1. Introduction
Complications of atherosclerosis are leading causes of mortality and morbidity worldwide. Substances like folic acid, vitamins B
12 and B
6, or betaine (trimethylglycine) can influence the methionine-homocysteine cycle and thus change concentrations of homocysteine (Hcy) or dimethylglycine (DMG) [
1], which are putative risk factors of atherosclerosis. High Hcy levels appear to be clearly associated with an increased risk of cardiovascular and cerebrovascular disease. However, Hcy does not appear to be as important as other risk factors, such as hypercholesterolemia, smoking, diabetes mellitus, and hypertension [
2]. Despite promising results from observational studies (e.g., [
3]), clinical trials have not confirmed efficiency of supplementation with low and high doses of folic acid and vitamins B
6 or B
12 in decreasing risk of cardiovascular diseases [
4]
. However, folic acid supplementation proved to be effective for stroke prevention [
5,
6,
7]. On the other hand, there is still discussion of whether the clinical trials have the power to prove a potential benefit in a relatively short time and with concurrent hypolipidemic therapy (especially statins), and whether the complexity of influencing factors requires more detailed analysis [
8]. One of the confounding factors is the consumption of alcoholic beverages. It is known that ethanol and its metabolites influence several key enzymes of the methionine-homocysteine cycle (
Figure 1, e.g., they inhibit methionine synthase (MS), activate betaine homocysteine methyltransferase (BHMT), and possibly inhibit methionine adenosyltransferase (MAT)) and thus ethanol has a homocysteine-increasing effect, depletes liver S-adenosylmethionine (SAM), and causes fatty liver disease [
9]. Due to the inhibition of methionine synthase, the BHMT pathway becomes more important as a source of SAM and a determinant of Hcy in alcohol consumers [
10]. Therefore, in alcoholics, betaine theoretically seems to be a more effective methyl group donor than folate. To add more complexity, betaine may decrease the demand for choline methyl groups, thus increasing choline availability for lipid metabolism. Betaine can also support carnitine synthesis and thus a further lipotropic effect [
11]. Furthermore, the transsulfuration pathway of Hcy degradation can be a source of cysteine and glutathione, which are major extracellular and intracellular antioxidants, respectively [
8]. Of note, betaine and SAM supplementation increases the rate of ethanol elimination in rats [
12].
Figure 1.
Effect of ethanol on key enzymes of one-carbon metabolism. Ethanol inhibits methionine synthase (MS), activates betaine homocysteine methyltransferase (BHMT), and possibly inhibits methionine adenosyltransferase (MAT) and several methyltransferases (MTase). Thus, the proposed effect of ethanol is a decrease in Hcy remethylation through the methionine synthase pathway, an increase in remethylation through the BHMT pathway, and a decrease in methylation potential through a decrease in SAM production and inhibition of methylation reactions [
9].
Figure 1.
Effect of ethanol on key enzymes of one-carbon metabolism. Ethanol inhibits methionine synthase (MS), activates betaine homocysteine methyltransferase (BHMT), and possibly inhibits methionine adenosyltransferase (MAT) and several methyltransferases (MTase). Thus, the proposed effect of ethanol is a decrease in Hcy remethylation through the methionine synthase pathway, an increase in remethylation through the BHMT pathway, and a decrease in methylation potential through a decrease in SAM production and inhibition of methylation reactions [
9].
Moreover, several clinical, epidemiological [
13,
14] and experimental studies [
15] have proposed that light-to-moderate alcohol consumption is associated with a decreased risk of atherosclerosis. The relationship between alcohol and vascular risk or total mortality has been repeatedly depicted as a J-shaped curve. After an initial decrease in the vascular risk with increasing amounts of alcohol, the curve reaches a plateau and increases at higher doses [
16,
17,
18]. Not only the amount of alcohol but also the drinking pattern is important,
i.e., protective effects are described in moderate regular drinkers, whereas episodic (binge) heavy drinking has detrimental effects [
18]. Various mechanisms of action have been proposed for the manner in which moderate alcohol consumption affords its protective action. The reduced cardiovascular risk has been in turn explained by the ability of ethanol to increase plasma high-density lipoprotein-cholesterol (HDL) [
19,
20] and apolipoprotein A-I (apoA) [
21], to decrease platelets aggregation and fibrinogen levels [
22], and to promote antioxidant defenses [
23,
24]. It is still not clear whether a particular type of alcoholic beverage (red or white wine, beers or spirits) is important in this context [
25,
26,
27,
28] or not [
29,
30], and there is no consensus on the recommended amount (if any) of daily consumed alcohol [
23,
31,
32]. However, it is obvious that alcohol consumption is one of the leading risk factors for mortality and morbidity worldwide [
18], and any recommendation regarding positive effects of alcohol drinking must be managed extremely cautiously.
In summary, the interactions of ethanol metabolism with the methionine–homocysteine cycle, together with the effects of folic acid, betaine, and vitamins B12 and B6, are not fully understood and intervention trials are needed. We aimed (1) to investigate the effect of alcohol consumption on vitamins and several metabolites involved in one-carbon metabolism; and (2) to find the most effective way of decreasing Hcy during moderate alcohol consumption.
4. Discussion
We present a randomized controlled trial that explores the interaction between moderate alcohol consumption, supplementation of substances important in the methionine methylation cycle, and risk factors of atherosclerosis.
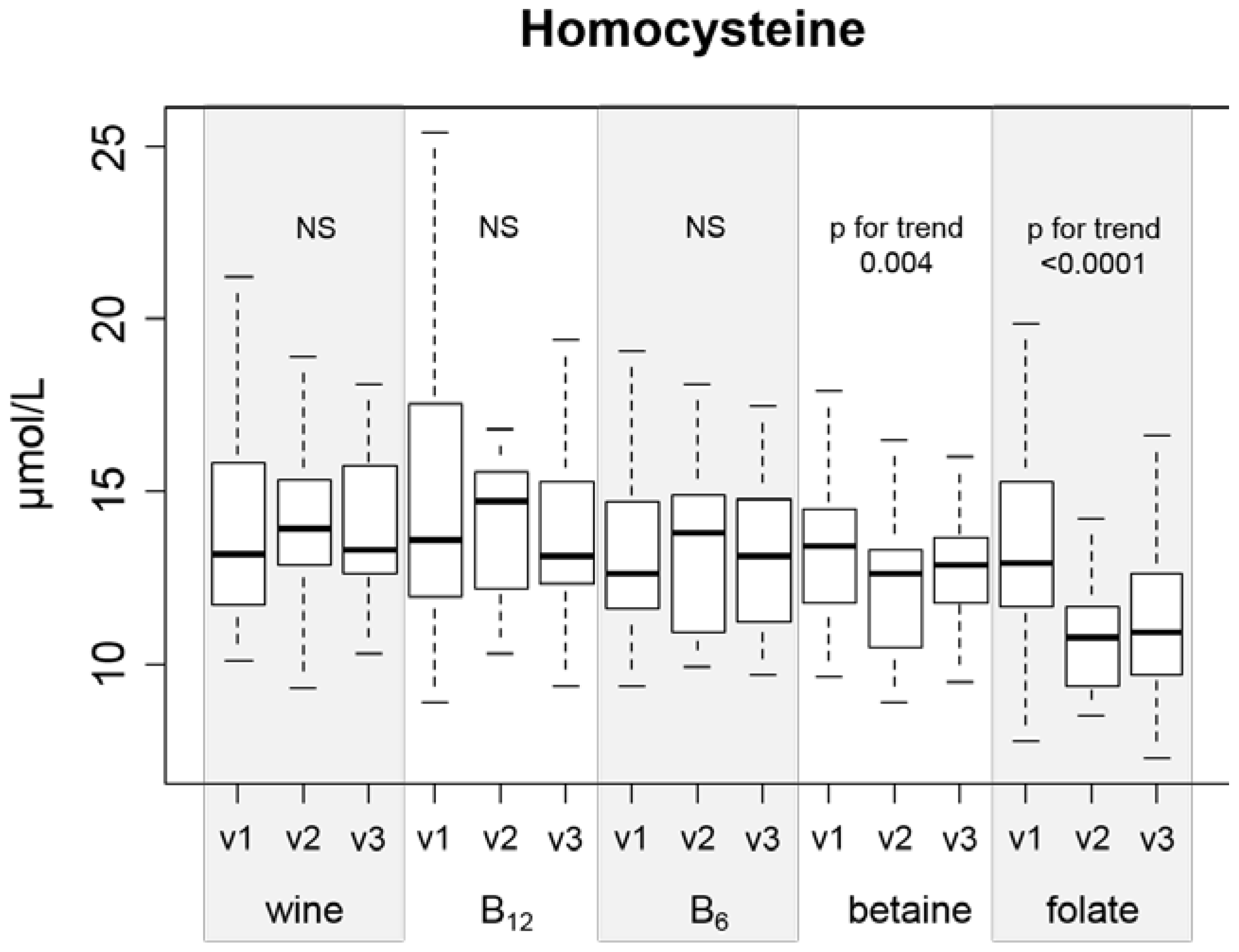
There are two main findings in our study that deserve attention. First, folic acid and betaine are the only substances that can effectively lower Hcy during ingestion of a moderate amount (42 g daily) of ethanol (
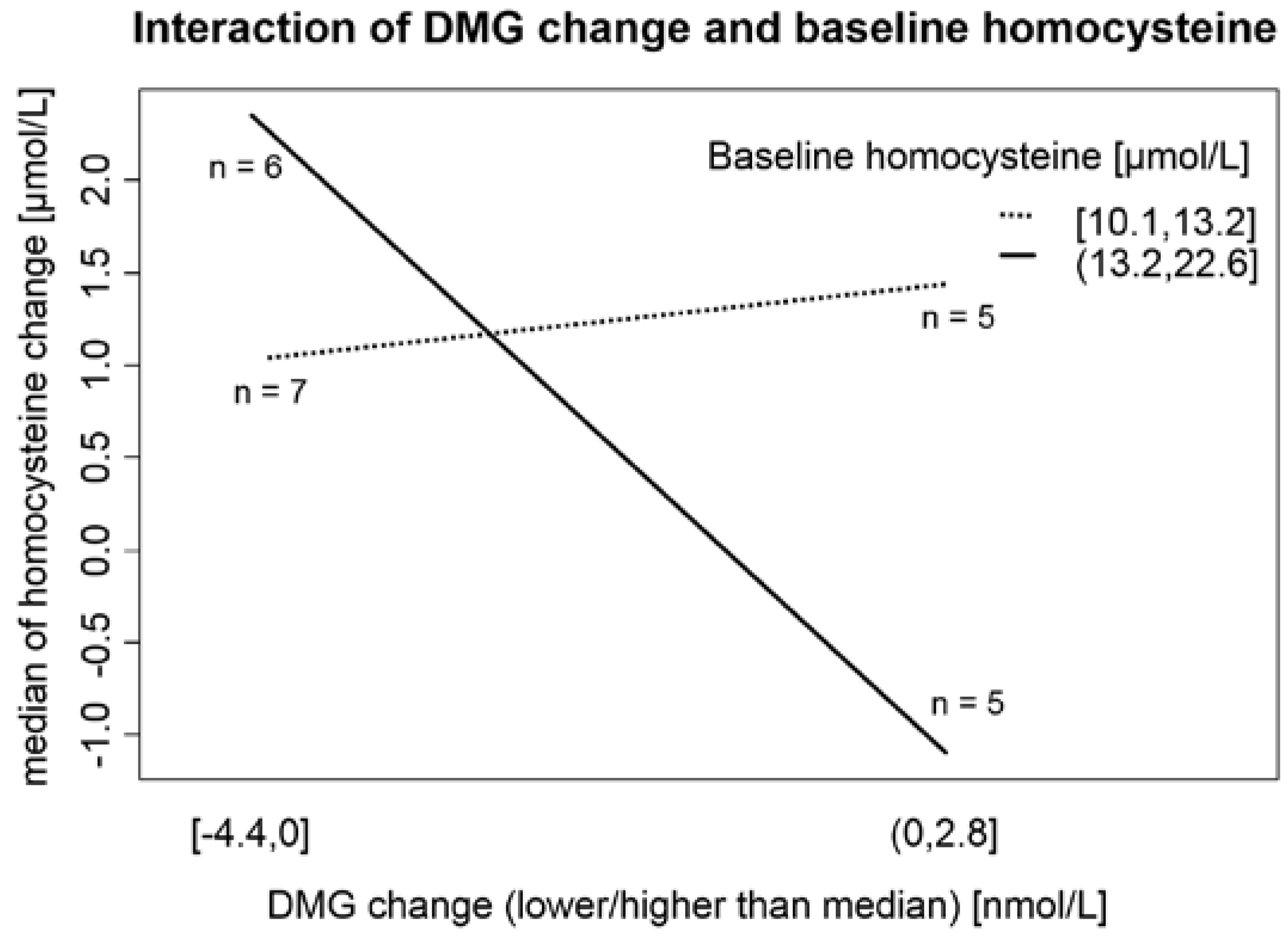
Figure 2). Second, the baseline Hcy value is important in the response of Hcy metabolism enzymatic systems to ethanol ingestion (
Figure 3). To our knowledge, there is no published study with a similar design (concurrent controlled moderate alcohol consumption and supplementation of vitamins).
The observed effect of supplemented substances can be influenced by the selection of the dose of substance. Generally, we used similar vitamin doses to those commonly used in supplementation trials [
36], although the folate dose was somewhat higher (5 mg daily). This dose was recommended by Brouwer [
37] and was intentionally higher to overcome the effect of alcohol on Hcy levels. The ratio between the Recommended Daily Allowance (RDA; or in the case of betaine, mean average daily intake) and supplemented dose was 12.5 (5 mg/0.4 mg), 12 (3 g/0.25 g), 83 (200 µg/2.4 µg), and 26 (40 mg/1.5 mg) for folate, betaine, vitamin B
12, and vitamin B
6 respectively. We do not have dietary intake data for our participants, but the baseline concentrations of involved vitamins reflect intake of supplemented vitamins. The prevalence of presupplementation vitamin deficiency was low in the case of folate (no value <2 µg/L) and vitamin B
12 (one value <110 ng/ L). However, in the case of vitamin B
6 (PLP), the prevalence of plasma values <20 nmol/L (recommended cut-off for adequate intake [
38]) was 71%, and 24% had values of <10 nmol/L. We have no adequate explanation for this unusually high prevalence of vitamin B
6 deficiency in our study population.
We found a decrease in vitamin B
12 and an increase in betaine after 1 month of drinking white wine (without any supplementation), while other vitamins (folate, PLP) remained unchanged. The data on the effect of moderate alcohol consumption on B-vitamin levels are conflicting in the literature. Van der Gaag observed in 11 healthy men drinking red wine, spirits, or beer (40 g of ethanol daily) a decrease in folate concentration only after drinking spirits, with no change in B
12 in all study groups, and an increase in vitamin B
6 after drinking beer and, surprisingly, red wine and spirits too [
39]. Gibson showed a decrease in folate and vitamin B
12 after drinking red wine or spirits (24 g of ethanol daily) in 78 healthy males [
40]. Laufer demonstrated a decrease in vitamin B
12 and no change in folate in 52 postmenopausal women receiving 15 and 30 g daily (ethanol in orange juice) in a diet-controlled crossover trial [
41]. Although our study was not diet-controlled, we can reasonably suppose that eating habits of our participants did not change substantially, thus interference of ethanol (white wine) on the absorption process of vitamin B
12 is the most probable cause. The observed increase in betaine concentration has not been reported in any published study. However, Mar [
42] showed that red and white wines have small amounts of betaine, and we can hypothesize that this could be a reason for the increase.
The effect of folic acid and betaine on Hcy levels in different groups of healthy subjects or patients is well described in the literature [
3,
4,
11]. In our setting (concurrent alcohol consumption), folate and betaine were the only supplemented substances that effectively decreased Hcy, with folate being the most effective. Therefore, we did not confirm the proposed (due to the metabolic interference of ethanol with folate metabolism that was discussed in the Introduction) superiority of betaine. As discussed above, selection of the supplement dose could be the reason for this observation. Our data did not show efficacy of either vitamin B
6 or vitamin B
12 supplementation in decreasing Hcy. This is in concordance with other authors [
37], but van der Gaag [
39] found a correlation between B
6 and Hcy change, which was especially pronounced after beer consumption. This does not directly contradict our results, as beer is a source of folic acid, and it is known that a combination of folic acid and vitamin B
6 is more effective than folic acid itself [
43]. Together with the relatively high supplemented doses of vitamins B
6 and B
12 (relative to RDA), our results underscore the fact that vitamins B
12 and B
6 are not effective as Hcy-lowering agents in this setting.
The hypothesis that even mild alcohol consumption is associated with a total Hcy increase is supported by several studies [
39,
40], including our previous research [
44]. The fact that some studies (and this article) did not show an increase in Hcy after moderate alcohol consumption [
45,
46] indicates that influencing factors are rather complex. However, the effects of ethanol on several enzymes involved in Hcy metabolism (
Figure 1) and the consistent finding of hyperhomocysteinemia in alcoholics [
47] allows us to consider ethanol as a generally hyperhomocysteinemic substance.
In our study, 27% (
n = 31) of subjects can be classified as having moderate hyperhomocysteinemia (concentration of Hcy >15.0 mmol/L) according to presupplementation values. Prevalence of mild hyperhomocysteinemia is a relatively common finding and ranges from a comparable 24% in Greater Tunis [
48] to 68% in northern China [
49]. Plasma Hcy concentration reflects a complex status of remethylation and transsulfuration pathways (including levels of folate, vitamin B
12, betaine, and vitamin B
6). Furthermore, plasma DMG levels are a better marker for the amount of remethylation in the BHMT system than plasma betaine concentration [
11]. Our results suggest that in individuals with higher Hcy (>13.2 µmol/L), the main factor that governs the Hcy change after consumption of 42 g of ethanol daily is DMG (as a marker of Hcy remethylation to methionine mediated by BHMT;
Figure 2). This unique finding from our interventional trial can partially explain the conflicting results of studies observing the association between alcohol consumption and Hcy levels (moderate alcohol consumption as a factor associated with lower plasma Hcy levels in Hordaland study [
50]
vs. alcohol consumption associated with increased plasma Hcy levels [
44,
49,
51]). Svingen
et al. [
1] found in a large (4150 patients) prospective study that high plasma DMG levels enhance the risk of acute myocardial infarction. Unfortunately, the authors did not mention alcohol consumption as a possible confounding factor and 80% of participants were on statin therapy (usually in addition to other drugs, factors known to influence Hcy and DMG levels [
52,
53]). On the other hand, our participants were not undergoing statin or fibrate therapy, and we obtained fasting morning serum samples, thus allowing more controlled and standardized results. Therefore, our results may change the interpretation of DMG as a putative risk factor of atherosclerotic complications.
The results of our study on lipoprotein particles are consistent with various data in the literature [
20]: in the whole study group (regardless of supplementation groups), HDL-cholesterol plus apoA increased, and LDL cholesterols plus apoB and coagulation factor fibrinogen decreased after a month of white wine drinking. There was no statistically significant difference between study groups, and a specific effect of betaine on lipoprotein levels could not be demonstrated. Betaine supplementation showed a lipotropic effect in some studies [
11], and betaine is used in animal breeding to increase lean body mass and in humans to prevent alcoholic [
54] and non-alcoholic steatohepatitis [
55]. The mechanism of this action is not clear, but methylation of active substances (e.g., norepinephrine to epinephrine [
54], synthesis of carnitine [
11] or synthesis of creatine [
56]) and methylation of DNA and subsequent regulation of gene expression (e.g., increased apoB synthesis [
57], activation of peroxisome proliferator-activated receptor-α (PPARα), or an increase in microsomal triglyceride transfer protein [
58]) are probably involved. Sparks [
57] depicted, in an animal model (rats), a rise in apoB mRNA expression after BHMT activation and betaine supplementation that led to increased VLDL and TG production and a decrease in TG in liver tissue. On the other hand, Wang [
58] found other mechanisms of betaine protection against steatohepatitis,
i.e., prevention of increased expression of enzymes involved in fatty acid synthesis (fatty acid synthase, acyl-CoA oxidase) and prevention of the PPARα and microsomal triglyceride transfer protein mRNA increase, which are factors involved in lipoprotein metabolism and fatty acid breakdown. Interestingly,
apoB expression was not influenced by betaine. CRP in concentrations below 10 mg/L (hsCRP) can be used for atherosclerosis risk assessment. Moreover, a J-shaped association between hsCRP and alcohol consumption is described [
59]. One of explanations of this phenomenon is that low alcohol concentrations may inhibit interleukin-6 secretion from adipocytes [
60] and folate can also modify this relation [
61]. We observed a significant positive correlation between initial hsCRP (visist 1) and BMI (
r = 0.45,
p < 0.0001). In the regression model with initial hsCRP as independent and initial BMI, body fat and pre-study alcohol consumption as independent variables, BMI and alcohol consumption were marginally significant (
p = 0.055 and 0.056 resp.; adjusted
R2 = 0.19,
p < 0.0001; data not shown). There were no significant changes of hsCRP in all supplemented groups (
Table 5); therefore, it is not reasonable to seek for a relationship to white wine and supplemented substance administration.
We conclude that the effect of consumed white wine on lipoproteins is “atheroprotective” (decrease in the LDL/HDL ratio) and decreases coagulation by lowering fibrinogen. On the other hand, known “side effects” (increase of liver enzymes, triglycerides and uric acid) of alcohol consumption were not (in our setting) clinically significant: TG and GGT slightly (statistically significantly) increased and ALT remained unchanged. Alcohol consumption can cause fatty liver disease (alcoholic fatty liver disease, AFLD), similarly, obesity, insulin resistance and other conditions are associated with nonalcoholic liver disease (NAFLD) [
62]. In our study, we have no diagnostic measurement (e.g., liver biopsy or ultrasound) to evaluate prevalence of AFLD or NAFLD and possible effect of alcohol consumption and supplemented substances on these entities. Diagnostic performance (AFLD, NAFLD) of laboratory tests and BMI are very limited, however, there were 85 (73%) participants with BMI ≥ 25 kg/m
2 and 19 (16%) participants with BMI ≥ 30 kg/m
2. In addition, 15 participants (12%) had fasting glucose ≥5.6 mmol/L. Thus overweight, obesity and possibly insulin resistance are prevalent in our study population and presence of NAFLD cannot be excluded. A closer look to individual values of ALT, GGT and TG as possible laboratory surrogates for AFLD and NAFLD (
Table 6) reveals that especially increased TG are prevalent in our study population.
Table 6.
Number (percentage) of participants with values of ALT, GGT or TG higher than upper reference limit.
Table 6.
Number (percentage) of participants with values of ALT, GGT or TG higher than upper reference limit.
| | Visit 1 | Visit 2 |
|---|
| ALT >1 µkat/L | 10 (9%) | 9 (8%) |
| GGT >1.3 µkat/L | 2 (2%) | 3 (3%) |
| TG >1.7 mmol/L | 26 (22%) | 29 (25%) |
Some authors [
63] describe a relationship between activity of GGT and ALT. To further elucidate factors influencing changes of ALT, GGT and TG, we built multiple regression models with changes (before and after white wine drinking) of ALT, GGT and TG as dependent variables and starting value (visit 1) of ALT, GGT, TG, BMI, body fat, type of supplemented substance and initial (pre-study) ethanol consumption as explaining variables. Generally, the most important factor influencing changes in abovementioned markers are the starting values of it (e.g., the higher the concentration of TG was before white wine drinking, the lower the increase after white wine drinking,
Table 7). Type of supplemented substance did not influence changes in these markers, thus none of the supplemented substances can be considered as “hepatoprotective” in this setting. Some authors [
64] published indirect evidence that modest alcohol consumption (<10 g/day) can protect against NAFLD. In our study, none of the laboratory markers were influenced by the pre-study consumption of ethanol. However, there was a one-month abstinence from ethanol before visit 1; therefore, a putative effect of modest alcohol consumption could be diminished.
Table 7.
Factors influencing changes of TG, GGT and ALT. Ethanol, pre-study ethanol consumption in grams per day.
Table 7.
Factors influencing changes of TG, GGT and ALT. Ethanol, pre-study ethanol consumption in grams per day.
| | TG Change | GGT Change | ALT Change |
|---|
| Estimate | Std. Error | p | Estimate | Std. Error | p | Estimate | Std. Error | p |
|---|
| Intercept | 1.0451 | 0.4488 | 0.0219 | 0.0741 | 0.0855 | 0.3885 | 0.3548 | 0.1588 | 0.0277 |
| BMI (kg/m2) | −0.0491 | 0.0248 | 0.0502 | −0.0051 | 0.0047 | 0.2795 | −0.0144 | 0.0087 | 0.1016 |
| TG (mmol/L) | −0.3359 | 0.0928 | 0.0005 | −0.0091 | 0.0182 | 0.6188 | −0.0551 | 0.0336 | 0.1036 |
| Body fat (%) | 0.0406 | 0.0181 | 0.0269 | 0.0066 | 0.0034 | 0.0562 | 0.0117 | 0.0063 | 0.0669 |
| ALT (µkat/L) | −0.2276 | 0.1697 | 0.1827 | −0.0397 | 0.0461 | 0.3906 | −0.4328 | 0.0847 | <0.0001 |
| GGT (µkat/L) | −0.0278 | 0.2133 | 0.8966 | −0.1421 | 0.0520 | 0.0075 | 0.0207 | 0.0798 | 0.7956 |
| Group B12 | −0.2247 | 0.1641 | 0.1740 | −0.0237 | 0.0316 | 0.4559 | 0.0576 | 0.0577 | 0.3205 |
| Group B6 | −0.2127 | 0.1643 | 0.1983 | 0.0178 | 0.0313 | 0.5703 | 0.0223 | 0.0576 | 0.7003 |
| Group betain | −0.0357 | 0.1640 | 0.8282 | 0.0304 | 0.0314 | 0.3354 | 0.0480 | 0.0581 | 0.4105 |
| Group folate | −0.1381 | 0.1635 | 0.4002 | 0.0578 | 0.0317 | 0.0714 | 0.0549 | 0.0581 | 0.3467 |
| Ethanol (g/day) | 0.0053 | 0.0030 | 0.0797 | 0.0006 | 0.0006 | 0.3102 | 0.0010 | 0.0011 | 0.3579 |
| | R2 = 0.1214; p = 0.0095 | R2 = 0.1273; p = 0.0095 | R2 = 0.2659; p < 0.0001 |
The liver plays a central role in production and catabolism of Hcy and there is some data that Hcy is higher in patients with NAFLD, [
65] but, in our study, we found no correlation between putative markers of NAFLD (ALT, GGT, TG, BMI) and Hcy (neither in absolute values before white wine drinking, nor comparing changes of these markers before and after wine consumption period; data not shown).
The main limitation of our study is the availability of DMG measurements in the wine-only group and in the betaine-supplementation group. Concentration of DMG was not measured in folate, vitamin B12 and B6 groups. This fact does not allow us to derive conclusions about DMG changes in relation to ingestion of other supplemented substances. Another limitation of our study is that we did not determine genetic factors (e.g., MTHFR mutations) that clearly influence Hcy levels (one-carbon metabolism). Finally, one of the important limitations of our study is that the intervention and follow-up times were short, thus not allowing us to concentrate on the relationship between biochemical markers and outcome (mortality and morbidity) of study subjects.






{kind=link}
{kind=link}
{kind=link}
{kind=link}

