathol. 2006, 169, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Okada, F.; Tazawa, H.; Kobayashi, T.; Kobayashi, M.; Hosokawa, M. Involvement of reactive nitrogen oxides for acquisition of metastatic properties of benign tumors in a model of inflammation-based tumor progression. Nitric Oxide 2006, 14, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Okada, F.; Hamada, J.; Hosokawa, M.; Kobayashi, H. Changes in the tumorigenic and metastatic properties of tumor cells treated with quercetin or 5-azacytidine. Int. J. Cancer 1987, 39, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Okada, F.; Hosokawa, M.; Hasegawa, J.; Ishikawa, M.; Chiba, I.; Nakamura, Y.; Kobayashi, H. Regression mechanisms of mouse fibrosarcoma cells after in vitro exposure to quercetin: Diminution of tumorigenicity with a corresponding decrease in the production of prostaglandin E2. Cancer Immunol. Immunother. 1990, 31, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Onuma, K.; Suenaga, Y.; Sakaki, R.; Yoshitome, S.; Sato, Y.; Ogawara, S.; Suzuki, S.; Kuramitsu, Y.; Yokoyama, H.; Murakami, A.; et al. Development of a quantitative bioassay to assess preventive compounds against inflammation-based carcinogenesis. Nitric Oxide 2011, 25, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Okada, F. Inflammation-related carcinogenesis: Current findings in epidemiological trends, causes and mechanisms. Yonago Acta Med. 2014, 57, 65–72. [Google Scholar] [PubMed]
- Tanaka, T.; Kohno, H.; Suzuki, R.; Hata, K.; Sugie, S.; Niho, N.; Sakano, K.; Takahashi, M.; Wakabayashi, K. Dextran sodium sulfate strongly promotes colorectal carcinogenesis in ApcMin/+ mice: Inflammatory stimuli by dextran sodium sulfate results in development of multiple colonic neoplasms. Int. J. Cancer 2006, 118, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Strieter, R.M.; Standiford, T.J.; Huffnagle, G.B.; Colletti, L.M.; Lukacs, N.W.; Kunkel, S.L. “The good, the bad, and the ugly”. The role of chemokines in models of human disease. J. Immunol. 1996, 156, 3583–3586. [Google Scholar] [PubMed]
- Mazzone, A.; Ricevuti, G. Leukocyte CD11/CD18 integrins: Biological and clinical relevance. Haematologica 1995, 80, 161–175. [Google Scholar] [PubMed]
- Huang, S.; Paulauskis, J.D.; Godleski, J.J.; Kobzik, L. Expression of macrophage inflammatory protein-2 and KC mRNA in pulmonary inflammation. Am. J. Pathol. 1992, 141, 981–988. [Google Scholar] [PubMed]
- Driscoll, K.E.; Hassenbein, D.G.; Carter, J.; Poynter, J.; Asquith, T.N.; Grant, R.A.; Whitten, J.; Purdon, M.P.; Takigiku, R. Macrophage inflammatory proteins 1 and 2: Expression by rat alveolar macrophages, fibroblasts, and epithelial cells and in rat lung after mineral dust exposure. Am. J. Respir. Cell Mol. Biol. 1993, 8, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Tessier, P.A.; Naccache, P.H.; Clark-Lewis, I.; Gladue, R.P.; Neote, K.S.; McColl, S.R. Chemokine networks in vivo: Involvement of C-X-C and C-C chemokines in neutrophil extravasation in vivo in response to TNF-alpha. J. Immunol. 1997, 159, 3595–3602. [Google Scholar] [PubMed]
- Liu, Q.; Wang, Y.; Thorlacius, H. Dexamethasone inhibits tumor necrosis factor-alpha-induced expression of macrophage inflammatory protein-2 and adhesion of neutrophils to endothelial cells. Biochem. Biophys. Res. Commun. 2000, 271, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Pichyangkul, S.; Schick, D.; Schober, W.; Dixon, G.; Khan, A. Increased expression of adhesive proteins on leukocytes by TNF alpha. Exp. Hematol. 1998, 16, 588–593. [Google Scholar]
- Okada, F.; Shionoya, H.; Kobayashi, M.; Kobayashi, T.; Tazawa, H.; Onuma, K.; Iuchi, Y.; Matsubara, N.; Ijichi, T.; Dugas, B.; et al. Prevention of inflammation-mediated acquisition of metastatic properties of benign mouse fibrosarcoma cells by administration of an orally available superoxide dismutase. Br. J. Cancer 2006, 94, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, H.; Ikata, K.; Arimochi, H.; Iwasaki, T.; Ohnishi, Y.; Kuwahara, T.; Kataoka, K. Effects of fermented brown rice on the intestinal environments in healthy adult. J. Med. Investig. 2011, 58, 235–245. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onuma, K.; Kanda, Y.; Suzuki Ikeda, S.; Sakaki, R.; Nonomura, T.; Kobayashi, M.; Osaki, M.; Shikanai, M.; Kobayashi, H.; Okada, F. Fermented Brown Rice and Rice Bran with Aspergillus oryzae (FBRA) Prevents Inflammation-Related Carcinogenesis in Mice, through Inhibition of Inflammatory Cell Infiltration. Nutrients 2015, 7, 10237-10250. https://doi.org/10.3390/nu7125531
Onuma K, Kanda Y, Suzuki Ikeda S, Sakaki R, Nonomura T, Kobayashi M, Osaki M, Shikanai M, Kobayashi H, Okada F. Fermented Brown Rice and Rice Bran with Aspergillus oryzae (FBRA) Prevents Inflammation-Related Carcinogenesis in Mice, through Inhibition of Inflammatory Cell Infiltration. Nutrients. 2015; 7(12):10237-10250. https://doi.org/10.3390/nu7125531
Chicago/Turabian StyleOnuma, Kunishige, Yusuke Kanda, Saori Suzuki Ikeda, Ryuta Sakaki, Takuya Nonomura, Masanobu Kobayashi, Mitsuhiko Osaki, Masataka Shikanai, Hiroshi Kobayashi, and Futoshi Okada. 2015. "Fermented Brown Rice and Rice Bran with Aspergillus oryzae (FBRA) Prevents Inflammation-Related Carcinogenesis in Mice, through Inhibition of Inflammatory Cell Infiltration" Nutrients 7, no. 12: 10237-10250. https://doi.org/10.3390/nu7125531
APA StyleOnuma, K., Kanda, Y., Suzuki Ikeda, S., Sakaki, R., Nonomura, T., Kobayashi, M., Osaki, M., Shikanai, M., Kobayashi, H., & Okada, F. (2015). Fermented Brown Rice and Rice Bran with Aspergillus oryzae (FBRA) Prevents Inflammation-Related Carcinogenesis in Mice, through Inhibition of Inflammatory Cell Infiltration. Nutrients, 7(12), 10237-10250. https://doi.org/10.3390/nu7125531