Nutritional Ingredients Modulate Adipokine Secretion and Inflammation in Human Primary Adipocytes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation and Handling of the Nutritional Factors
2.2.1. Profile of the Extensive Casein Hydrolysate (eCH)


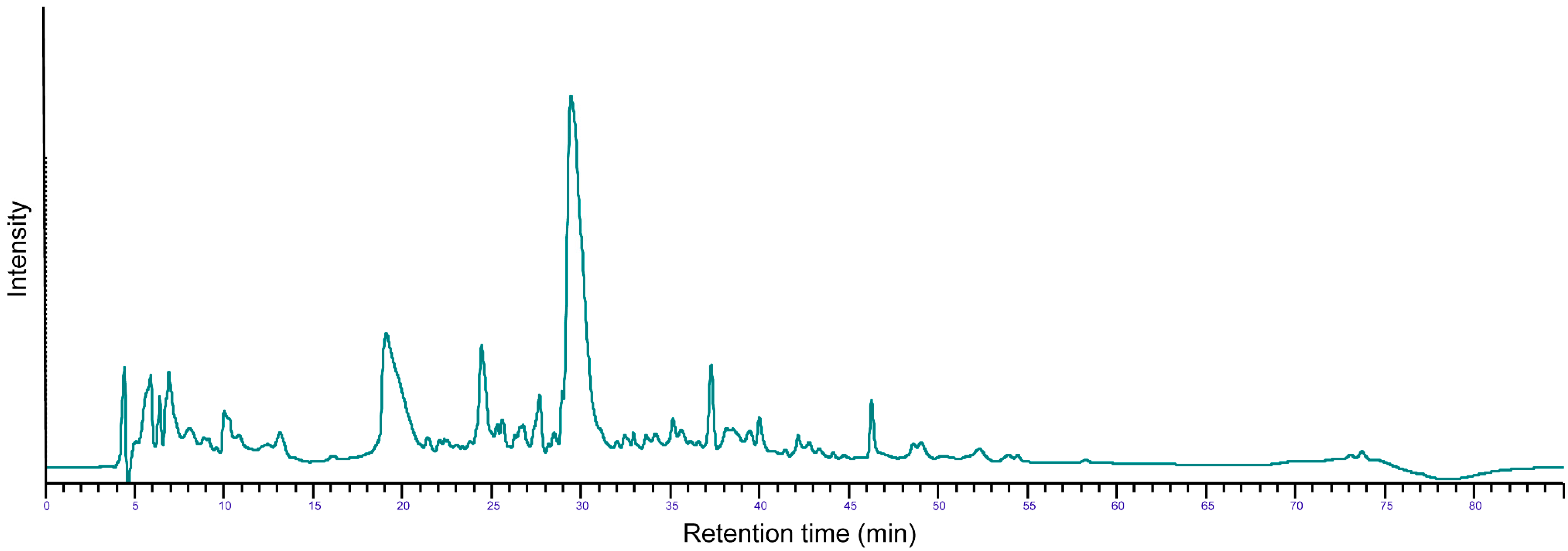{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide length | Relative signal (%) |
|---|---|
| 1–2 | 44.0 |
| 2–3 | 17.4 |
| 3–4 | 9.7 |
| 4–5 | 7.9 |
| 5–6 | 5.2 |
| 6–7 | 4.5 |
| 7–8 | 3.0 |
| 8–9 | 1.9 |
| 9–10 | 1.7 |
| 10–11 | 1.1 |
| 11–12 | 1.1 |
| 12–13 | 0.9 |
| 13–14 | 0.5 |
| 14–15 | 0.5 |
| 15–16 | 0.6 |
2.2.2. Preparation and Handling of LC-PUFAs
2.3. Isolation and Culture of Primary Human Preadipocytes and Experimental Design
2.4. LDH Activity
2.5. Neutral Red Staining
2.6. Oil Red O Staining
2.7. Western Blot
2.8. Adiponectin, Leptin, and Monocyte Chemoattractant Protein-1 (MCP-1) Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Data Analysis
3. Results
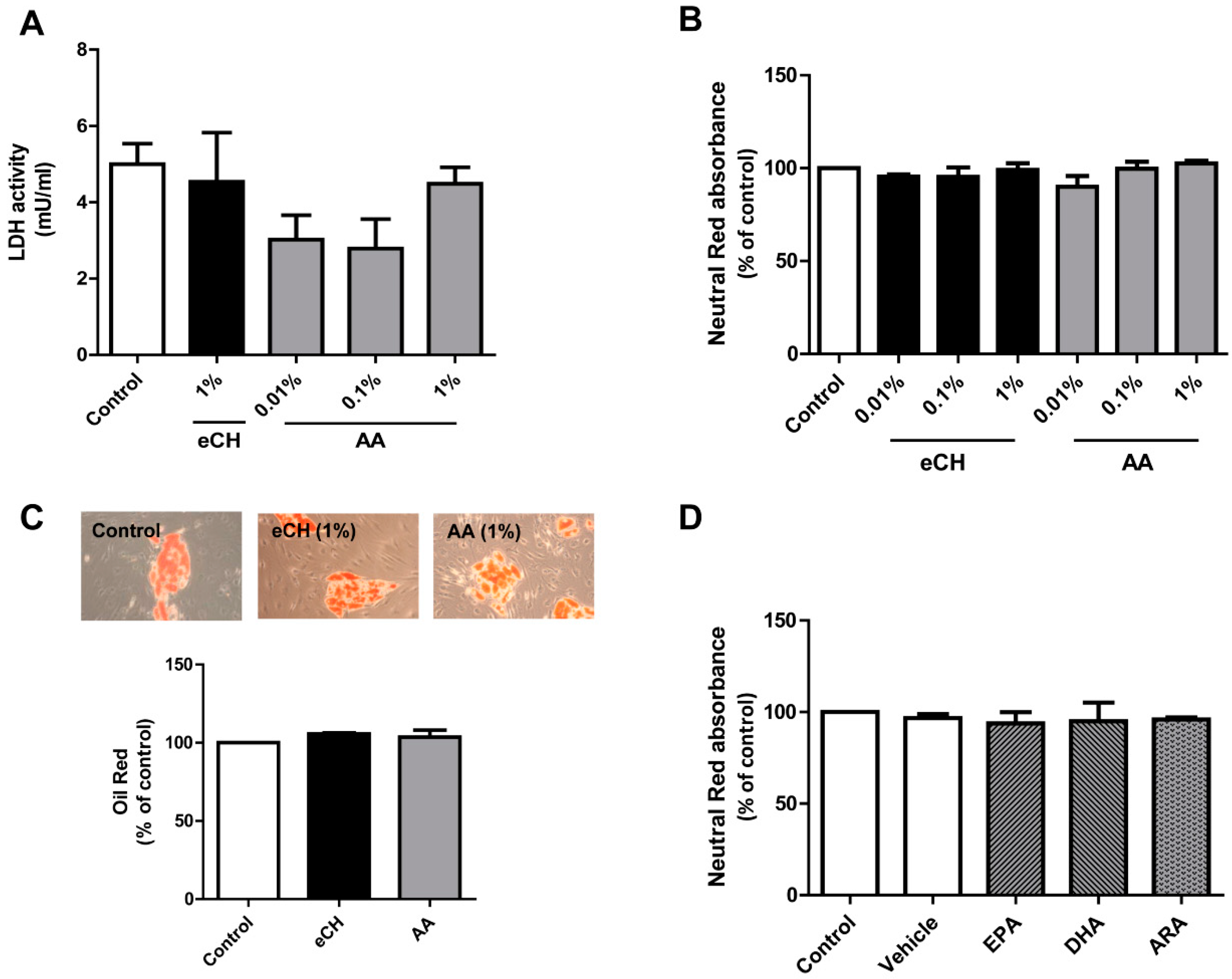
3.1. eCH and AA Do Not Alter Cell Viability or Morphology
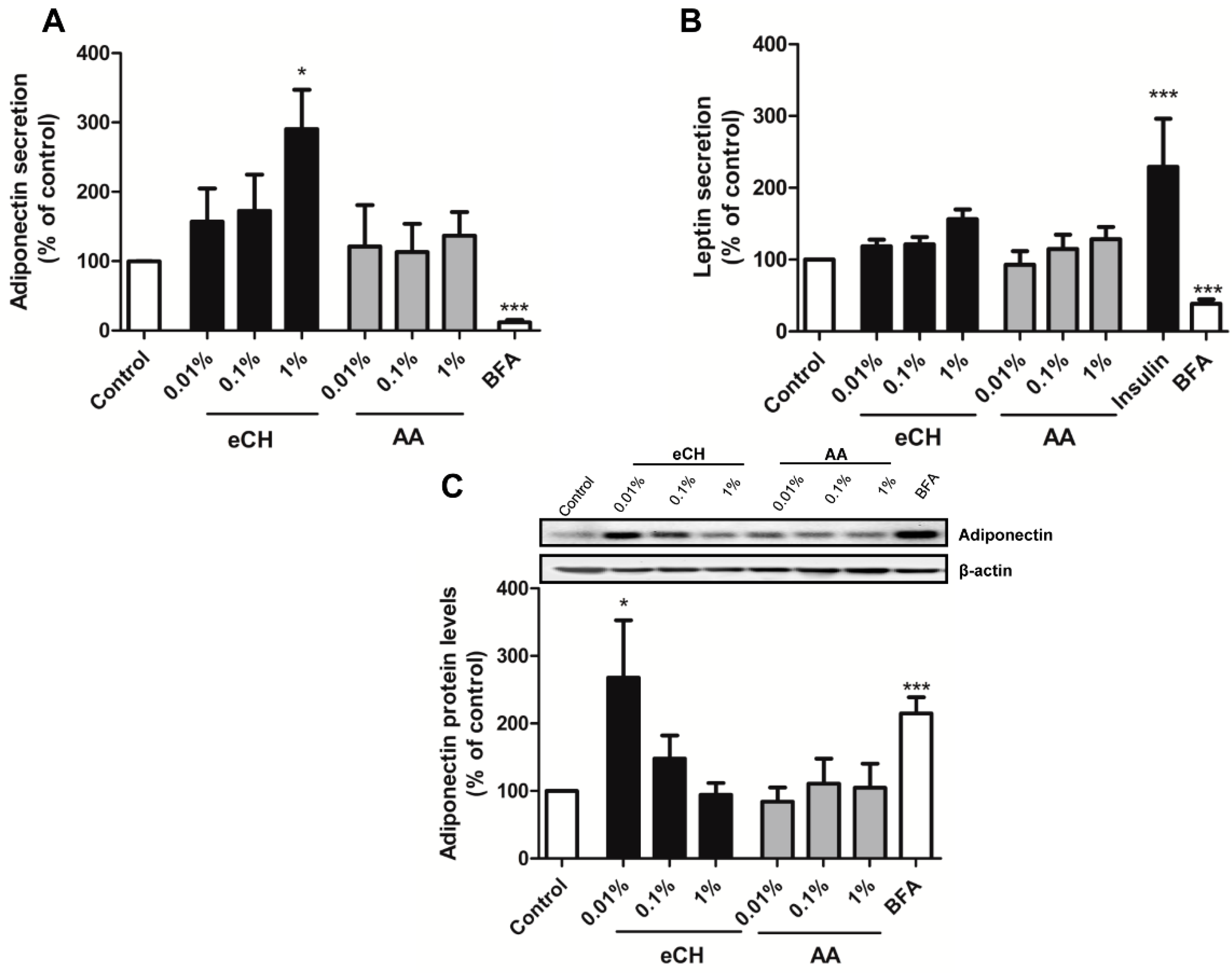
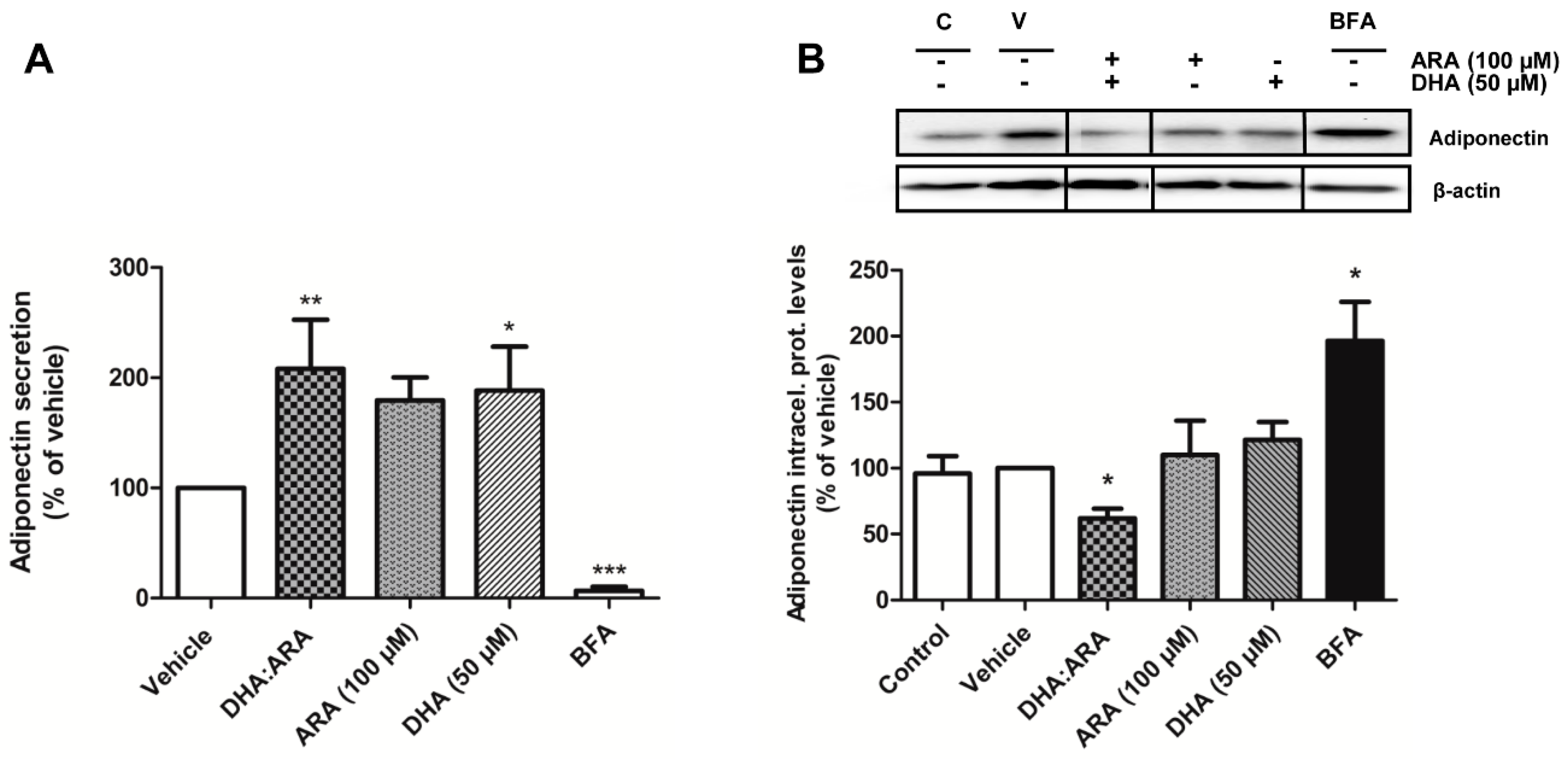
3.2. eCH Upregulates Adiponectin Secretion in a Concentration-Dependent Manner

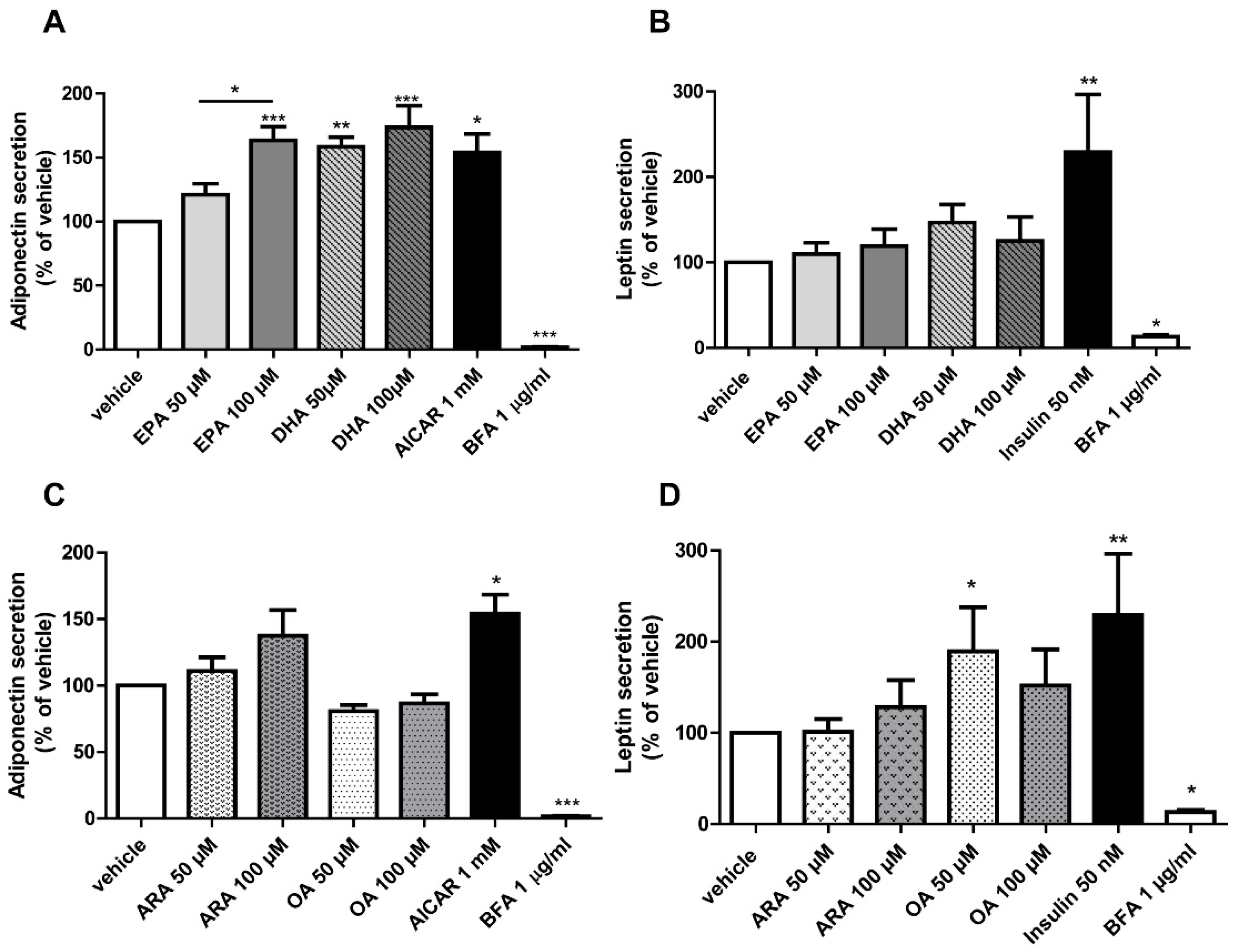
3.3. EPA and DHA Upregulate Adiponectin Secretion



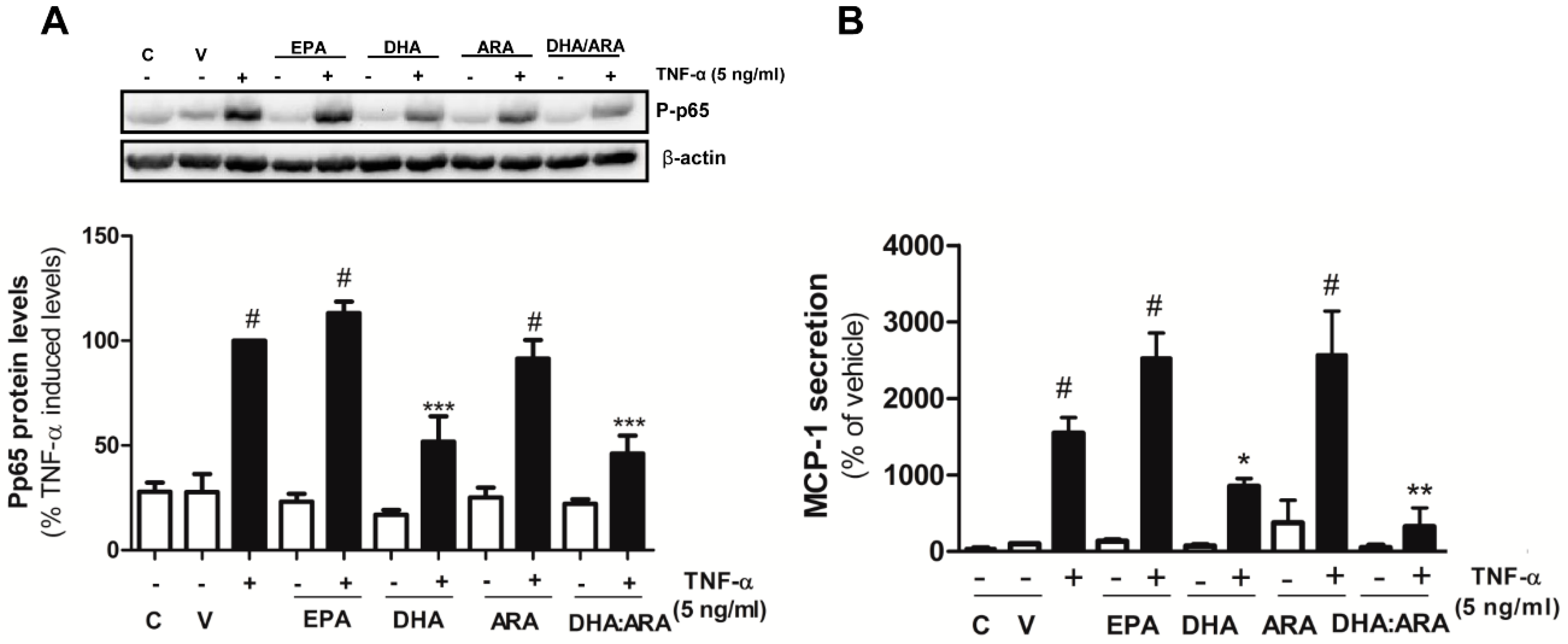
3.4. The DHA/ARA Combination Exerts a Prominent Anti-Inflammatory Effect

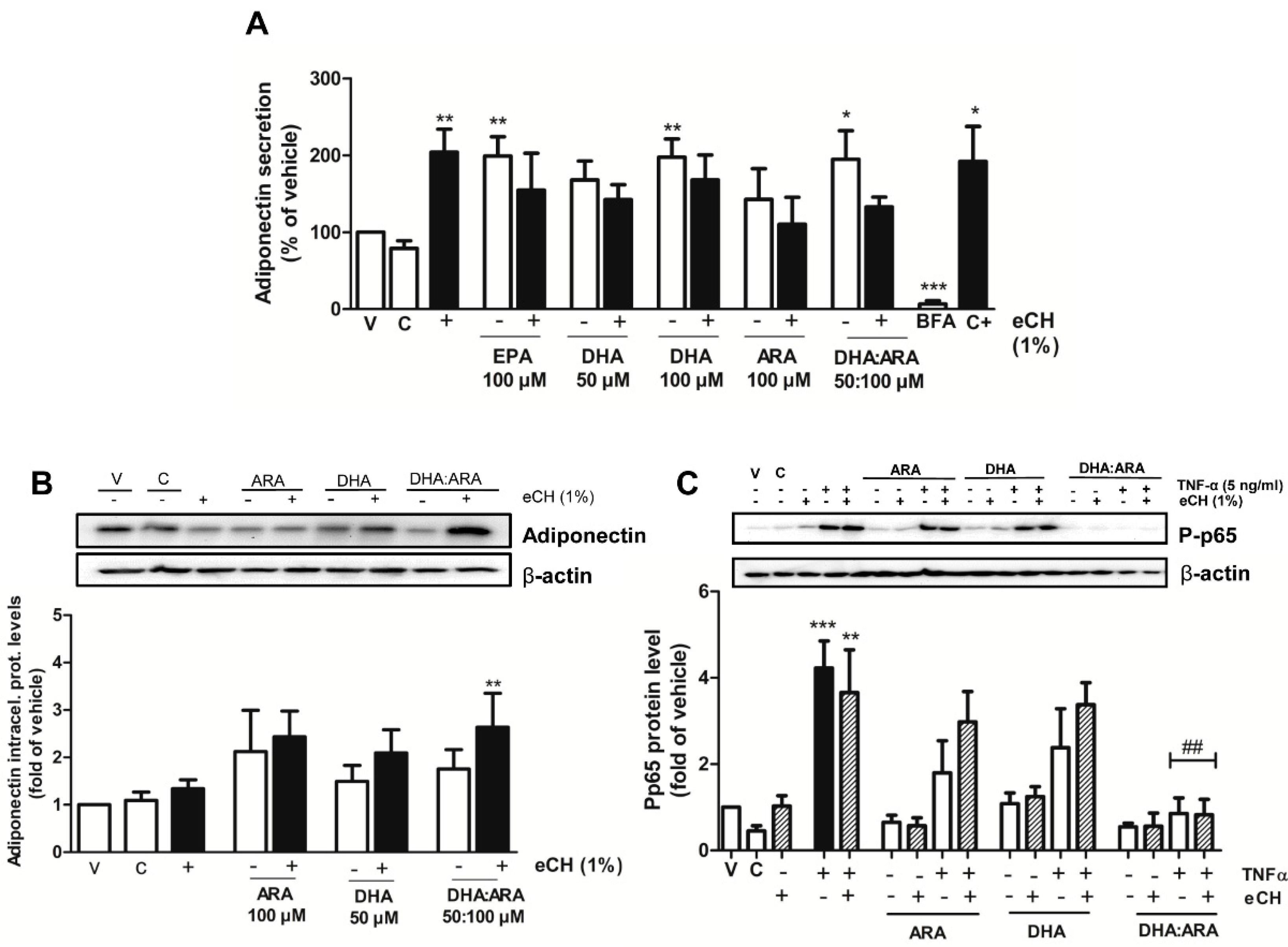
3.5. eCH Upregulates Intracellular Adiponectin Expression in Combination with DHA

4. Discussion
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Goran, M.I.; Treuth, M.S. Energy expenditure, physical activity, and obesity in children. Pediatr. Clin. North Am. 2001, 48, 931–953. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.S.; Hosking, J.; Metcalf, B.S.; Jeffery, A.N.; Voss, L.D.; Wilkin, T.J. Contribution of early weight gain to childhood overweight and metabolic health: A longitudinal study (earlybird 36). Pediatrics 2009, 123, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J. Obesity and early life. Obes. Rev. 2007, 8 (Suppl. 1), 45–49. [Google Scholar] [CrossRef]
- McMillen, I.C.; Robinson, J.S. Developmental origins of the metabolic syndrome: Prediction, plasticity, and programming. Physiol. Rev. 2005, 85, 571–633. [Google Scholar] [CrossRef] [PubMed]
- Miralles, O.; Sanchez, J.; Palou, A.; Pico, C. A physiological role of breast milk leptin in body weight control in developing infants. Obesity (Silver Spring) 2006, 14, 1371–1377. [Google Scholar] [CrossRef]
- Power, C.; Parsons, T. Nutritional and other influences in childhood as predictors of adult obesity. Proc. Nutr. Soc. 2000, 59, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Romacho, T.; Elsen, M.; Rohrborn, D.; Eckel, J. Adipose tissue and its role in organ crosstalk. Acta Physiol. (Oxf.) 2014, 210, 733–753. [Google Scholar] [CrossRef]
- Arner, P. Insulin resistance in type 2 diabetes—Role of the adipokines. Curr. Mol. Med. 2005, 5, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Shimomura, I.; Kishida, K.; Nishizawa, H.; Matsuda, M.; Nagaretani, H.; Furuyama, N.; Kondo, H.; Takahashi, M.; Arita, Y.; et al. Diet-induced insulin resistance in mice lacking adiponectin/acrp30. Nat. Med. 2002, 8, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P.; Wood, I.S. Adipokines: Inflammation and the pleiotropic role of white adipose tissue. Br. J. Nutr. 2004, 92, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Savino, F.; Liguori, S.A.; Fissore, M.F.; Oggero, R. Breast milk hormones and their protective effect on obesity. Int. J. Pediatr. Endocrinol. 2009, 2009, 327505. [Google Scholar] [CrossRef] [PubMed]
- Lillefosse, H.H.; Tastesen, H.S.; Du, Z.Y.; Ditlev, D.B.; Thorsen, F.A.; Madsen, L.; Kristiansen, K.; Liaset, B. Hydrolyzed casein reduces diet-induced obesity in male c57bl/6j mice. J. Nutr. 2013, 143, 1367–1375. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T.; Varamini, B.; Jensen, R.G.; Diersen-Schade, D.A.; Boettcher, J.A.; Arterburn, L.M. Docosahexaenoic and arachidonic acid concentrations in human breast milk worldwide. Am. J. Clin. Nutr. 2007, 85, 1457–1464. [Google Scholar] [PubMed]
- Birch, E.E.; Garfield, S.; Castaneda, Y.; Hughbanks-Wheaton, D.; Uauy, R.; Hoffman, D. Visual acuity and cognitive outcomes at 4 years of age in a double-blind, randomized trial of long-chain polyunsaturated fatty acid-supplemented infant formula. Early Hum. Dev. 2007, 83, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Flachs, P.; Rossmeisl, M.; Bryhn, M.; Kopecky, J. Cellular and molecular effects of n-3 polyunsaturated fatty acids on adipose tissue biology and metabolism. Clin. Sci. (Lond.) 2009, 116, 1–16. [Google Scholar] [CrossRef]
- Kopecky, J.; Rossmeisl, M.; Flachs, P.; Kuda, O.; Brauner, P.; Jilkova, Z.; Stankova, B.; Tvrzicka, E.; Bryhn, M. N-3 PUFA: Bioavailability and modulation of adipose tissue function. Proc. Nutr. Soc. 2009, 68, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Kuda, O.; Jelenik, T.; Jilkova, Z.; Flachs, P.; Rossmeisl, M.; Hensler, M.; Kazdova, L.; Ogston, N.; Baranowski, M.; Gorski, J.; et al. N-3 fatty acids and rosiglitazone improve insulin sensitivity through additive stimulatory effects on muscle glycogen synthesis in mice fed a high-fat diet. Diabetologia 2009, 52, 941–951. [Google Scholar]
- Lorente-Cebrian, S.; Perez-Matute, P.; Martinez, J.A.; Marti, A.; Moreno-Aliaga, M.J. Effects of eicosapentaenoic acid (EPA) on adiponectin gene expression and secretion in primary cultured rat adipocytes. J. Physiol. Biochem. 2006, 62, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Aliaga, M.J.; Lorente-Cebrian, S.; Martinez, J.A. Regulation of adipokine secretion by n-3 fatty acids. Proc. Nutr. Soc. 2010, 69, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Oliver, E.; McGillicuddy, F.; Phillips, C.; Toomey, S.; Roche, H.M. The role of inflammation and macrophage accumulation in the development of obesity-induced type 2 diabetes mellitus and the possible therapeutic effects of long-chain n-3 PUFA. Proc. Nutr. Soc. 2010, 69, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Ailhaud, G.; Guesnet, P.; Cunnane, S.C. An emerging risk factor for obesity: Does disequilibrium of polyunsaturated fatty acid metabolism contribute to excessive adipose tissue development? Br. J. Nutr. 2008, 100, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Flachs, P.; Rossmeisl, M.; Kopecky, J. The effect of n-3 fatty acids on glucose homeostasis and insulin sensitivity. Physiol. Res. 2014, 63 (Suppl. 1), S93–S118. [Google Scholar]
- Kalupahana, N.S.; Claycombe, K.J.; Moustaid-Moussa, N. (n-3) fatty acids alleviate adipose tissue inflammation and insulin resistance: Mechanistic insights. Adv. Nutr. 2011, 2, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Wielinga, P.Y.; Harthoorn, L.F.; Verschuren, L.; Schoemaker, M.H.; Jouni, Z.E.; van Tol, E.A.; Kleemann, R.; Kooistra, T. Arachidonic acid/docosahexaenoic acid-supplemented diet in early life reduces body weight gain, plasma lipids, and adiposity in later life in apoe*3leiden mice. Mol. Nutr. Food Res. 2012, 56, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Visser, S.; Slangen, C.J.; Rollema, H.S. Phenotyping of bovine milk proteins by reversed-phase high-performance liquid chromatography. J. Chromatogr. 1991, 548, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Lamers, D.; Schlich, R.; Greulich, S.; Sasson, S.; Sell, H.; Eckel, J. Oleic acid and adipokines synergize in inducing proliferation and inflammatory signalling in human vascular smooth muscle cells. J. Cell. Mol. Med. 2011, 15, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Dietze-Schroeder, D.; Sell, H.; Uhlig, M.; Koenen, M.; Eckel, J. Autocrine action of adiponectin on human fat cells prevents the release of insulin resistance-inducing factors. Diabetes 2005, 54, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Bogin, E.; Ziv, G. Enzymes and minerals in normal and mastitic milk. Cornell Vet. 1973, 63, 666–676. [Google Scholar] [PubMed]
- Kim, H.K.; Della-Fera, M.; Lin, J.; Baile, C.A. Docosahexaenoic acid inhibits adipocyte differentiation and induces apoptosis in 3T3-L1 preadipocytes. J. Nutr. 2006, 136, 2965–2969. [Google Scholar] [PubMed]
- Von Berg, A.; Filipiak-Pittroff, B.; Krämer, U.; Link, E.; Bollrath, C.; Brockow, I.; Koletzko, S.; Grübl, A.; Heinrich, J.; Wichmann, H.-E.; et al. Preventive effect of hydrolyzed infant formulas persists until age 6 years: Long-term results from the German Infant Nutritional Intervention study (GINI). Am. Acad. Allergy Asthma Immunol. 2008, 121, 1442–1447. [Google Scholar]
- Chatterton, D.E.W.; Nguyen, D.N.; Bering, S.B.; Sangild, P.T. Anti-inflammatory mechanisms of bioactive milk proteins in the intestine of newborns. Int. J. Biochem. Cell Biol. 2013, 45, 1730–1747. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhya, A.; Noronha, N.; Bahar, B.; Ryan, M.T.; Murray, B.A.; Kelly, P.M.; O’Loughlin, I.B.; O’Doherty, J.V.; Sweeney, T. Anti-inflammatory effects of a casein hydrolysate and its peptide-enriched fractions on TNFα-challenged Caco-2 cells and LPS-challenged porcine colonic explants. Food Sci. Nutr. 2014, 2, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, A.N.; Murphy, M.J.; Metcalf, B.S.; Hosking, J.; Voss, L.D.; English, P.; Sattar, N.; Wilkin, T.J. Adiponectin in childhood. Int. J. Pediatr. Obes. 2008, 3, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Flexeder, C.; Thiering, E.; Kratzsch, J.; Klumper, C.; Koletzko, B.; Muller, M.J.; Koletzko, S.; Heinrich, J. Is a child’s growth pattern early in life related to serum adipokines at the age of 10 years? Eur. J. Clin. Nutr. 2014, 68, 25–31. [Google Scholar] [CrossRef] [PubMed]
- De Zegher, F.; Sebastiani, G.; Diaz, M.; Sanchez-Infantes, D.; Lopez-Bermejo, A.; Ibanez, L. Body composition and circulating high-molecular-weight adiponectin and IGF-I in infants born small for gestational age: Breast- vs. formula-feeding. Diabetes 2012, 61, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cundiff, J.K.; Maria, S.D.; McMahon, R.J.; Wickham, M.S.; Faulks, R.M.; van Tol, E.A. Differential digestion of human milk proteins in a simulated stomach model. J. Proteome Res. 2014, 13, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, D.S.; Theil, P.K.; Larsen, L.B.; Purup, S. Effect of milk hydrolysates on inflammation markers and drug-induced transcriptional alterations in cell-based models. J. Anim. Sci. 2013, 90 (Suppl. 4), 403–405. [Google Scholar]
- Phelan, M.; Aherne-Bruce, S.A.; O’Sullivan, D.; FitzGerald, R.J.; O’Brien, N.M. Potential bioactive effects of casein hydrolysates on human cultured cells. Int. Dairy J. 2009, 19, 279–285. [Google Scholar] [CrossRef]
- Phelan, M.; Aisling Aherne, S.; O’Sullivan, D.; FitzGerald, R.J.; O’Brien, N.M. Growth inhibitory effects of casein hydrolysates on human cancer cell lines. J. Dairy Res. 2010, 77, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Mandujano, G.; Weiss-Steider, B.; Melo, B.; Cordova, Y.; Ledesma-Martinez, E.; Bustos, S.; Silvestre, O.; Aguiniga, I.; Sosa, N.; Martinez, I.; et al. Alpha-, beta- and kappa-caseins inhibit the proliferation of the myeloid cell lines 32d cl3 and wehi-3 and exhibit different differentiation properties. Immunobiology 2008, 213, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Visser, J.T.; Bos, N.A.; Harthoorn, L.F.; Stellaard, F.; Beijer-Liefers, S.; Rozing, J.; van Tol, E.A. Potential mechanisms explaining why hydrolyzed casein-based diets outclass single amino acid-based diets in the prevention of autoimmune diabetes in diabetes-prone BB rats. Diabetes Metab. Res. Rev. 2012, 28, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Emani, R.; Asghar, M.N.; Toivonen, R.; Lauren, L.; Soderstrom, M.; Toivola, D.M.; van Tol, E.A.; Hanninen, A. Casein hydrolysate diet controls intestinal T cell activation, free radical production and microbial colonisation in nod mice. Diabetologia 2014, 56, 1781–1791. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Dipeptidyl peptidase IV inhibitory and antioxidative properties of milk protein-derived dipeptides and hydrolysates. Peptides 2012, 39, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Perez-Matute, P.; Marti, A.; Martinez, J.A.; Fernandez-Otero, M.P.; Stanhope, K.L.; Havel, P.J.; Moreno-Aliaga, M.J. Eicosapentaenoic fatty acid increases leptin secretion from primary cultured rat adipocytes: Role of glucose metabolism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1682–R1688. [Google Scholar] [CrossRef] [PubMed]
- Oster, R.T.; Tishinsky, J.M.; Yuan, Z.; Robinson, L.E. Docosahexaenoic acid increases cellular adiponectin mRNA and secreted adiponectin protein, as well as PPARgamma mRNA, in 3T3-L1 adipocytes. Appl. Physiol. Nutr. Metab. 2010, 35, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Tishinsky, J.M.; Ma, D.W.; Robinson, L.E. Eicosapentaenoic acid and rosiglitazone increase adiponectin in an additive and PPARgamma-dependent manner in human adipocytes. Obesity (Silver Spring) 2011, 19, 262–268. [Google Scholar] [CrossRef]
- Banga, A.; Unal, R.; Tripathi, P.; Pokrovskaya, I.; Owens, R.J.; Kern, P.A.; Ranganathan, G. Adiponectin translation is increased by the PPARgamma agonists pioglitazone and omega-3 fatty acids. Am. J. Physiol. Endocrinol. Metab. 2009, 296, 480–489. [Google Scholar] [CrossRef]
- Rossi, A.S.; Lombardo, Y.B.; Lacorte, J.M.; Chicco, A.G.; Rouault, C.; Slama, G.; Rizkalla, S.W. Dietary fish oil positively regulates plasma leptin and adiponectin levels in sucrose-fed, insulin-resistant rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, 486–494. [Google Scholar] [CrossRef]
- Lefils-Lacourtablaise, J.; Socorro, M.; Géloën, A.; Daira, P.; Debard, C.; Loizon, E.; Guichardant, M.; Dominguez, Z.; Vidal, H.; Lagarde, M.; Bernoud-Hubac, N. The eicosapentaenoic acid metabolite 15-deoxy-δ(12,14)-prostaglandin J3 increases adiponectin secretion by adipocytes partly via a PPARγ-dependent mechanism. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lam, K.S.L.; Yau, M.; Xu, A. Post-translational modifications of adiponectin: Mechanisms and functional implications. Biochem. J. 2008, 409, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Granado, M.; Fuente-Martin, E.; Garcia-Caceres, C.; Argente, J.; Chowen, J.A. Leptin in early life: A key factor for the development of the adult metabolic profile. Obes. Facts 2012, 5, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Kaji, H.; Takahashi, Y.; Iida, K.; Mizuno, I.; Okimura, Y.; Abe, H.; Chihara, K. Stimulation by eicosapentaenoic acids of leptin mrna expression and its secretion in mouse 3T3-L1 adipocytes in vitro. Biochem. Biophys. Res. Commun. 2000, 270, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Reseland, J.E.; Haugen, F.; Hollung, K.; Solvoll, K.; Halvorsen, B.; Brude, I.R.; Nenseter, M.S.; Christiansen, E.N.; Drevon, C.A. Reduction of leptin gene expression by dietary polyunsaturated fatty acids. J. Lipid Res. 2001, 42, 743–750. [Google Scholar] [PubMed]
- Maury, E.; Brichard, S.M. Adipokine dysregulation, adipose tissue inflammation and metabolic syndrome. Mol. Cell. Endocrinol. 2010, 314, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Kihara, S.; Funahashia, T.; Matsuzawaa, Y.; Walsh, K. Obesity, adiponectin and vascular inflammatory disease. Curr. Opin. Lipidol. 2003, 14, 561–566. [Google Scholar] [CrossRef]
- Valentine, C.J. Maternal dietary dha supplementation to improve inflammatory outcomes in the preterm infant. Adv. Nutr. 2012, 3, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, A.A.; Rayner, D.V.; Mitchell, S.E.; Bashir, S.; Ha, J.H.; Wahle, K.W.; Morris, A.C.; Williams, L.M. Dietary supplementation with conjugated linoleic acid plus n-3 polyunsaturated fatty acid increases food intake and brown adipose tissue in rats. Nutrients 2009, 1, 178–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titos, E.; Rius, B.; Gonzalez-Periz, A.; Lopez-Vicario, C.; Moran-Salvador, E.; Martinez-Clemente, M.; Arroyo, V.; Claria, J. Resolvin d1 and its precursor docosahexaenoic acid promote resolution of adipose tissue inflammation by eliciting macrophage polarization toward an m2-like phenotype. J. Immunol. 2012, 187, 5408–5418. [Google Scholar] [CrossRef]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. Gpr120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Russell, F.D.; Burgin-Maunder, C.S. Distinguishing health benefits of eicosapentaenoic and docosahexaenoic acids. Mar. Drugs 2012, 10, 2535–2559. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romacho, T.; Glosse, P.; Richter, I.; Elsen, M.; Schoemaker, M.H.; Van Tol, E.A.; Eckel, J. Nutritional Ingredients Modulate Adipokine Secretion and Inflammation in Human Primary Adipocytes. Nutrients 2015, 7, 865-886. https://doi.org/10.3390/nu7020865
Romacho T, Glosse P, Richter I, Elsen M, Schoemaker MH, Van Tol EA, Eckel J. Nutritional Ingredients Modulate Adipokine Secretion and Inflammation in Human Primary Adipocytes. Nutrients. 2015; 7(2):865-886. https://doi.org/10.3390/nu7020865
Chicago/Turabian StyleRomacho, Tania, Philipp Glosse, Isabel Richter, Manuela Elsen, Marieke H. Schoemaker, Eric A. Van Tol, and Jürgen Eckel. 2015. "Nutritional Ingredients Modulate Adipokine Secretion and Inflammation in Human Primary Adipocytes" Nutrients 7, no. 2: 865-886. https://doi.org/10.3390/nu7020865
APA StyleRomacho, T., Glosse, P., Richter, I., Elsen, M., Schoemaker, M. H., Van Tol, E. A., & Eckel, J. (2015). Nutritional Ingredients Modulate Adipokine Secretion and Inflammation in Human Primary Adipocytes. Nutrients, 7(2), 865-886. https://doi.org/10.3390/nu7020865