Anti-Obesity Effect of 6,8-Diprenylgenistein, an Isoflavonoid of Cudrania tricuspidata Fruits in High-Fat Diet-Induced Obese Mice


Abstract
:1. Introduction

2. Experimental Section
2.1. Chemicals
2.2. Preparation of DPG
2.3. Animal Treatment
2.4. Serum Biochemical Parameters
2.5. Histopathology
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
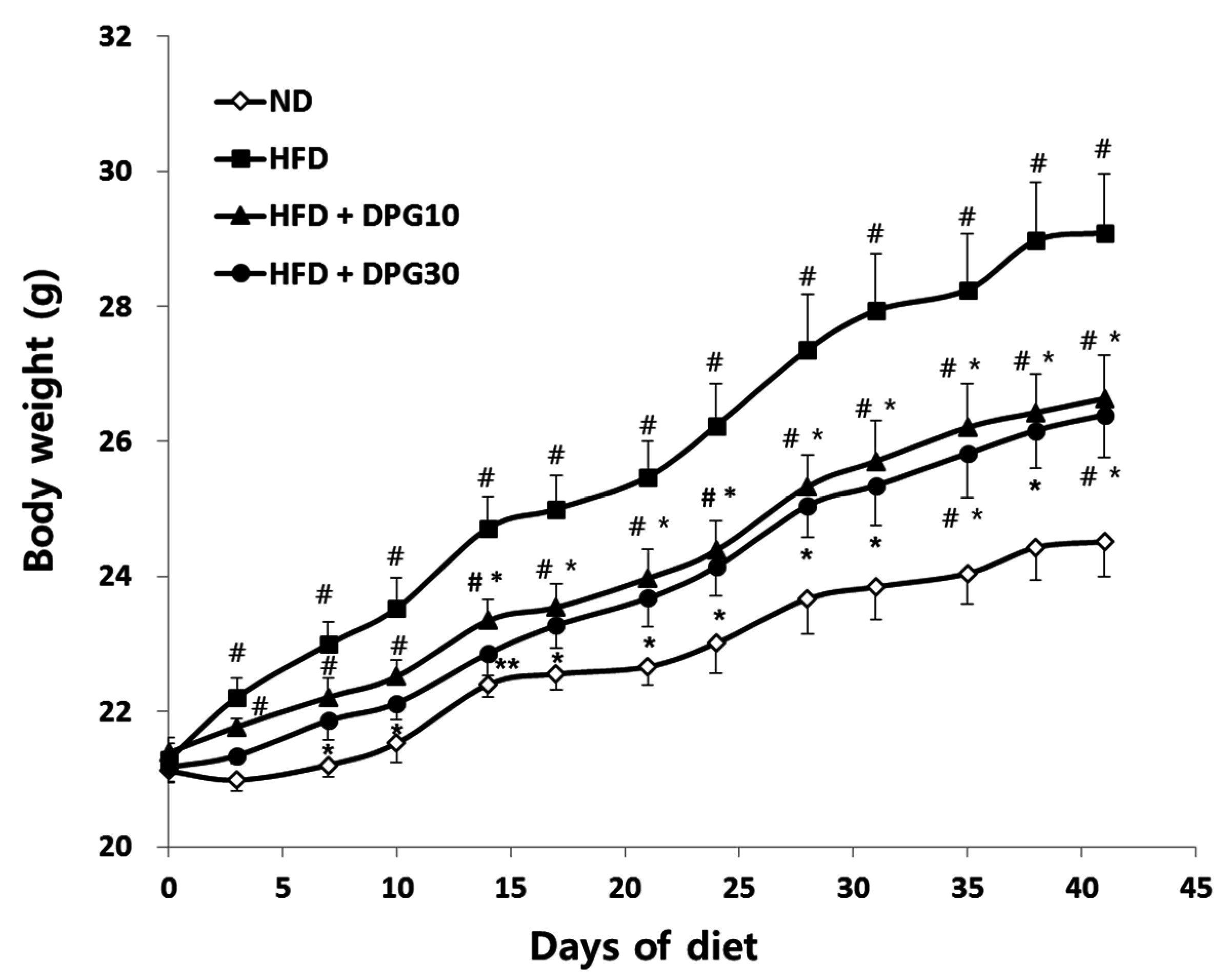
3.1. Effects of DPG on Body Weight and Food Efficiency Ratio


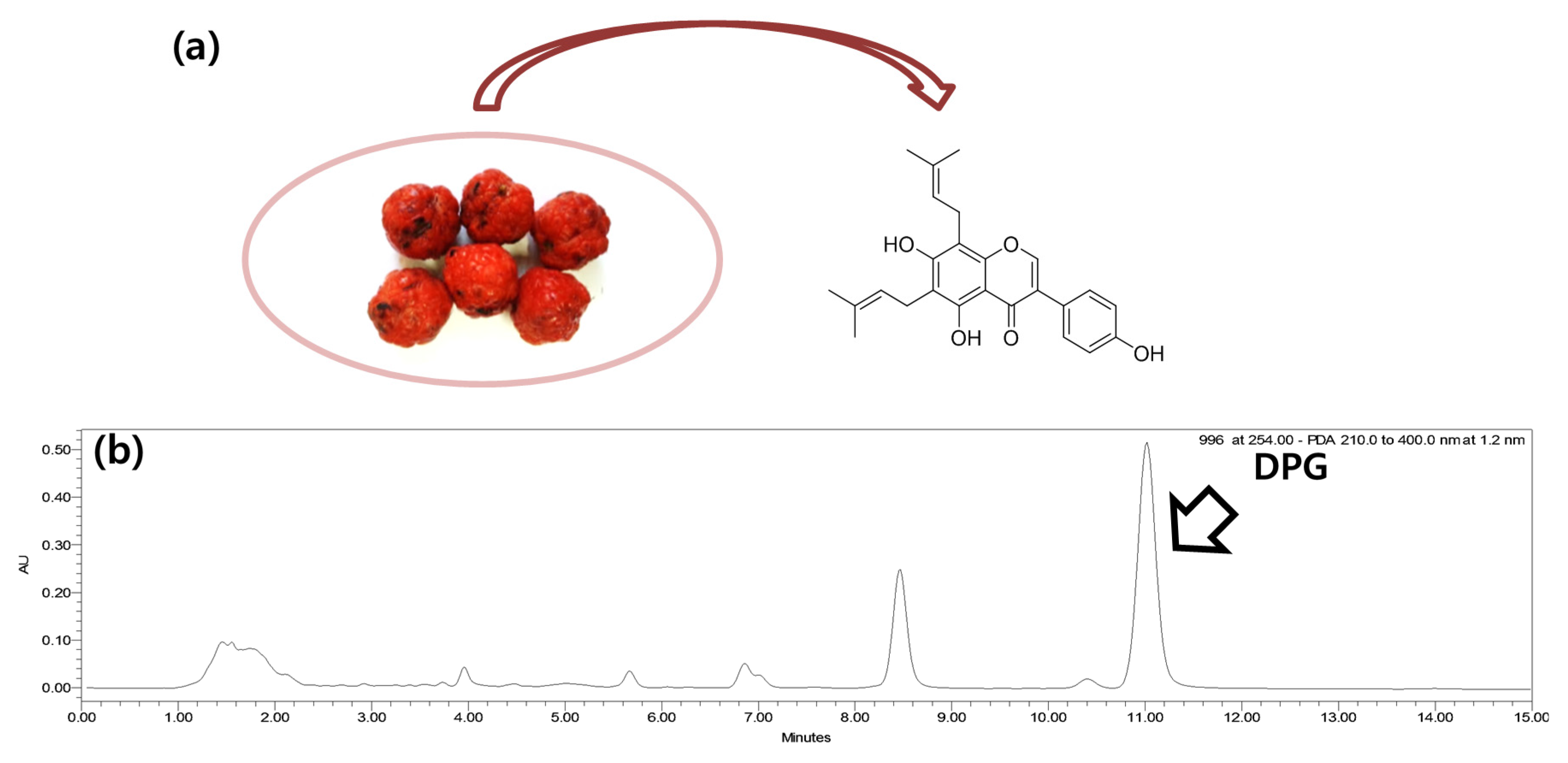{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body Weight Gain (g/Mice/6 Weeks) | Food Intake (g/Day/Mice) | FER 1 |
|---|---|---|---|
| ND | 3.39 ± 0.48 | 3.49 ± 0.03 | 0.023 ± 0.003 |
| HFD | 7.81 ± 0.98 ## | 2.71 ± 0.03 ## | 0.069 ± 0.008 ## |
| HFD + DPG10 | 5.25 ± 0.65 #,* | 2.73 ± 0.04 ## | 0.046 ± 0.005 ##,* |
| HFD + DPG30 | 5.13 ± 0.50 # * | 2.74 ± 0.09 ## | 0.044 ± 0.004 ##,* |
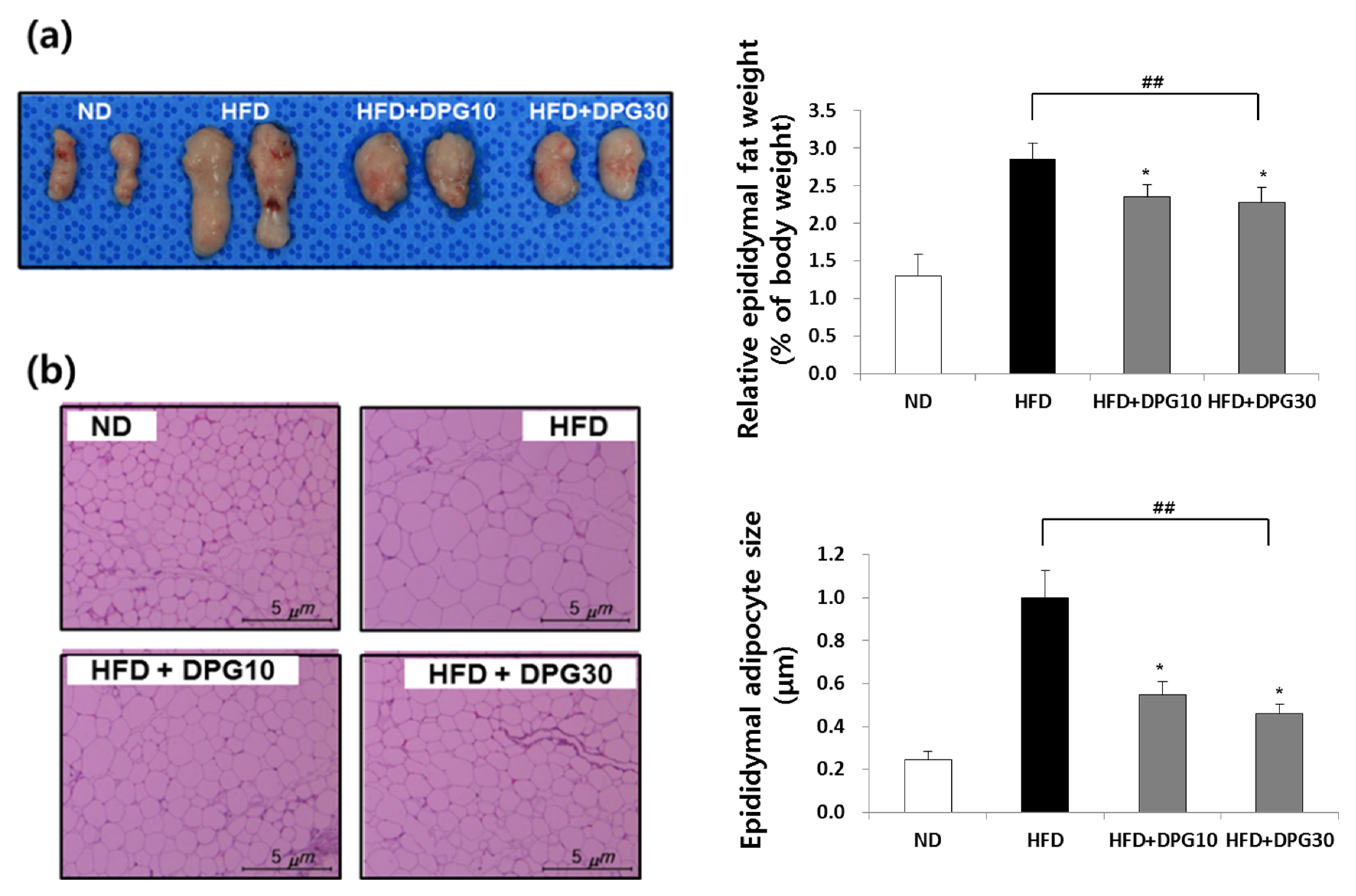
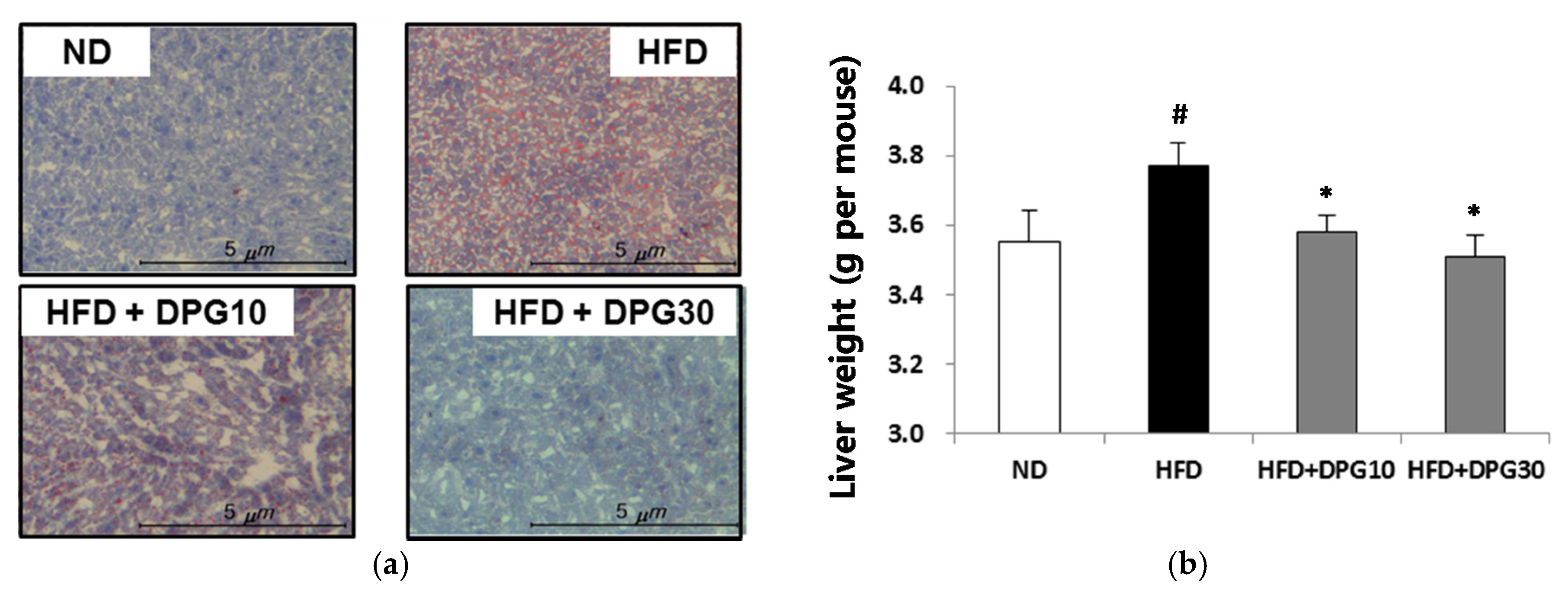
3.2. Effect of DPG on Fat Accumulation


3.3. Effect of DPG on Serum Lipid Profiles
| Group | ND | HFD | HFD + DPG10 | HFD + DPG30 |
|---|---|---|---|---|
| AST 1 | 46.54 ± 5.55 | 58.85 ± 8.83 | 53.22 ± 5.21 | 50.48 ± 4.02 ** |
| ALT 1 | 20.13 ± 2.85 | 24.25 ± 3.35 | 21.18 ± 2.14 * | 20.54 ± 2.98 ** |
| Total cholesterol 2 | 101.95 ± 10.46 | 151.25 ± 14.02 | 135.78 ± 13.44 * | 130.15 ± 15.20 ** |
| HDL cholesterol 2 | 62.61 ± 7.41 | 83.03 ± 6.63 | 76.19 ± 6.75 * | 75.75 ± 6.87 * |
| LDL cholesterol 2 | 4.60 ± 0.82 | 8.17 ± 1.31 | 6.77 ± 1.41 * | 6.45 ± 1.38 * |
| Triglyceride 2 | 30.71 ± 10.13 | 50.55 ± 13.71 | 35.19 ± 10.65 * | 33.72 ± 7.48 ** |
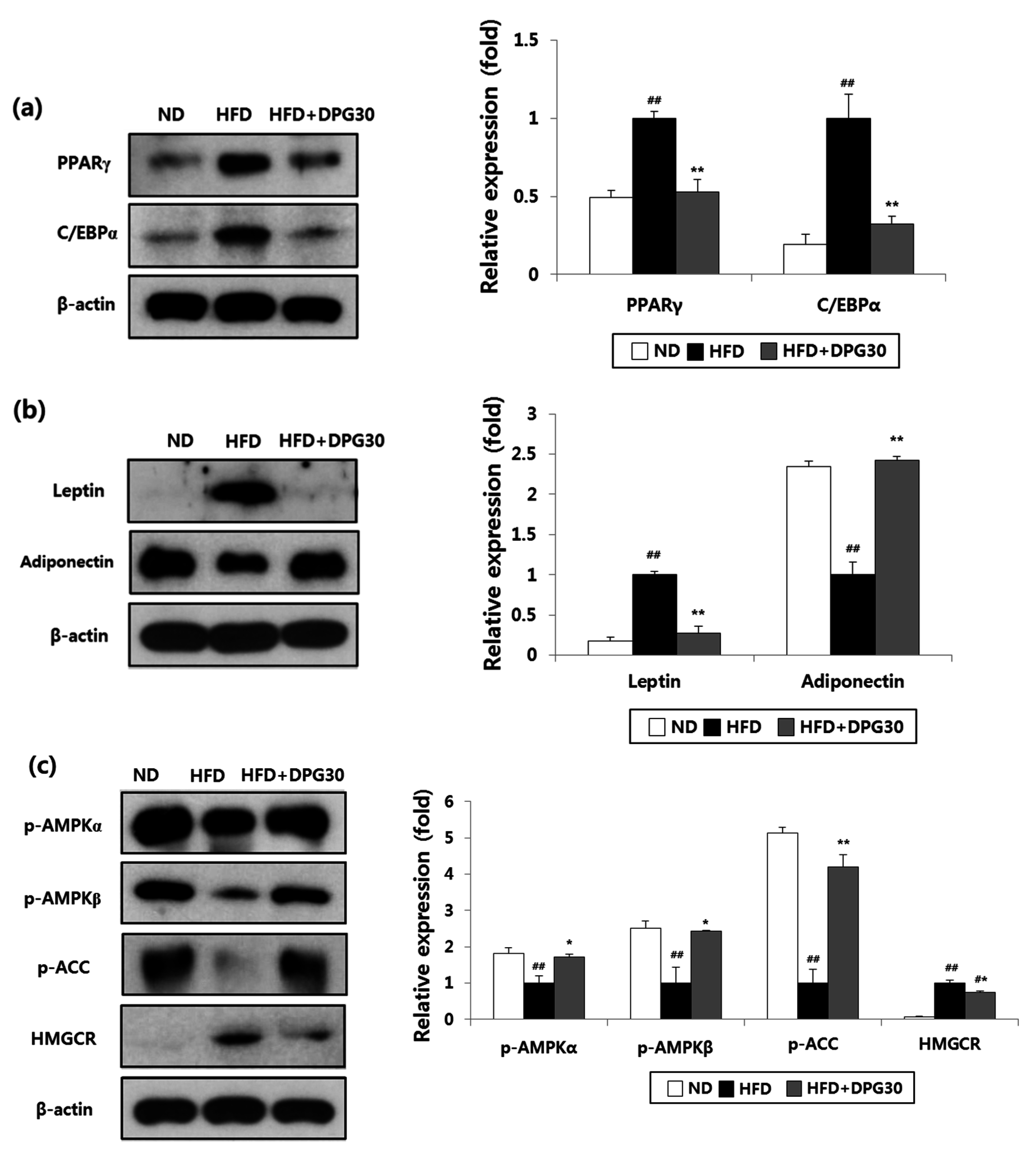
3.4. Effect of DPG on Adipogenesis in Adipose Tissue

3.5. Effect of DPG on AMPK Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [PubMed]
- Visscher, T.L.; Seidell, J.C. The public health impact of obesity. Ann. Rev. Public Health 2001, 22, 355–375. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.W. Possible anti-obesity therapeutics from nature—A review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef] [PubMed]
- Farmer, S.R. Transcriptional control of adipocyte formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.; Halaas, J.; Ravussin, E.; Pratley, R.E.; Lee, G.H.; Zhang, Y. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Ruderman, N.B.; Carling, D.; Prentki, M.; Cacicedo, J.M. AMPK, insulin resistance, and the metabolic syndrome. J. Clin. Investig. 2013, 123, 2764–2772. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. Regulation of fatty acid and cholesterol metabolism by the AMP-activated protein kinase. Biochim. Biophys. Acta 1992, 1123, 231–238. [Google Scholar] [CrossRef]
- Legeay, S.; Rodier, M.; Fillon, L.; Faure, S.; Clere, N. Epigallocatechin gallate: A review of its beneficial properties to prevent metabolic syndrome. Nutrients 2015, 7, 5443–5468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.M.; Jeon, Y.S.; Kim, W.; Lee, A.; Kim, Y.G.; Lee, J.H.; Kang, Y.E.; Jung, J.C.; Lee, J.; Min, B.; et al. Xanthigen attenuates high-fat diet-induced obesity through down-regulation of PPARγ and activation of the AMPK pathway. Food Sci. Biotechnol. 2014, 23, 931–935. [Google Scholar] [CrossRef]
- Choi, K.M.; Lee, Y.S.; Kim, W.; Kim, S.J.; Shin, K.O.; Yu, J.Y.; Lee, M.K.; Lee, Y.M.; Hong, J.T.; Yun, Y.P.; et al. Sulforaphane attenuates obesity by inhibiting adipogenesis and activating the AMPK pathway in obese mice. J. Nutr. Biochem. 2014, 25, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Kim, E.S.; Lee, C.; Kim, S.; Cho, S.H.; Hwang, B.Y.; Lee, M.K. Chemical constituents from Nelumbo nucifera leaves and their anti-obesity effects. Bioorg. Med. Chem. Lett. 2013, 23, 3604–3608. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, T.; Melzig, M.F. Polyphenolic compounds as pancreatic lipase inhibitors. Planta Med. 2015, 81, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Senaphan, K.; Kukongviriyapan, U.; Sangartit, W.; Pakdeechote, P.; Pannangpetch, P.; Prachaney, P.; Greenwald, S.E.; Kukongviriyapan, V. Ferulic acid alleviates changes in a rat model of metabolic syndrome induced by high-carbohydrate, high-fat diet. Nutrients 2015, 7, 6446–6464. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Kiyosawa, T.; Nakamura, S.; Osada, K. Effects of isoflavone supplementation on disturbances in lipid metabolism and antioxidant system due to exogenous cholesterol oxidation products in rats. J. Funct. Foods 2014, 7, 212–218. [Google Scholar] [CrossRef]
- Lee, H.; Ha, H.; Lee, J.K.; Seo, C.S.; Lee, N.H.; Jung, D.Y.; Park, S.J.; Shin, H.K. The fruits of Cudrania tricuspidata suppress development of atopic dermatitis in NC/Nga mice. Phytother. Res. 2012, 26, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Han, X.H.; Hong, S.S.; Jin, Q.; Li, D.; Kim, H.K.; Lee, J.; Kwon, S.H.; Lee, D.; Lee, C.K.; Lee, M.K.; et al. Prenylated and benzylated flavonoids from the fruits of Cudrania tricuspidata. J. Nat. Prod. 2009, 72, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.J.; Yokozawa, T.; Rhyu, D.Y.; Kim, S.C.; Shibahara, N.; Park, J.C. Study on the inhibitory effects of Korean medicinal plants and their main compounds on the 1,1-diphenyl-2-picrylhydrazyl radical. Phytomedicine 2003, 10, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.Y.; Jo, Y.H.; Lee, K.Y.; Do, S.G.; Hwang, B.Y.; Lee, M.K. Optimization of pancreatic lipase inhibition by Cudrania tricuspidata fruits using response surface methodology. Bioorg. Med. Chem. Lett. 2014, 24, 2329–2333. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.K.; Lee, C.H.; Seo, J.H.; Chung, M.Y.; Cui, L.; Fomum, Z.T.; Kang, J.S.; Lee, H.S. Diacylglycerol acyltransferase-inhibitory compounds from Erythrina senegalensis. Arch. Pharm. Res. 2009, 32, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.D.; Greenspan, P. Application of Nile red, a fluorescent hydrophobic probe, for the detection of neutral lipid deposits in tissue sections: Comparison with oil red O. J. Histochem. Cytochem. 1985, 33, 833–836. [Google Scholar] [CrossRef] [PubMed]
- White, U.A.; Stephens, J.M. Transcriptional factors that promote formation of white adipose tissue. Mol. Cell Endocrinol. 2010, 318, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Rajala, M.W.; Scherer, P.E. Minireview: The adipocyte-at the crossroads of energy homeostasis, inflammation, and atherosclerosis. Endocrinology 2003, 144, 3765–3773. [Google Scholar] [CrossRef] [PubMed]
- Tobert, J.A. Lovastatin and beyond: The history of the HMG-CoA reductase inhibitors. Nat. Rev. Drug Discov. 2003, 2, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.M.; Lee, Y.S.; Shin, D.M.; Lee, S.; Yoo, K.S.; Lee, M.K.; Lee, J.H.; Kim, S.Y.; Lee, Y.M.; Hong, J.T.; et al. Green tomato extract attenuates high-fat-diet-induced obesity through activation of the AMPK pathway in C57BL/6 mice. J. Nutr. Biochem. 2013, 24, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Bhaswant, M.; Poudyal, H.; Mathai, M.L.; Ward, L.C.; Mouatt, P.; Brown, L. Green and black cardamom in a diet-induced rat model of metabolic syndrome. Nutrients 2015, 7, 7691–7707. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, L.; Fernández-Quintela, A.; Arias, N.; Portillo, P.P. Resveratrol: Anti-obesity mechanisms of action. Molecules 2014, 19, 18632–18655. [Google Scholar] [CrossRef] [PubMed]
- Okuda, M.H.; Zemdegs, J.C.S.; Santana, A.A.; Santamarina, A.B.; Moreno, M.F.; Hachul, A.C.L.; Santos, B.; Nascimento, O.; Ribeiro, E.B.; Oyama, L.M. Green tea extract improves high fat diet-induced hypothalamic inflammation, without affecting serotoninergic system. J. Nutr. Biochem. 2014, 25, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Sclafani, A.; Springer, D. Dietary obesity in adult rats: Similarities to hypothalamic and human obesity syndromes. Physiol. Behav. 1976, 17, 461–471. [Google Scholar] [CrossRef]
- Ntambi, J.M.; Young-Cheul, K. Adipocyte differentiation and gene expression. J. Nutr. 2000, 130, 3122S–3126S. [Google Scholar] [PubMed]
- Tong, Q.; Hotamisligil, G.S. Molecular mechanisms of adipocyte differentiation. Rev. Endocr. Metab. Disord. 2001, 2, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Nedvidkova, J.; Smitka, K.; Kopsky, V.; Hainer, V. Adiponectin, an adipocyte-derived protein. Physiol. Res. 2005, 54, 133–140. [Google Scholar] [PubMed]
- Diez, J.J.; Iglesias, P. The role of the novel adipocyte-derived hormone adiponectin in human disease. Eur. J. Endocrinol. 2003, 148, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Carling, D.; Zammit, V.A.; Hardie, D.G. A common bicyclic protein kinase cascade inactivates the regulatory enzymes of fatty acid and cholesterol biosynthesis. FEBS Lett. 1987, 223, 217–222. [Google Scholar] [CrossRef]
- Steinberg, G.R.; Kemp, B.E. AMPK in health and disease. Physiol. Rev. 2008, 89, 1025–1078. [Google Scholar] [CrossRef] [PubMed]
- Behloul, N.; Wu, G. Genistein: A promising therapeutic agent for obesity and diabetes treatment. Eur. J. Pharmacol. 2013, 698, 31–38. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, Y.H.; Choi, K.-M.; Liu, Q.; Kim, S.B.; Ji, H.-J.; Kim, M.; Shin, S.-K.; Do, S.-G.; Shin, E.; Jung, G.; et al. Anti-Obesity Effect of 6,8-Diprenylgenistein, an Isoflavonoid of Cudrania tricuspidata Fruits in High-Fat Diet-Induced Obese Mice. Nutrients 2015, 7, 10480-10490. https://doi.org/10.3390/nu7125544
Jo YH, Choi K-M, Liu Q, Kim SB, Ji H-J, Kim M, Shin S-K, Do S-G, Shin E, Jung G, et al. Anti-Obesity Effect of 6,8-Diprenylgenistein, an Isoflavonoid of Cudrania tricuspidata Fruits in High-Fat Diet-Induced Obese Mice. Nutrients. 2015; 7(12):10480-10490. https://doi.org/10.3390/nu7125544
Chicago/Turabian StyleJo, Yang Hee, Kyeong-Mi Choi, Qing Liu, Seon Beom Kim, Hyeong-Jin Ji, Myounghwan Kim, Sang-Kyung Shin, Seon-Gil Do, Eunju Shin, Gayoung Jung, and et al. 2015. "Anti-Obesity Effect of 6,8-Diprenylgenistein, an Isoflavonoid of Cudrania tricuspidata Fruits in High-Fat Diet-Induced Obese Mice" Nutrients 7, no. 12: 10480-10490. https://doi.org/10.3390/nu7125544
APA StyleJo, Y. H., Choi, K. -M., Liu, Q., Kim, S. B., Ji, H. -J., Kim, M., Shin, S. -K., Do, S. -G., Shin, E., Jung, G., Yoo, H. -S., Hwang, B. Y., & Lee, M. K. (2015). Anti-Obesity Effect of 6,8-Diprenylgenistein, an Isoflavonoid of Cudrania tricuspidata Fruits in High-Fat Diet-Induced Obese Mice. Nutrients, 7(12), 10480-10490. https://doi.org/10.3390/nu7125544