Can We Translate Vitamin D Immunomodulating Effect on Innate and Adaptive Immunity to Vaccine Response?

Abstract
:1. Introduction
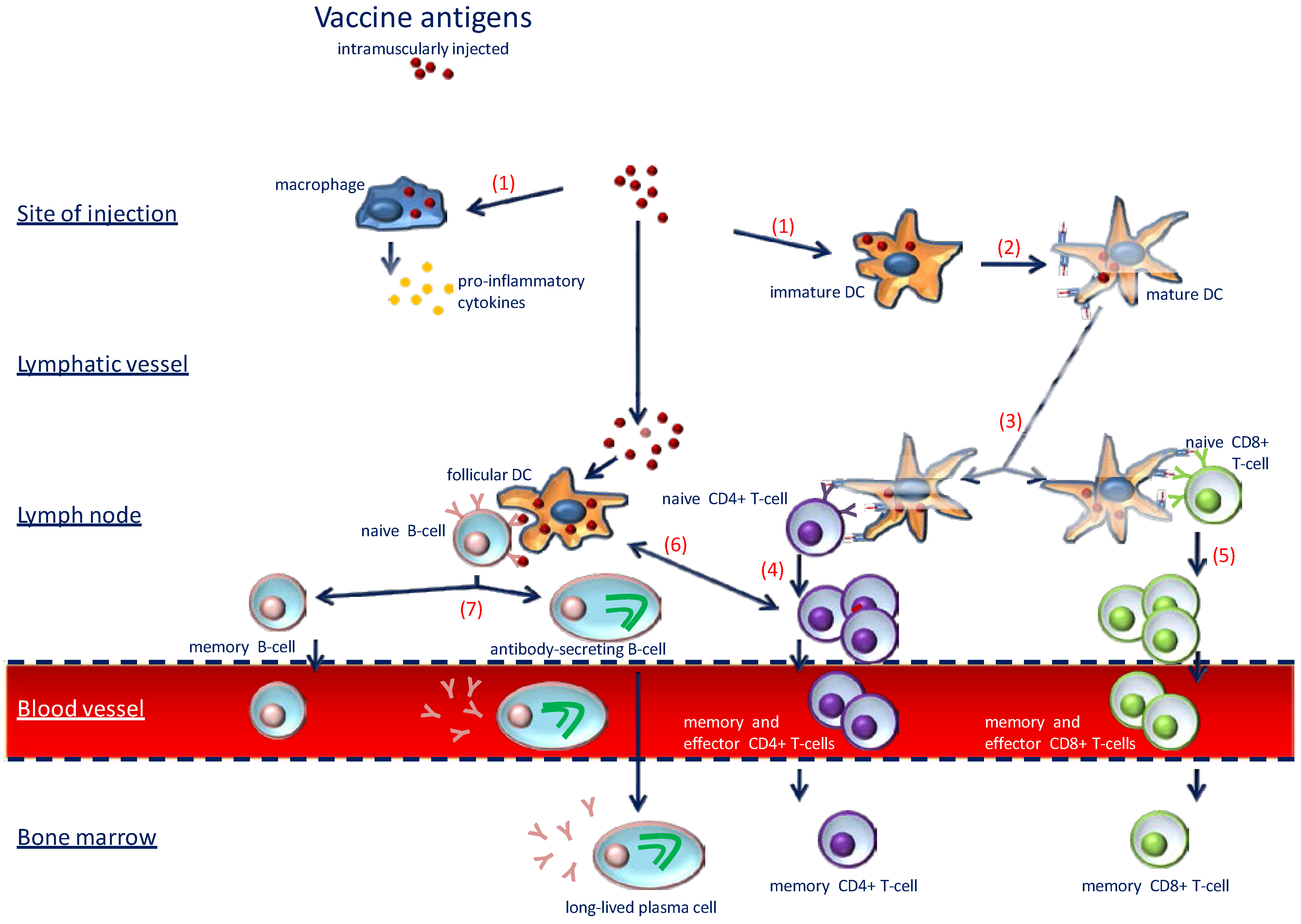
2. The Immune System: The Art of War
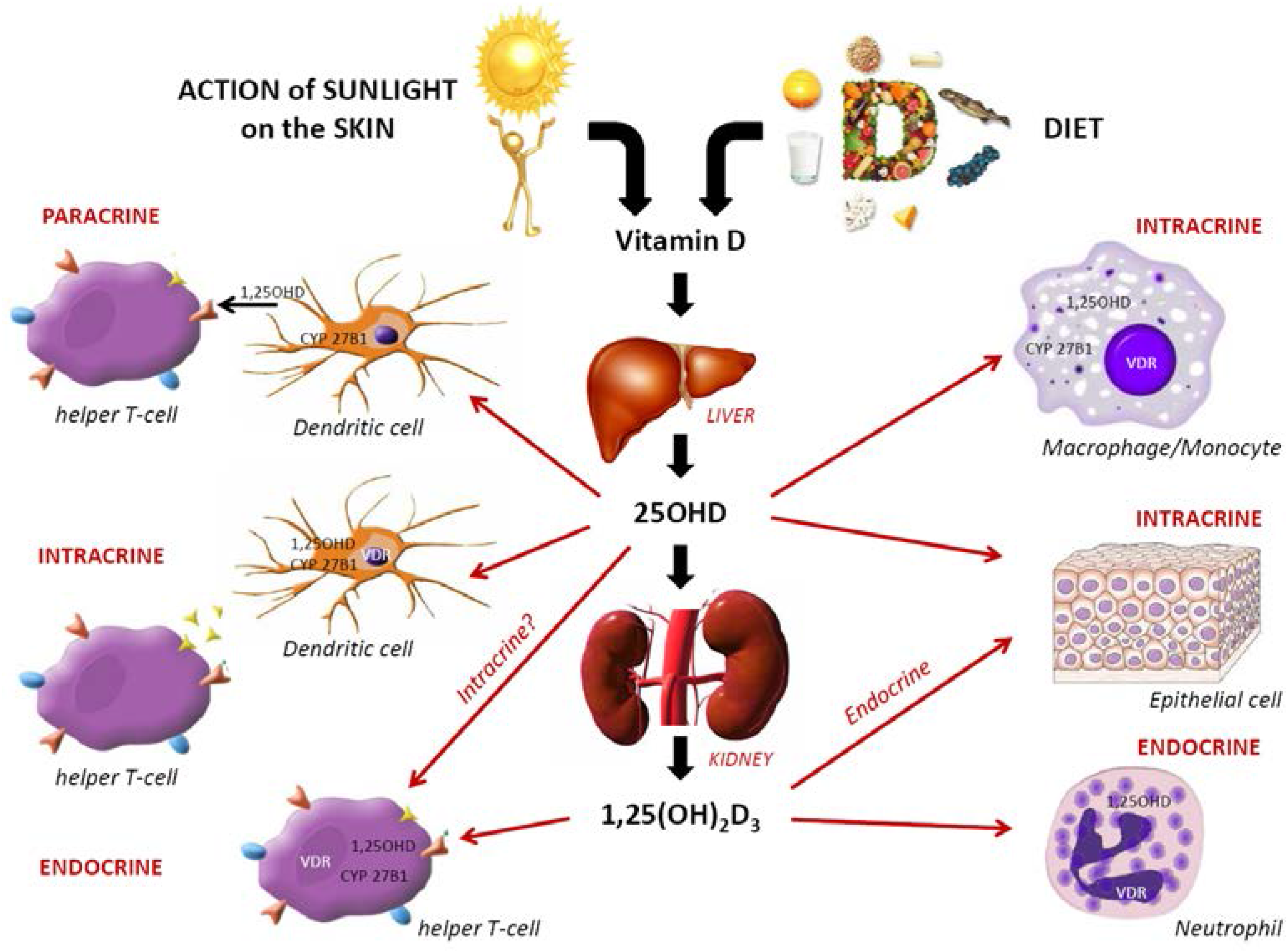
3. Immunomodulating Role of VitD in Host Protection


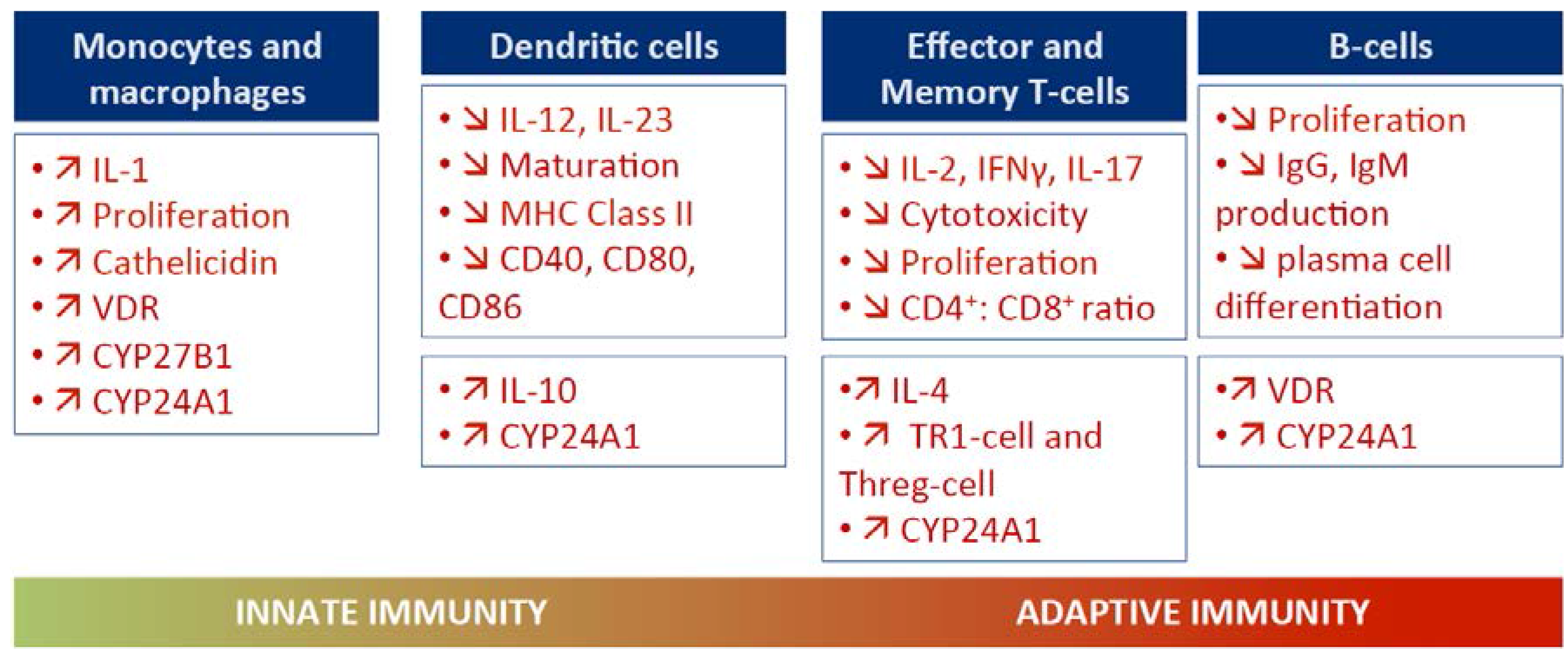
3.1. VitD and Innate Immunity
3.2. VitD and Adaptive Immunity
3.3. Mechanisms by Which VitD May Induce Anti-Infection Protection
4. VitD Status and Vaccine Immunogenicity
4.1. Good Immune Response: The Key to Success of Vaccinations

4.2. Translating Immunomodulatory Roles of VitD in Vaccine Response
5. What Do We Know from Clinical Trial and Observational Studies?

{kind=link}
{kind=link}
{kind=link}
| Vaccines | Reference | Study | VitD Supplementation | VitD Status (post-Replacement) | Results | ||
|---|---|---|---|---|---|---|---|
| Design | Population | Dosage (IU) | Duration | ||||
| Influenza | Kriesel et al. [64] | RCT | 175 adults (USA) | 40 ** | One dose | NR | A |
| Hepatitis B | Moe et al. [68] | RCT | 31 haemodialysis patients (USA) | 144 μg * | 12 weeks | NR | A |
| Herpes Zoster | Ginde et al. [69] | SGA | 150 nursing home residents (60 years old or over—USA) | High dose vs. normal dose | 4 months | ? | ? |
| VDR SNPs/Haplotype | |||||||
| Measles | Ovsyannikova et al. [61] | SGA | 745 healthy children genotyped for the 391 polymorphisms in their VDR (11–22 years old—USA) | Association between multiple VDR SNPs/haplotype and vaccine adaptive immune response | NR | C | |
| Rubella | Ovsyannikova et al. [62] | SGA | 714 healthy children genotyped for the 148 candidate SNPs (11–19 years old—USA) | Association between polymorphism in VDR and vaccine adaptive immune response | NR | C | |
| Yearly Season of Inoculation | |||||||
| Rubella | Linder et al. [70] | SGA | Children aged 4–5 years (Israel) divided in 3 groups according to the season of the year in which vaccination was performed | The association between vaccine immunogenicity and with sunlight variability-related to yearly season | NR | C | |
6. Conclusions
Author Contributions
Conflicts of Interest
References
- Holick, M.F. Sunlight and vitamin D for bone health and prevention of autoimmune disease, cancers, and cardiovascular disease. Am. J. Clin. Nutr. 2004, 80, 1678S–1688S. [Google Scholar] [PubMed]
- Rosen, C.J. Vitamin D insufficiency. N Engl. J. Med. 2011, 364, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Wolpowitz, D.; Gilchrest, B.A. The vitamin D questions: How much do you need and how should you get it? J. Am. Acad. Dermatol. 2006, 54, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.O.; Samaras, D.; Samaras, N. Does vitamin D deficiency contribute to further impige the state of vulnerability to infections of aging and aged adults? Eur. Geriatr. Med. 2013, 4, 59–65. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, J.; DeLuca, H.F. Where is the vitamin D receptor? Arch. Biochem. Biophys. 2012, 523, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A.; Repa, J.; Evans, R.M.; Mangelsdorf, D.J. Nuclear receptors and lipid physiology: Opening hte X-files. Science 2001, 294, 1866–1870. [Google Scholar] [CrossRef]
- Lang, P.O.; Samaras, D.; Samaras, N.; Aspinall, R. How important is vitamin D in preventing infections? Osteoporos. Int. 2013, 24, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- Dawson-Hughes, B.; Mithal, A.; Bonjour, J.P.; Boonen, S.; Burckhardt, P.; Fuleihan, G.E.; Josse, R.G.; Lips, P.; Morales-Torres, J.; Yoshimura, N. IOF position statement: Vitamin D recommendations for older adults. Osteoporos. Int. 2010, 21, 1151–1154. [Google Scholar] [CrossRef] [PubMed]
- Biesalski, H.K. Vitamin D recommendations—Beyond deficiency. Ann. Nutr. Metab. 2011, 59, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Khoo, L.A.; Chai, L.; Koenen, H.; Joosten, I.; Netea, M.; van der Ven, A. Translating the role of vitamin D3 in infectious diseases. Crit. Rev. Microbiol. 2012, 38, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Battersby, A.J.; Kampmann, B.; Burl, S. Vitamin D in early childhood and the effect on immunity to mycobacterium tuberculosis. Clin. Dev. Immunol. 2012, 2012, 430972. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D: Extraskeletal health. Rheum. Dis. Clin. North. Am. 2012, 38, 141–160. [Google Scholar] [CrossRef] [PubMed]
- Prietl, B.; Treiber, G.; Pieber, T.R.; Amrein, K. Vitamin D and immune function. Nutrients 2013, 5, 2502–2521. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, M.; Malaguarnera, M.; Nicoletti, F.; Malaguarnera, L. Vitamin D3: A helpful immuno-modulator. Immunology 2011, 134, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.O.; Samaras, D. Aging adults and seasonal influenza: Does the vitamin D status (h) arm the body? J. Aging Res. 2012, 2012, 806198. [Google Scholar] [CrossRef] [PubMed]
- Bischoff-Ferrari, H.A.; Dawson-Hughes, B.; Platz, A.; Orav, E.J.; Stähelin, H.B.; Willett, W.C.; Can, U.; Egli, A.; Mueller, N.J.; Looser, S.; et al. Effect of high-dosage cholecalciferol and extended physiotherapy on complications after hip fracture: A randomized controlled trial. Arch. Intern. Med. 2010, 170, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.O.; Govind, S.; Aspinall, R. L’immunosénescence. Neurol. Psychiatr. Geriatr. 2012, 12, 171–181. [Google Scholar]
- Lang, P.O.; Aspinall, R. Vaccination in the elderly: What can be recommended. Drugs Aging 2014, 31, 581–599. [Google Scholar] [CrossRef] [PubMed]
- White, J.H. Vitamin D metabolism and signaling in the immune system. Rev. Endocr. Metab. Disord. 2012, 13, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Hewison, M. An update on vitamin D and human immunity. Clin. Endocrinol. 2012, 76, 315–325. [Google Scholar] [CrossRef]
- Liu, P.T.; Stenger, S.; Li, H.; Wenzel, L.; Tan, B.H.; Krutzik, S.R.; Ochoa, M.T.; Schauber, J.; Wu, K.; Meinken, C.; et al. Toll-like receptor triggering of a vitamin D—mediated human antimicrobial response. Science 2006, 311, 1770–1773. [Google Scholar] [CrossRef] [PubMed]
- Van Belle, T.L.; Gysemans, C.; Mathieu, C. Vitamin D in autoimmune, infectious and allergic diseases: A vital player? Best. Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, K.; Wessner, B.; Laggner, U.; Ploder, M.; Tamandl, D.; Friedl, J.; Zügel, U.; Steinmeyer, A.; Pollak, A.; Roth, E.; et al. 1,25(OH)2D3 down-regulates monocyte TLR expression and triggers hyporesponsiveness to pathogen-associated molecular patterns. Eur. J. Immunol. 2006, 36, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human cathelicidin antimicrobial peptide (CAMP) is a direct target of the vitamin D receptor and is strongly upregulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.; Heilborn, J.D.; Chamorro Jimenez, C.I.; Hammarsjo, A.; Törmä, H.; Stahle, M. Vitamin D induces the antimicrobial protein hCAP18 in human skin. J. Invest. Dermatol. 2005, 124, 1080–1082. [Google Scholar] [CrossRef] [PubMed]
- Christakos, S.; Ajibade, D.V.; Dhawan, P.; Fechner, A.J.; Mady, L.J. Vitamin D: Metabolism. Endocrinol. Metab. Clin. North. Am. 2010, 39, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Hewison, M.; Freeman, L.; Hughes, S.V.; Evans, K.N.; Bland, R.; Eliopoulos, A.G.; Kilby, M.D.; Moss, P.A.; Chakraverty, R. Differential regulation of vitamin D receptor and its ligand in human monocyte-derived dendritic cells. J. Immunol. 2003, 170, 5382–5390. [Google Scholar] [CrossRef] [PubMed]
- Van Etten, E.; Mathieu, C. Immunoregulation by 1,25-di-hydroxy1,25(OH)2D3: Basic concepts. J. Steroid Biochem. Mol. Biol. 2005, 97, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Alroy, I.; Towers, T.; Freedman, L.P. Transcriptional repression of interleukin-2 gene by 1,25(OH)2D3: Direct inhibition NFATp/AP-1 complex formation by a hormone receptor. Mol. Cell Biol. 1995, 15, 5789–5799. [Google Scholar] [PubMed]
- Mahon, B.D.; Wittke, A.; Weaver, V.; Cantorna, M.T. The targets of vitamin D depend on the differentiation and activation status of CD4-positive T-cells. J. Cell Biochem. 2003, 89, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Sun, J. Vitamin D and mucosal immune function. Curr. Opin. Gastroenterol. 2010, 26, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.S.; Henriquez, F.; Sumar, N.; Patel, S. T helper cell subsets in arthritis and the benefits of immunomodulation by 1,25(OH)2 vitamin D. Rheumatol. Int. 2012, 32, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Ooi, J.H.; Chen, J.; Cantorna, M.T. Vitamin D regulation of immune function in the gut: Why do T cells have vitamin D receptors? Mol. Aspects Med. 2012, 33, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Peck, A.; Mellins, E.D. Precarious balance: Th17 cells in host defense. Infect. Immun. 2010, 78, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Rhee, K.J.; Albesiano, E.; Yen, H.R.; Huso, D.L.; Brancati, F.L.; Wick, E.; McAllister, F.; Housseau, F.; Pardoll, D.M.; et al. A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses. Nat. Med. 2009, 15, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Oukka, M.; Kuchroo, V.; Betteli, E. Th17 cells: Effector cells with inflammatory properties. Semin. Immunol. 2007, 19, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Sims, G.P.; Chen, X.X.; Gu, Y.Y.; Chen, S.; Lipsky, P.E. Modulatory effects of 1,25-Dihydroxyvitamin D3 on human B cell differentiation. J. Immunol. 2007, 179, 1634–1647. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.S.; Gacad, M.A. Characterization of 1 alpha-hydroxylation of 1,25(OH)2D3 sterol by cultured alveolar macrophages from patients with sarcoidosis. J. Exp. Med. 1985, 161, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Vernino, L.; McAnally, L.M.; Ramberg, J.; Lipsky, P.E. Generation of nondividing high-rate Ig-secreting plasma-cells in cultures of human B-cells stimulated with anti-Cd3-acti-vated T-cells. J. Immunol. 1992, 148, 404–410. [Google Scholar] [PubMed]
- Heine, G.; Niesner, U.; Chang, H.D.; Steinmeyer, A.; Zügel, U.; Zuberbier, T.; Radbruch, A.; Worm, M. 1,25-dihydroxyvitamin D(3) promotes IL-10 production in human B cells. Eur. J. Immunol. 2008, 38, 2210–2218. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, A.K.; Nagakubo, D.; Hieshima, K.; Nakayama, T.; Jin, Z.; Yoshie, O. 1,25-dihydroxyvitamin D3 induces CCR10 expression in terminally differentiating human B cells. J. Immunol. 2008, 180, 2786–2795. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, L.E.; Burke, F.; Mura, M.; Zheng, Y.; Qureshi, O.S.; Hewison, M.; Walker, L.S.; Lammas, D.A.; Raza, K.; Sansom, D.M. 1,25-dihydroxyvitamin D(3) and IL-2 combine to inhibit T cell production of inflammatory cytokines and promote development of regulatory T cells expressing CTLA-4 and FoxP3. J. Immunol. 2009, 183, 5458–5467. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Nestel, F.P.; Bourdeau, V.; Nagai, Y.; Wang, Q.; Liao, J.; Tavera-Mendoza, L.; Lin, R.; Hanrahan, J.W.; Mader, S.; et al. Cutting edge: 1,25-dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J. Immunol. 2004, 173, 2909–2912. [Google Scholar] [CrossRef] [PubMed]
- Montoya, D.; Cruz, D.; Teles, R.M.; Lee, D.J.; Ochoa, M.T.; Krutzik, S.R.; Chun, R.; Schenk, M.; Zhang, X.; Ferguson, B.G.; et al. Divergence of macrophage phagocytic and antimicrobial programs in leprosy. Cell Host Microbe 2009, 6, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Bergman, P.; Walter-Jallow, L.; Broliden, K.; Agerberth, B.; Söderlund, J. The antimicrobial peptide LL-37 inhibits HIV-1 replication. Curr. HIV Res. 2007, 5, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Gunville, C.F.; Mourani, P.M.; Ginde, A.A. The role of vitamin D in prevention and treatment of infection. Inflamm. Allergy Drug Targets 2013, 11, 239–245. [Google Scholar] [CrossRef]
- Lang, P.O.; Govind, S.; Bokum, A.T.; Kenny, N.; Matas, E.; Pitts, D.; Aspinall, R. Immune senescence and vaccination in the elderly. Curr. Top. Med. Chem. 2013, 13, 2541–2550. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, C.A.; Aspinall, R. B-cell responses to vaccination at the extreme of age. Nat. Rev. Immunol. 2009, 9, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Aspinall, R.; Lang, P.O. Vaccine responsiveness in the elderly: Best practice for the clinic. Expert Rev. Vaccines 2014, 13, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Penna, G.; Adorini, L. 1 alpha,25-dihydroxyvitamin D-3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J. Immunol. 2000, 164, 2405–2411. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, D.; Cippitelli, M.; Cocciolo, G.; Mazzeo, D.; di Lucia, P.; Lang, R.; Sinigaglia, F.; Panina-Bordignon, P. Inhibition of IL-12 production by 1,25-dihydroxyvitamin D-3: Involvement of NF-κB downregulation in transcriptional repression of the p40 gene. J. Clin Invest. 1998, 101, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, A.; Barrat, F.J.; Crain, C.; Heath, V.L.; Savelkoul, H.F.J.; O’Garra, A. 1 alpha,25-dihydroxyvitamin D3 has a direct effect on naive CD4+ T cells to enhance the development of Th2 cells. J. Immunol. 2001, 167, 4974–4980. [Google Scholar] [CrossRef]
- Cippitelli, M.; Fionda, C.; di Bona, D.; di Rosa, F.; Lupo, A.; Piccoli, M.; Frati, L.; Santoni, A. Negative regulation of CD95 ligand gene expression by vitamin D3 in T lymphocytes. J. Immunol. 2002, 168, 1154–1166. [Google Scholar] [CrossRef] [PubMed]
- Bruce, D.; Ooi, J.H.; Yu, S.; Cantorna, M.T. Vitamin D and host resistance to infection? Putting the cart in front of the horse. Exp. Biol. Med. (Maywood) 2010, 235, 921–927. [Google Scholar] [CrossRef]
- Hewison, M. Vitamin D and the immune system: New perspectives on an old theme. Endocrinol. Met. Clin. N. Am. 2010, 39, 365–379. [Google Scholar] [CrossRef]
- Enioutina, E.Y.; Bareyan, D.; Daynes, R.A. TLR ligands that stimulate the metabolism of vitamin D3 in activated murine dendritic cells can function as effective mucosal adjuvants to subcutaneously administered vaccines. Vaccine 2008, 26, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Enioutina, E.Y.; Bareyan, D.; Daynes, R.A. TLR-induced local metabolism of vitamin D3 plays an important role in the diversification of adaptive immune responses. J. Immunol. 2009, 182, 4296–4305. [Google Scholar] [CrossRef] [PubMed]
- Daynes, R.A.; Araneo, B.A.; Hennebold, J.; Enioutina, E.; Mu, H.H. Steroids as regulators of the mammalian immune response. J. Investig. Dermatol. 1995, 105, 14S–19S. [Google Scholar] [CrossRef] [PubMed]
- Daynes, R.A.; Araneo, B.A. The development of effective vaccine adjuvants employing natural regulators of T-cell lymphokine production in vivo. Ann. N. Y. Acad. Sci. 1994, 730, 144–161. [Google Scholar] [CrossRef] [PubMed]
- Zitt, E.; Sprenger-Mähr, H.; Knoll, F.; Neyer, U.; Lhotta, K. Vitamin D deficiency is associated with poor response to active hepatitis B immunisation in patients with chronic kidney disease. Vaccine 2012, 30, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Ovsyannikova, I.G.; Haralambieva, I.H.; Vierkant, R.A.; OʼByrne, M.M.; Jacobson, R.M.; Poland, G.A. Effects of vitamin A and D receptor gene polymorphisms/haplotypes on immune responses to measles vaccine. Pharm. Genomics 2012, 22, 20–31. [Google Scholar] [CrossRef]
- Ovsyannikova, I.G.; Dhiman, N.; Haralambieva, I.H.; Vierkant, R.A.; OʼByrne, M.M.; Jacobson, R.M.; Poland, G.A. Rubella vaccine-induced cellular immunity: Evidence of associations with polymorphisms in the Toll-like, vitamin A and D receptors, and innate immune response genes. Hum. Genet. 2010, 127, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Lalor, M.K.; Floyd, S.; Gorak-Stolinska, P.; Weir, R.E.; Blitz, R.; Branson, K.; Fine, P.E.; Dockrell, H.M. BCG vaccination: A role for vitamin D? PLoS One 2011, 6, e16709. [Google Scholar] [CrossRef] [PubMed]
- Kriesel, J.D.; Spruance, J. Calcitriol (1,25-dihydroxy-vitamin D3) coadministered with influenza vaccine does not enhance humoral immunity in human volunteers. Vaccine 1999, 17, 1883–1888. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.O.; Govind, S.; Mitchell, W.A.; Siegrist, C.A.; Aspinall, R. Vaccine effectiveness in older individuals: What has been learned from the influenza-vaccine experience. Ageing Res. Rev. 2011, 10, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bischoff-Ferrari, H.A.; Giovannucci, E.; Willett, W.C.; Dietrich, T.; Dawson-Hughes, B. Estimation of optimal serum concentrations of 25-hydroxyvitamin D for multiple health outcomes. Am. J. Clin. Nutr. 2006, 84, 18–28. [Google Scholar] [PubMed]
- Cooper, C.; Thorne, A. Canadian Hiv Trials Network Ctn Influenza Vaccine Research Group. Vitamin D supplementation does not increase immunogenicity of seasonal influenza vaccine in HIV-infected adults. HIV Clin. Trials 2011, 12, 275–276. [Google Scholar] [CrossRef] [PubMed]
- Moe, S.M.; Zekonis, M.; Harezlak, J.; Ambrosius, W.T.; Gassensmith, C.M.; Murphy, C.L.; Russell, R.R.; Batiuk, T.D. A placebo-controlled trial to evaluate immunomodulatory effects of paricalcitol. Am. J. Kidney Dis. 2001, 38, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Ginde, A.A. National Institute on Aging (NIA), Mucosal and Vaccine Research Colorado, Merck Sharp & Dohme Corp. Vitamin D Supplementation and Varicella Zoster Virus Vaccine Responsiveness in Older Long-Term Care Residents. http://clinicaltrials.gov/ct2/show/NCT01262300 (accessed on 16 March 2015).
- Linder, N.; Abudi, Y.; Abdalla, W.; Badir, M.; Amitai, Y.; Samuels, J.; Mendelson, E.; Levy, I. Effect of season of inoculation on immune response to rubella vaccine in children. J. Trop. Pediatr. 2011, 57, 299–302. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, P.O.; Aspinall, R. Can We Translate Vitamin D Immunomodulating Effect on Innate and Adaptive Immunity to Vaccine Response? Nutrients 2015, 7, 2044-2060. https://doi.org/10.3390/nu7032044
Lang PO, Aspinall R. Can We Translate Vitamin D Immunomodulating Effect on Innate and Adaptive Immunity to Vaccine Response? Nutrients. 2015; 7(3):2044-2060. https://doi.org/10.3390/nu7032044
Chicago/Turabian StyleLang, Pierre Olivier, and Richard Aspinall. 2015. "Can We Translate Vitamin D Immunomodulating Effect on Innate and Adaptive Immunity to Vaccine Response?" Nutrients 7, no. 3: 2044-2060. https://doi.org/10.3390/nu7032044
APA StyleLang, P. O., & Aspinall, R. (2015). Can We Translate Vitamin D Immunomodulating Effect on Innate and Adaptive Immunity to Vaccine Response? Nutrients, 7(3), 2044-2060. https://doi.org/10.3390/nu7032044