Increased PUFA Content and 5-Lipoxygenase Pathway Expression Are Associated with Subcutaneous Adipose Tissue Inflammation in Obese Women with Type 2 Diabetes


Abstract
:1. Introduction
2. Experimental Section
2.1. Subjects
2.2. Medication
2.3. Analysis of Number of Crown-Like Structures and Adipocyte Size
2.4. Fatty Acid Composition of WAT by GC-MS
2.5. Oxylipin Measurements in WAT by LC-MS/MS
2.6. Statistics
3. Results
3.1. Characteristics of Participants

{kind=link}
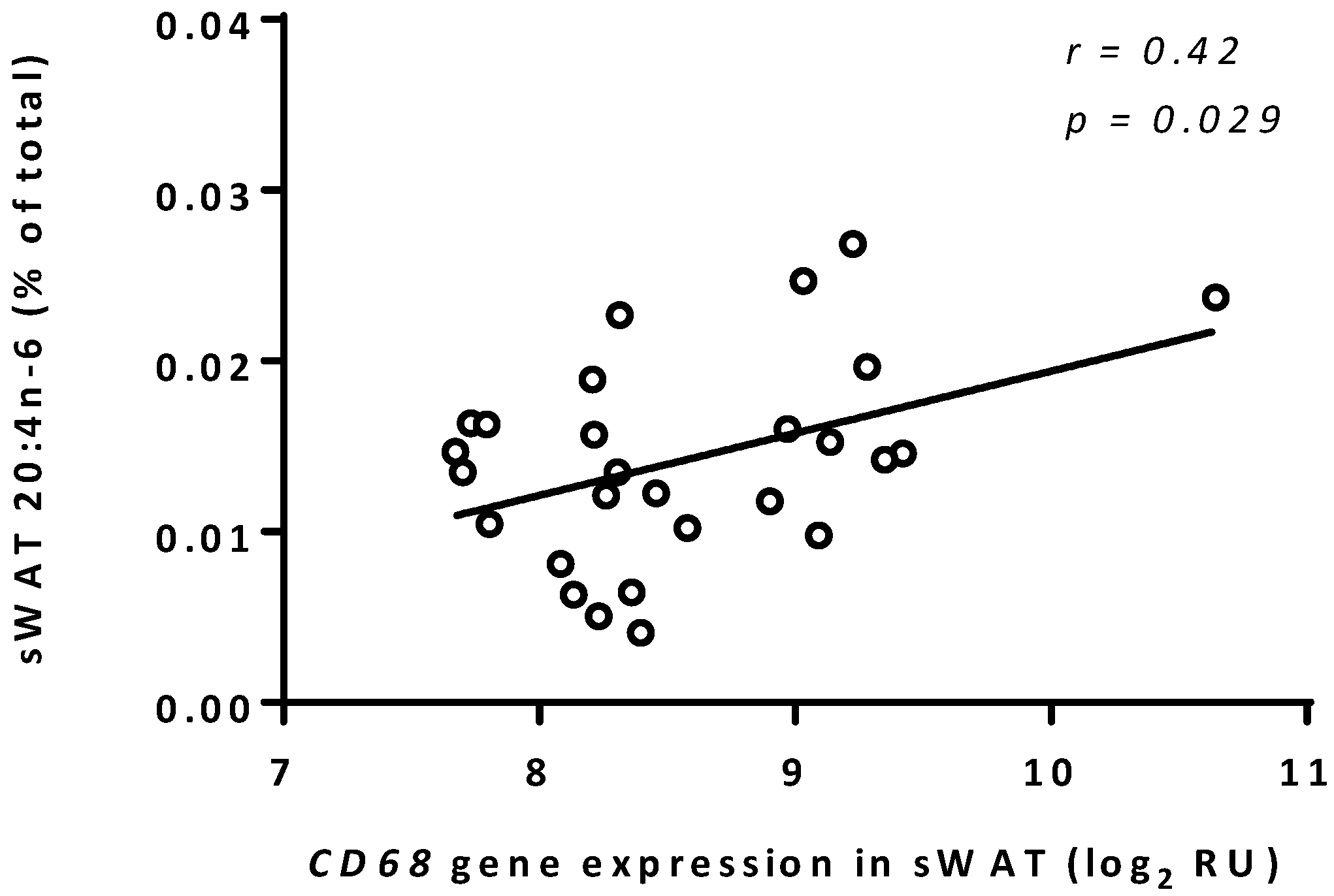{kind=link}
{kind=link}
{kind=link}
| NGT | T2DM | p-Value t-Test | |
|---|---|---|---|
| N | 19 | 16 | |
| BMI (kg/m2) | 43.4 ± 3.3 | 43.4 ± 4.5 | NS |
| Age (years) | 47 ± 7 | 52 ± 6 | NS |
| Waist circumference (cm) | 121 ± 8 | 127 ± 12 | NS |
| HOMA-IR | 2.7 ± 2.2 | 4.0 ± 3.0 | 0.08 |
| Fasting glucose (mmol/L) | 5.0 ± 0.6 | 9.0 ± 2.6 | <0.01 |
| Fasting insulin (mU/L) | 10.9 ± 7.7 | 13.3 ± 7.3 | NS |
| Total cholesterol (mmol/L) | 4.7 ± 1.1 | 4.2 ± 0.8 | NS |
| HDL cholesterol (mmol/L) | 1.1 ± 0.3 | 1.1 ± 0.3 | NS |
| LDL cholesterol (mmol/L) | 3.0 ± 1.0 | 2.2 ± 0.6 | 0.03 |
| Triglycerides (mmol/L) | 1.5 ± 0.7 | 2.0 ± 0.7 | 0.08 |
| CRP (mg/L) | 7.8 ± 7.5 | 8.2 ± 6.3 | NS |
| CD68 gene expression in sWAT | 8.2 ± 0.4 | 8.9 ± 0.7 | 0.003 |
| CD68 gene expression in vWAT | 8.2 ± 0.4 | 8.4 ± 0.5 | NS |
| # of CLS in sWAT (no/AT section) | 2.3 ± 1.4 | 12.3 ± 7.1 | 0.05 |
| # of CLS in vWAT (no/AT section) | 1.9 ± 1.2 | 2.7 ± 1.0 | NS |
| Adipocyte size in sWAT (µm) | 122 ± 16 | 126 ± 15 | NS |
| Adipocyte size in vWAT (µm) | 112 ± 16 | 118 ± 8 | NS |
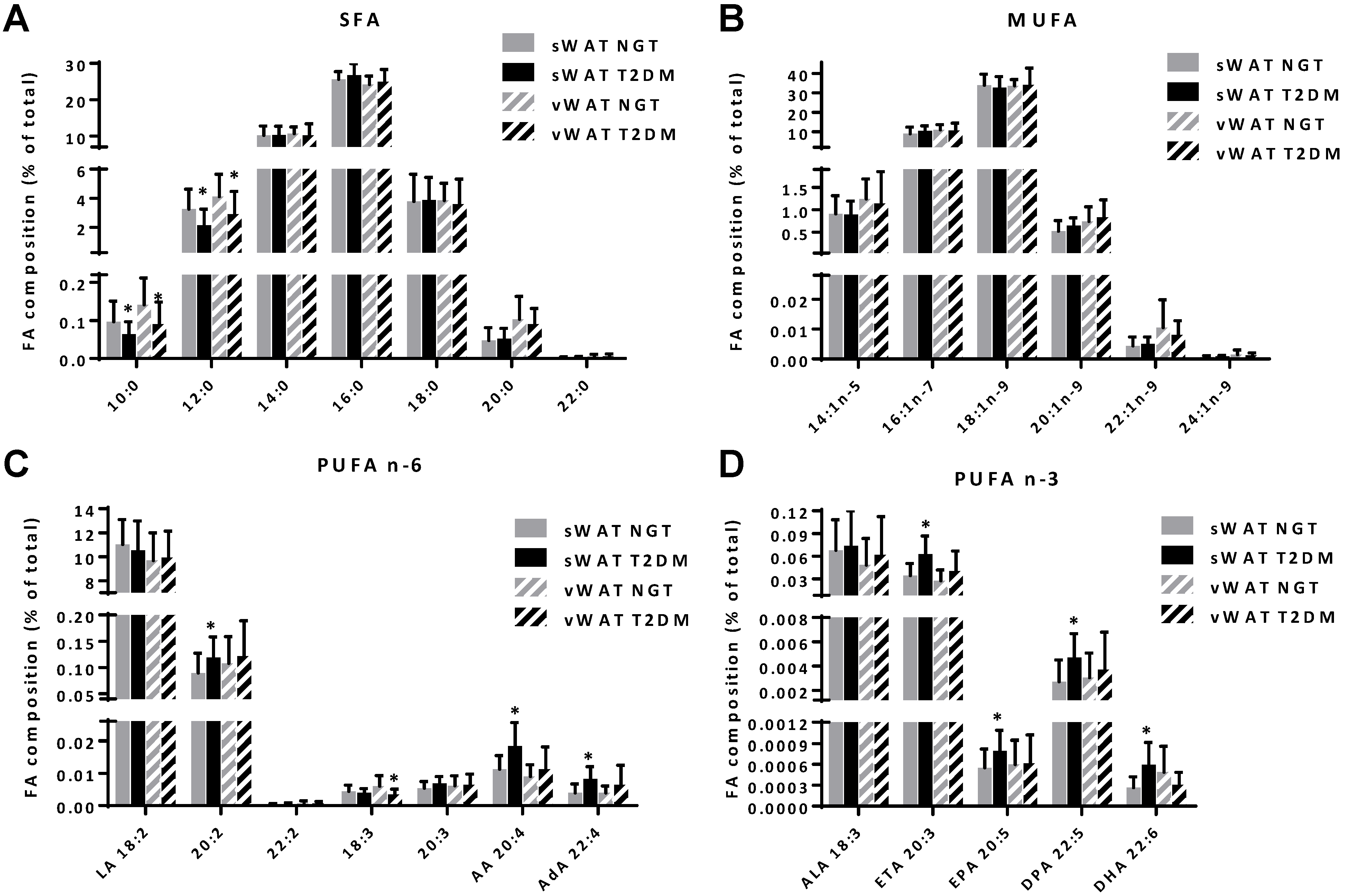
3.2. Differences in Adipose Tissue Content of Saturated Fatty Acids and PUFAs between NGT and T2DM Obese Women

3.3. Arachidonic Acid Correlated to CD68 Expression in sWAT

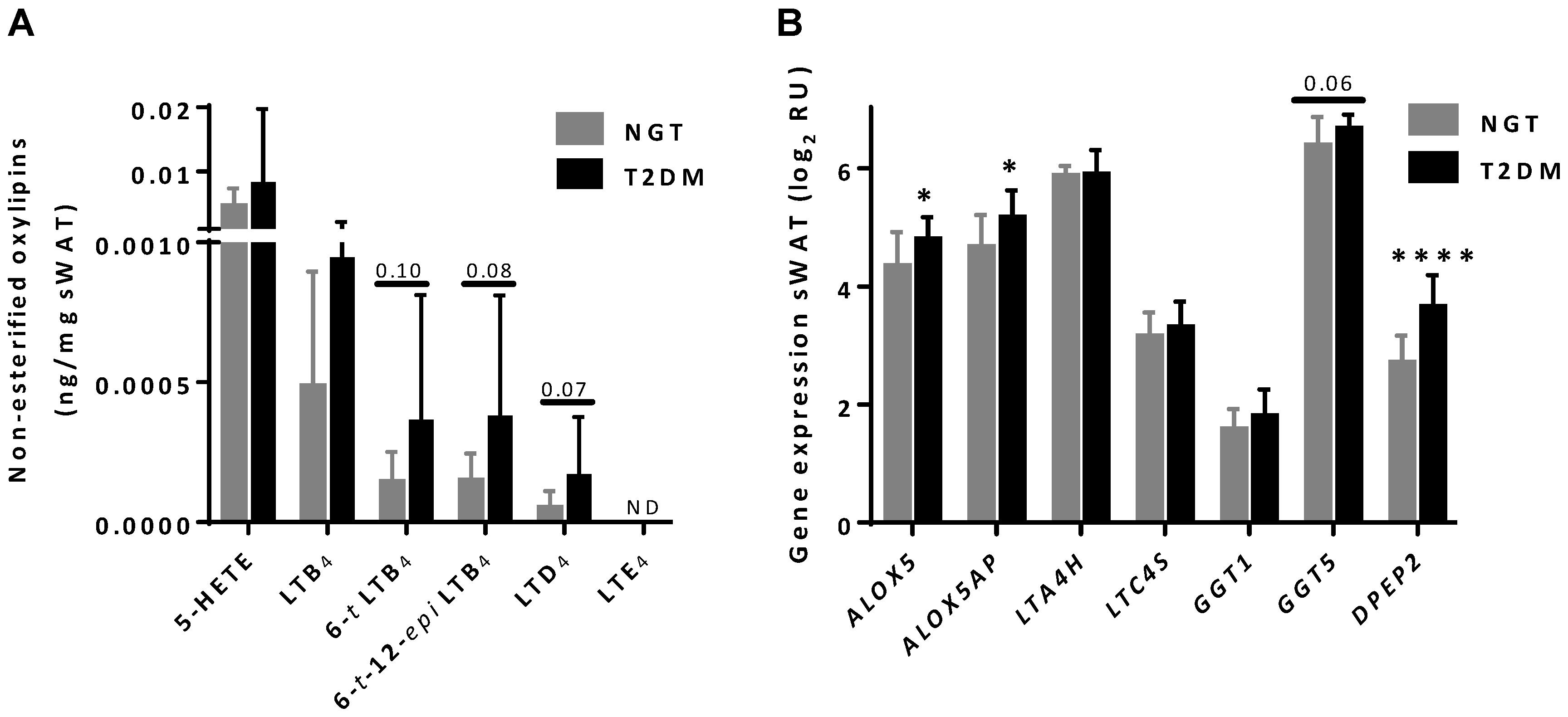
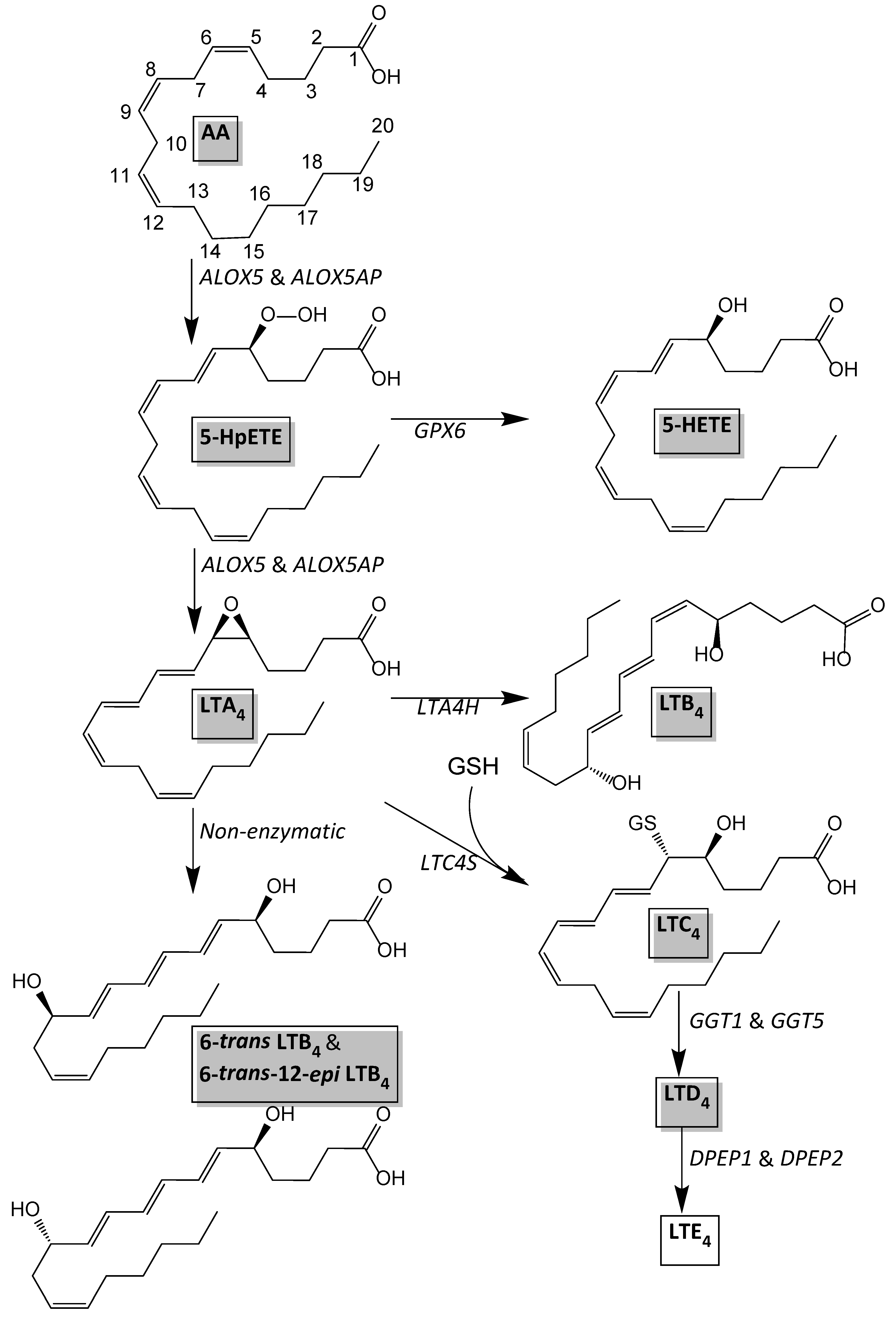
3.4. Oxylipin Levels in sWAT of Women with NGT and T2DM
3.5. Differential Expression of ALOX5, ALOX5AP and DPEP2 in sWAT between NGT and T2DM Women
| Oxylipin (pg/mg sWAT) | NGT | T2DM | NGT vs. T2DM | ||
|---|---|---|---|---|---|
| Range | Median | Range | Median | p-Value | |
| 5-HETE | 0.8–32 | 3.7 | 1.5–57 | 5.0 | NS |
| 8-HETE | 0.5–18 | 2.3 | 0.6–22 | 2.9 | NS |
| 11-HETE | 0.4–15 | 1.6 | 0.4–18 | 1.9 | NS |
| 12-HETE | 1.6–84 | 11 | 2.8–79 | 12 | NS |
| 15-HETE | 0.7–32 | 3.5 | 0.8–46 | 4.0 | NS |
| 15-HEPE | 0.09–3.1 | 0.4 | 0.2–2.6 | 0.6 | NS |
| 18-HEPE | 0.1–4.9 | 0.6 | 0.2–3.9 | 0.5 | NS |
| 7-HDHA | * | * | NS | ||
| 10-HDHA | * | * | NS | ||
| 17-HDHA | 0–3.2 | 0.5 | 0–3.2 | 1.1 | NS |
| LTB4 | 0.06–2.7 | 0.4 | 0.05–4.2 | 0.4 | NS |
| 6-trans-12-epi LTB4 | 0.03–0.3 | 0.1 | 0.06–1.7 | 0.2 | 0.08 |
| 6-trans LTB4 | 0.04–0.4 | 0.1 | 0.06–1.9 | 0.2 | 0.10 |
| 20-OH LTB4 | ND | ND | |||
| LTD4 | 0–0.2 | 0.04 | 0–0.8 | 0.08 | 0.06 |
| LTE4 | ND | ND | |||
| PGD2 | 0.06–1.1 | 0.3 | 0.04–2.7 | 0.3 | NS |
| PGE2 | 0.07–2.1 | 0.6 | 0.08–3.6 | 0.5 | NS |
| PGF2α | 0–2.1 | 0.7 | 0.1–2.5 | 0.5 | NS |
| TxB2 | 0–12 | 1.5 | 0.05–20 | 1.6 | NS |
| LXA4 | 0–0.2 | 0 | 0–1.5 | 0.02 | NS |
| LXB4 | ND | ND | |||
| AT LXA4 | ND | ND | |||
| 8-iso PGE2 | ND | ND | |||
| 8-iso PGF2α | 0–0.2 | 0.05 | 0–0.8 | 0.04 | NS |
| 15-keto PGE2 | 0.02–0.4 | 0.09 | 0.006–1.2 | 0.06 | NS |
| 13,14-dihydro-15-keto PGF2α | 0–5.0 | 0 | 0–4.6 | 0 | NS |
| RvD1 | ND | ND | |||
| RvD2 | ND | ND | |||
| AT RvD1 | ND | ND | |||
| RvE1 | ND | ND | |||
| RvE2 | # | # | |||
| 18S-RvE3 | ND | ND | |||
| 18R-RvE3 | ND | ND | |||
| 7,17-DiHDPA | 0–1.7 | 0.08 | 0–0.8 | 0 | NS |
| 19,20-DiHDPA | 0–0.3 | 0.09 | 0.06–0.4 | 0.1 | NS |
| 10S,17S-diHDHA (PDX) | 0–0.2 | 0.04 | 0–0.2 | 0.04 | NS |
| MaR1 | # | # | |||
| 7S-MaR1 | ND | ND | |||
| 5,15-diHETE | 0–20 | 2.2 | 0–87 | 3.4 | NS |
| 14,15-diHETE | 0–0.6 | 0.2 | 0–2.2 | 0.2 | NS |
| 8S,15S-diHETE | 0.04–1.3 | 0.1 | 0–1.6 | 0.2 | NS |
| 9-HoDE | 75–924 | 317 | 127–832 | 259 | NS |
| 13-HoDE | 47–527 | 161 | 65–471 | 142 | NS |
| 9-HoTrE | 3.4–49 | 21 | 3.9–37 | 15 | NS |
| 13-HoTrE | 45–502 | 285 | 57–396 | 195 | NS |
| 8(9)EET | * | * | NS | ||
| 11(12)EET | * | * | NS | ||
| 14(15)EET | * | * | NS | ||

4. Discussion

5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hotamisligil, G.S. Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef] [PubMed]
- Bluher, M. The distinction of metabolically “healthy” from “unhealthy” obese individuals. Curr. Opin. Lipidol. 2010, 21, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Rey-López, J.P.; de Rezende, L.F.; Pastor-Valero, M.; Tess, B.H. The prevalence of metabolically healthy obesity: A systematic review and critical evaluation of the definitions used. Obes. Rev. 2014, 15, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Van Beek, L.; Lips, M.A.; Visser, A.; Pijl, H.; Ioan-Facsinay, A.; Toes, R.; Berends, F.J.; Willems van Dijk, K.; Koning, F.; van Harmelen, V. Increased systemic and adipose tissue inflammation differentiates obese women with T2DM from obese women with normal glucose tolerance. Metabolism 2013, 63, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Dharuri, H.; t Hoen, P.A.; van Klinken, J.B.; Henneman, P.; Laros, J.F.; Lips, M.A.; el Bouazzaoui, F.; van Ommen, G.J.; Janssen, I.; van Ramshorst, B.; et al. Downregulation of the acetyl-CoA metabolic network in adipose tissue of obese diabetic individuals and recovery after weight loss. Diabetologia 2014, 57, 2384–2392. [Google Scholar] [CrossRef] [PubMed]
- Cinti, S.; Mitchell, G.; Barbatelli, G.; Murano, I.; Ceresi, E.; Faloia, E.; Wang, S.; Fortier, M.; Greenberg, A.S.; Obin, M.S. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J. Lipid Res. 2005, 46, 2347–2355. [Google Scholar] [CrossRef] [PubMed]
- Vachharajani, V.; Granger, D.N. Adipose tissue: A motor for the inflammation associated with obesity. IUBMB Life 2009, 61, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Masoodi, M.; Kuda, O.; Rossmeisl, M.; Flachs, P.; Kopecky, J. Lipid signaling in adipose tissue: Connecting inflammation & metabolism. Biochim. Biophys. Acta 2014, 1851, 503–518. [Google Scholar] [PubMed]
- Jacobsson, L.; Lindgarde, F.; Manthorpe, R.; Akesson, B. Correlation of fatty acid composition of adipose tissue lipids and serum phosphatidylcholine and serum concentrations of micronutrients with disease duration in rheumatoid arthritis. Ann. Rheum. Dis. 1990, 49, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Korani, M.; Firoozrai, M.; Maleki, J.; Ghahramanpour, F.; Heidari, I.; Fallah, S.; Seifi, M. Distribution of fatty acids in adipose tissue of patients with type 2 diabetes. Clin. Lab. 2012, 58, 457–464. [Google Scholar] [PubMed]
- Pezeshkian, M.; Mahtabipour, M.R. Epicardial and subcutaneous adipose tissue fatty acids profiles in diabetic and non-diabetic patients candidate for coronary artery bypass graft. BioImpacts 2013, 3, 83–89. [Google Scholar] [PubMed]
- Masi, L.N.; Rodrigues, A.C.; Curi, R. Fatty acids regulation of inflammatory and metabolic genes. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Vandanmagsar, B.; Youm, Y.H.; Ravussin, A.; Galgani, J.E.; Stadler, K.; Mynatt, R.L.; Ravussin, E.; Stephens, J.M.; Dixit, V.D. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat. Med. 2011, 17, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Tourdot, B.E.; Ahmed, I.; Holinstat, M. The emerging role of oxylipins in thrombosis and diabetes. Front. Pharmacol. 2014, 4, 176. [Google Scholar] [CrossRef] [PubMed]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Konkel, A.; Mehling, H.; Blossey, K.; Gapelyuk, A.; Wessel, N.; von Schacky, C.; Dechend, R.; Muller, D.N.; Rothe, M.; et al. Dietary omega-3 fatty acids modulate the eicosanoid profile in man primarily via the cyp-epoxygenase pathway. J. Lipid Res. 2014, 55, 1150–1164. [Google Scholar] [CrossRef] [PubMed]
- Von Moltke, J.; Trinidad, N.J.; Moayeri, M.; Kintzer, A.F.; Wang, S.B.; van Rooijen, N.; Brown, C.R.; Krantz, B.A.; Leppla, S.H.; Gronert, K. Rapid induction of inflammatory lipid mediators by the inflammasome in vivo. Nature 2012, 490, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Claria, J.; Nguyen, B.T.; Madenci, A.L.; Ozaki, C.K.; Serhan, C.N. Diversity of lipid mediators in human adipose tissue depots. Am. J. physiol. Cell physiol. 2013, 304, C1141–C1149. [Google Scholar] [CrossRef] [PubMed]
- Balvers, M.G.; Verhoeckx, K.C.; Bijlsma, S.; Rubingh, C.M.; Meijerink, J.; Wortelboer, H.M.; Witkamp, R.F. Fish oil and inflammatory status alter the n-3 to n-6 balance of the endocannabinoid and oxylipin metabolomes in mouse plasma and tissues. Metabolomics 2012, 8, 1130–1147. [Google Scholar] [CrossRef] [PubMed]
- Lips, M.A.; de Groot, G.H.; van Klinken, J.B.; Aarts, E.; Berends, F.J.; Janssen, I.M.; van Ramshorst, B.; van Wagensveld, B.A.; Swank, D.J.; van Dielen, F.; et al. Calorie restriction is a major determinant of the short-term metabolic effects of gastric bypass surgery in obese type 2 diabetic patients. Clin. Endocrinol. 2014, 80, 834–842. [Google Scholar] [CrossRef]
- Kloos, D.P.; Gay, E.; Lingeman, H.; Bracher, F.; Muller, C.; Mayboroda, O.A.; Deelder, A.M.; Niessen, W.M.; Giera, M. Comprehensive gas chromatography-electron ionisation mass spectrometric analysis of fatty acids and sterols using sequential one-pot silylation: Quantification and isotopologue analysis. Rapid Commun. Mass Spectrom. 2014, 28, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Heemskerk, M.M.; Dharuri, H.K.; van den Berg, S.A.; Jonasdottir, H.S.; Kloos, D.P.; Giera, M.; Willems van Dijk, K.; van Harmelen, V. Prolonged niacin treatment leads to increased adipose tissue poly-unsaturated fatty acid synthesis and an anti-inflammatory lipid and oxylipin plasma profile. J. Lipid Res. 2014, 55, 2532–2540. [Google Scholar] [CrossRef] [PubMed]
- Jónasdóttir, H.; Ioan-Facsinay, A.; Kwekkeboom, J.; Brouwers, H.; Zuurmond, A.-M.; Toes, R.; Deelder, A.; Giera, M. An advanced LC–MS/MS platform for the analysis of specialized pro-resolving lipid mediators. Chromatographia 2014, 78, 391–401. [Google Scholar] [CrossRef]
- Giera, M.; Ioan-Facsinay, A.; Toes, R.; Gao, F.; Dalli, J.; Deelder, A.M.; Serhan, C.N.; Mayboroda, O.A. Lipid and lipid mediator profiling of human synovial fluid in rheumatoid arthritis patients by means of LC-MS/MS. Biochim. Biophys. Acta 2012, 1821, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Periz, A.; Claria, J. Resolution of adipose tissue inflammation. ScientificWorldJournal 2010, 10, 832–856. [Google Scholar] [CrossRef] [PubMed]
- Kaaman, M.; Ryden, M.; Axelsson, T.; Nordstrom, E.; Sicard, A.; Bouloumie, A.; Langin, D.; Arner, P.; Dahlman, I. ALOX5AP expression, but not gene haplotypes, is associated with obesity and insulin resistance. Int. J. Obes. 2006, 30, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Clemente, M.; Claria, J.; Titos, E. The 5-lipoxygenase/leukotriene pathway in obesity, insulin resistance, and fatty liver disease. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Mothe-Satney, I.; Filloux, C.; Amghar, H.; Pons, C.; Bourlier, V.; Galitzky, J.; Grimaldi, P.A.; Feral, C.C.; Bouloumie, A.; van Obberghen, E.; et al. Adipocytes secrete leukotrienes: Contribution to obesity-associated inflammation and insulin resistance in mice. Diabetes 2012, 61, 2311–2319. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.C.; Gijon, M.A. Biosynthesis and metabolism of leukotrienes. Biochem. J. 2007, 405, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Claria, J.; Dalli, J.; Yacoubian, S.; Gao, F.; Serhan, C.N. Resolvin D1 and resolvin D2 govern local inflammatory tone in obese fat. J. Immunol. 2012, 189, 2597–2605. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, J.; Tang, Y.; Kosuri, M.; Bhatnagar, A.; Spite, M. Resolvin D1 decreases adipose tissue macrophage accumulation and improves insulin sensitivity in obese-diabetic mice. FASEB J. 2011, 25, 2399–2407. [Google Scholar] [CrossRef] [PubMed]
- Neuhofer, A.; Zeyda, M.; Mascher, D.; Itariu, B.K.; Murano, I.; Leitner, L.; Hochbrugger, E.E.; Fraisl, P.; Cinti, S.; Serhan, C.N.; et al. Impaired local production of proresolving lipid mediators in obesity and 17-HDHA as a potential treatment for obesity-associated inflammation. Diabetes 2013, 62, 1945–1956. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heemskerk, M.M.; Giera, M.; Bouazzaoui, F.E.; Lips, M.A.; Pijl, H.; Van Dijk, K.W.; Van Harmelen, V. Increased PUFA Content and 5-Lipoxygenase Pathway Expression Are Associated with Subcutaneous Adipose Tissue Inflammation in Obese Women with Type 2 Diabetes. Nutrients 2015, 7, 7676-7690. https://doi.org/10.3390/nu7095362
Heemskerk MM, Giera M, Bouazzaoui FE, Lips MA, Pijl H, Van Dijk KW, Van Harmelen V. Increased PUFA Content and 5-Lipoxygenase Pathway Expression Are Associated with Subcutaneous Adipose Tissue Inflammation in Obese Women with Type 2 Diabetes. Nutrients. 2015; 7(9):7676-7690. https://doi.org/10.3390/nu7095362
Chicago/Turabian StyleHeemskerk, Mattijs M., Martin Giera, Fatiha El Bouazzaoui, Mirjam A. Lips, Hanno Pijl, Ko Willems Van Dijk, and Vanessa Van Harmelen. 2015. "Increased PUFA Content and 5-Lipoxygenase Pathway Expression Are Associated with Subcutaneous Adipose Tissue Inflammation in Obese Women with Type 2 Diabetes" Nutrients 7, no. 9: 7676-7690. https://doi.org/10.3390/nu7095362
APA StyleHeemskerk, M. M., Giera, M., Bouazzaoui, F. E., Lips, M. A., Pijl, H., Van Dijk, K. W., & Van Harmelen, V. (2015). Increased PUFA Content and 5-Lipoxygenase Pathway Expression Are Associated with Subcutaneous Adipose Tissue Inflammation in Obese Women with Type 2 Diabetes. Nutrients, 7(9), 7676-7690. https://doi.org/10.3390/nu7095362