A Mixed Flavonoid-Fish Oil Supplement Induces Immune-Enhancing and Anti-Inflammatory Transcriptomic Changes in Adult Obese and Overweight Women—A Randomized Controlled Trial


 ,
,  ,
, 
Abstract
:1. Introduction
2. Subjects and Methods
2.1. Subjects
2.2. Research Design
2.3. Supplements
2.4. Blood Collection
2.5. Plasma Quercetin Analysis
2.6. Plasma F2-Isoprostanes
2.7. RNA Isolation and Microarray Hybridization
2.8. Microarray Data Analysis
2.9. Pathway and Network Enrichment Analysis
2.10. Statistics
3. Results
3.1. Participants’ Characteristics, Dietary Intake, and Blood Analytes
3.2. Single Gene-Level Analysis
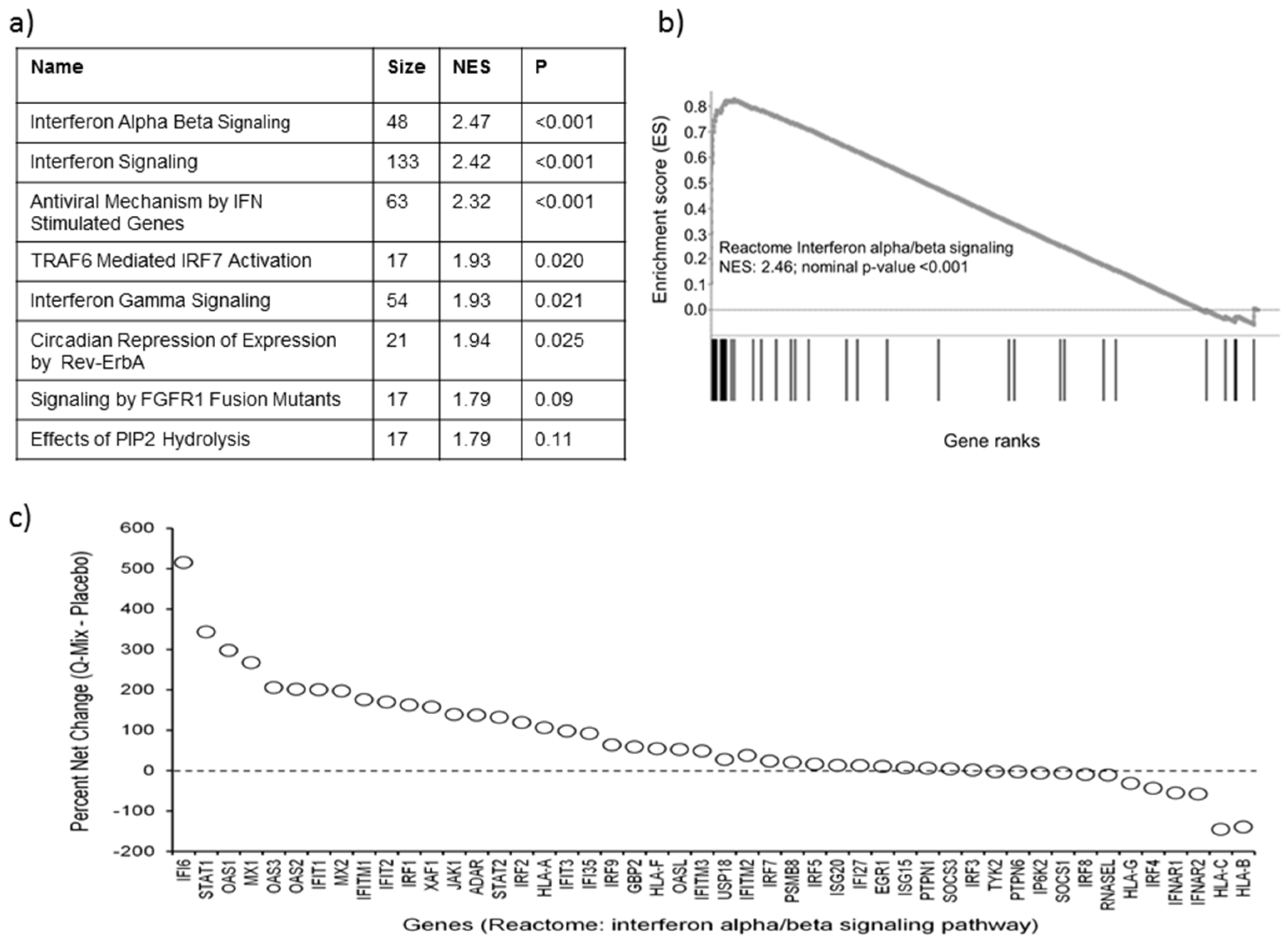
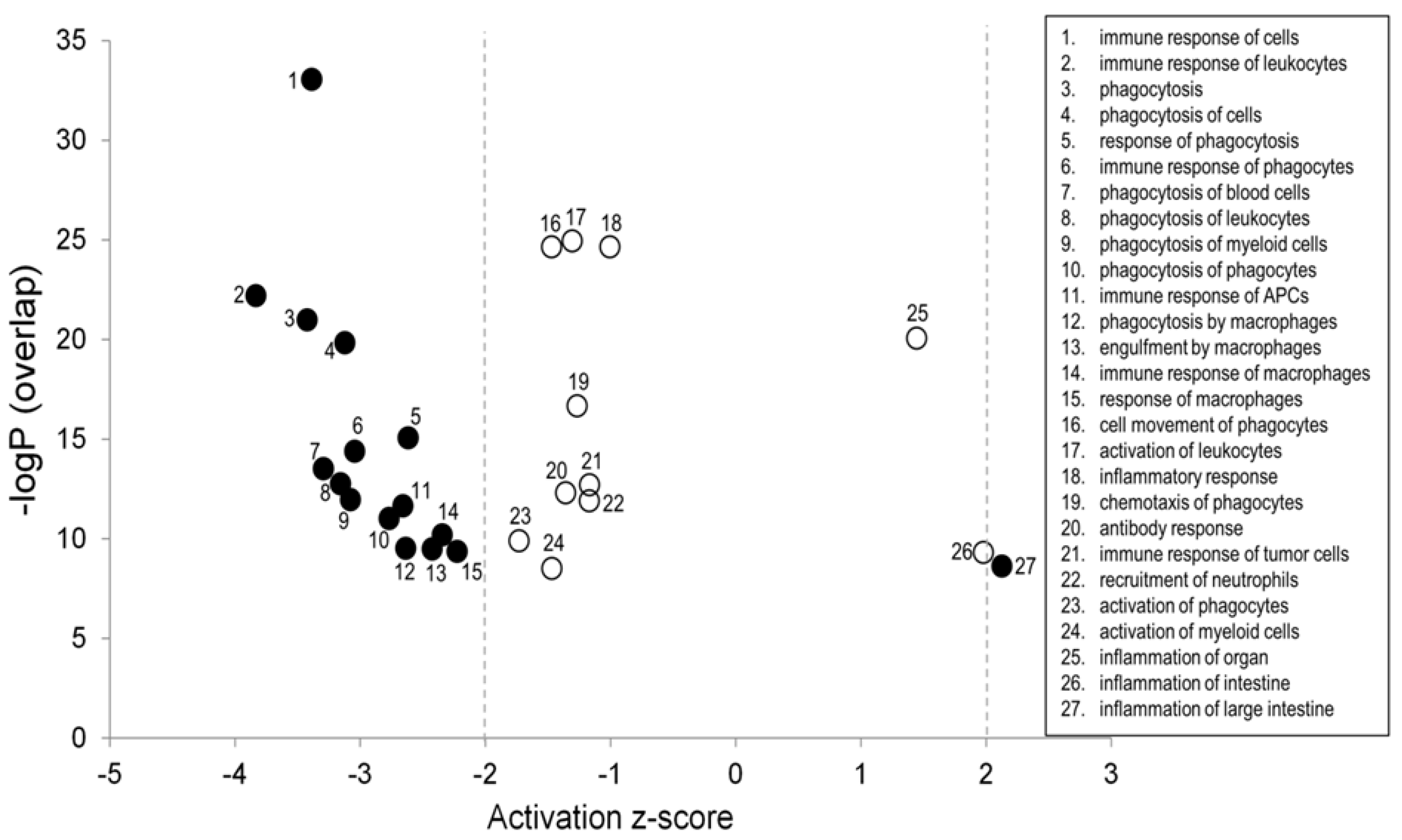
3.3. Pathway Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Barte, J.C.; ter Bogt, N.C.; Bogers, R.P.; Teixeira, P.J.; Blissmer, B.; Mori, T.A.; Bemelmans, W.J. Maintenance of weight loss after lifestyle interventions for overweight and obesity, a systematic review. Obes. Rev. 2010, 11, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Cherniack, E.P. Polyphenols: Planting the seeds of treatment for the metabolic syndrome. Nutrition 2011, 27, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Siriwardhana, N.; Kalupahana, N.S.; Cekanova, M.; LeMieux, M.; Greer, B.; Moustaid-Moussa, N. Modulation of adipose tissue inflammation by bioactive food compounds. J. Nutr. Biochem. 2013, 24, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Bhagwat, S.; Haytowitz, D.B.; Holden, J.M. 2014 USDA Database for the Flavonoid Content of Selected Foods, Release 3.1. U.S. Department of Agriculture, Agricultural Research Service. Nutrient Data Laboratory. Available online: http://www.ars.usda.gov/nutrientdata/flav (accessed on 26 August 2015).
- Heinz, S.A.; Henson, D.A.; Nieman, D.C.; Austin, M.D.; Jin, F. A 12-week supplementation with quercetin does not affect natural killer cell activity, granulocyte oxidative burst activity or granulocyte phagocytosis in female human subjects. Br. J. Nutr. 2010, 104, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Knab, A.M.; Shanely, R.A.; Jin, F.; Austin, M.D.; Sha, W.; Nieman, D.C. Quercetin with vitamin C and niacin does not affect body mass or composition. Appl. Physiol. Nutr. Metab. 2011, 36, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Knab, A.M.; Shanely, R.A.; Henson, D.A.; Jin, F.; Heinz, S.A.; Austin, M.D.; Nieman, D.C. Influence of quercetin supplementation on disease risk factors in community-dwelling adults. J. Am. Diet. Assoc. 2011, 111, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Heinz, S.A.; Henson, D.A.; Austin, M.D.; Jin, F.; Nieman, D.C. Quercetin supplementation and upper respiratory tract infection: A randomized community clinical trial. Pharmacol. Res. 2010, 62, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Grove, K.A.; Lambert, J.D. Laboratory, epidemiological, and human intervention studies show that tea (camellia sinensis) may be useful in the prevention of obesity. J. Nutr. 2010, 140, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Nagao, T.; Hase, T.; Tokimitsu, I. A green tea extract high in catechins reduces body fat and cardiovascular risks in humans. Obesity 2007, 15, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Hase, T.; Komine, Y.; Meguro, S.; Takeda, Y.; Takahashi, H.; Matsui, Y.; Inaoka, S.; Katsuragi, Y.; Tomitsu, J.; Shimasaki, H.; et al. Anti-obesity effects of tea catechins in humans. J. Oleo Sci. 2001, 50, 599–605. [Google Scholar] [CrossRef]
- Basu, A.; Sanchez, K.; Leyva, M.J.; Wu, M.; Betts, N.M.; Aston, C.E.; Lyons, T.J. Green tea supplementation affects body weight, lipids, and lipid peroxidation in obese subjects with metabolic syndrome. J. Am. Coll. Nutr. 2010, 29, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Du, M.; Sanchez, K.; Leyva, M.J.; Betts, N.M.; Blevins, S.; Wu, M.; Aston, C.E.; Lyons, T.J. Green tea minimally affects biomarkers of inflammation in obese subjects with metabolic syndrome. Nutrition 2011, 27, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.L.; Lane, J.; Coverly, J.; Stocks, J.; Jackson, S.; Stephen, A.; Bluck, L.; Coward, A.; Hendrickx, H. Effects of dietary supplementation with the green tea polyphenol epigallocatechin-3-gallate on insulin resistance and associated metabolic risk factors: Randomized controlled trial. Br. J. Nutr. 2009, 101, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Bakker, G.C.; van Erk, M.J.; Pellis, L.; Wopereis, S.; Rubingh, C.M.; Cnubben, N.H.; Kooistra, T.; van Ommen, B.; Hendriks, H.F. An antiinflammatory dietary mix modulates inflammation and oxidative and metabolic stress in overweight men: A nutrigenomics approach. Am. J. Clin. Nutr. 2010, 91, 1044–1059. [Google Scholar] [CrossRef] [PubMed]
- Camuesco, D.; Comalada, M.; Concha, A.; Nieto, A.; Sierra, S.; Xaus, J.; Zarzuelo, A.; Galvez, J. Intestinal anti-inflammatory activity of combined quercitrin and dietary olive oil supplemented with fish oil, rich in EPA and DHA (n-3) polyunsaturated fatty acids, in rats with dss-induced colitis. Clin. Nutr. 2006, 25, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Camuesco, D.; Galvez, J.; Nieto, A.; Comalada, M.; Rodriguez-Cabezas, M.E.; Concha, A.; Xaus, J.; Zarzuelo, A. Dietary olive oil supplemented with fish oil, rich in EPA and DHA (n-3) polyunsaturated fatty acids, attenuates colonic inflammation in rats with dss-induced colitis. J. Nutr. 2005, 135, 687–694. [Google Scholar] [PubMed]
- Nieman, D.C.; Henson, D.A.; Maxwell, K.R.; Williams, A.S.; McAnulty, S.R.; Jin, F.; Shanely, R.A.; Lines, T.C. Effects of quercetin and EGCG on mitochondrial biogenesis and immunity. Med. Sci. Sports Exerc. 2009, 41, 1467–1475. [Google Scholar] [CrossRef] [PubMed]
- McAnulty, S.R.; Nieman, D.C.; McAnulty, L.S.; Lynch, W.S.; Jin, F.; Henson, D.A. Effect of mixed flavonoids, n-3 fatty acids, and vitamin c on oxidative stress and antioxidant capacity before and after intense cycling. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 328–337. [Google Scholar] [PubMed]
- Lines, T.; Quercegen Pharmaceuticals LLC, 144 North Rd., Ste 2100, Sudbury, MA, USA. Personal communication, 2006.
- Nieman, D.C.; Henson, D.A.; Gross, S.J.; Jenkins, D.P.; Davis, J.M.; Murphy, E.A.; Carmichael, M.D.; Dumke, C.L.; Utter, A.C.; McAnulty, S.R.; et al. Quercetin reduces illness but not immune perturbations after intensive exercise. Med. Sci. Sports Exerc. 2007, 39, 1561–1569. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Morrow, J.D.; Yin, H. Quantification of f2-isoprostanes as a reliable index of oxidative stress in vivo using gas chromatography-mass spectrometry (gc-ms) method. Free Radic. Biol. Med. 2009, 47, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics (Oxf. UK) 2003, 4, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Sonna, L.A.; Kuhlmeier, M.M.; Khatri, P.; Chen, D.; Lilly, C.M. A microarray analysis of the effects of moderate hypothermia and rewarming on gene expression by human hepatocytes (HepG2). Cell Stress Chaperones 2010, 15, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Shen-Orr, S.S.; Tibshirani, R.; Khatri, P.; Bodian, D.L.; Staedtler, F.; Perry, N.M.; Hastie, T.; Sarwal, M.M.; Davis, M.M.; Butte, A.J. Cell type-specific gene expression differences in complex tissues. Nat. Methods 2010, 7, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Kuehn, H.; Gould, J.; Tamayo, P.; Mesirov, J.P. Gsea-p: A desktop application for gene set enrichment analysis. Bioinformatics (Oxf. UK) 2007, 23, 3251–3253. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Croft, D.; Mundo, A.F.; Haw, R.; Milacic, M.; Weiser, J.; Wu, G.; Caudy, M.; Garapati, P.; Gillespie, M.; Kamdar, M.R.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2014, 42, D472–D477. [Google Scholar] [CrossRef] [PubMed]
- Pearson, T.A.; Bazzarre, T.L.; Daniels, S.R.; Fair, J.M.; Fortmann, S.P.; Franklin, B.A.; Goldstein, L.B.; Hong, Y.; Mensah, G.A.; Sallis, J.F., Jr.; et al. American heart association guide for improving cardiovascular health at the community level: A statement for public health practitioners, healthcare providers, and health policy makers from the american heart association expert panel on population and prevention science. Circulation 2003, 107, 645–651. [Google Scholar] [PubMed]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type i interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Geller, R.; Andino, R.; Frydman, J. Hsp90 inhibitors exhibit resistance-free antiviral activity against respiratory syncytial virus. PLoS ONE 2013, 8, e56762. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F. Stress-induced changes in immune cell distribution and trafficking: Implications for immunoprotection versus immunopathology. In Neural and Neuroendocrine Mechanisms in Host Defense and Autoimmunity; Welsh, C.J., Meagher, M., Sternberg, E., Eds.; Springer: New York, NY, USA, 2006; pp. 7–25. [Google Scholar]
- Ahmed, M.; Henson, D.A.; Sanderson, M.C.; Nieman, D.C.; Gillitt, N.D.; Lila, M.A. The protective effects of a polyphenol-enriched protein powder on exercise-induced susceptibility to virus infection. Phytother. Res. 2014, 28, 1829–1836. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Son, M.; Ryu, E.; Shin, Y.S.; Kim, J.G.; Kang, B.W.; Cho, H.; Kang, H. Quercetin-induced apoptosis prevents ebv infection. Oncotarget 2015, 6, 12603–12624. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Narayanan, S.; Chang, K.O. Inhibition of influenza virus replication by plant-derived isoquercetin. Antivir. Res. 2010, 88, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, S.Y.; Lee, H.W.; Shin, J.S.; Kim, P.; Jung, Y.S.; Jeong, H.S.; Hyun, J.K.; Lee, C.K. Inhibition of influenza virus internalization by (−)-epigallocatechin-3-gallate. Antivir. Res. 2013, 100, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.; Shirasuna, K.; Usui, F.; Karasawa, T.; Mizushina, Y.; Kimura, H.; Kawashima, A.; Ohkuchi, A.; Matsuyama, S.; Kimura, K.; et al. Interferon-tau attenuates uptake of nanoparticles and secretion of interleukin-1beta in macrophages. PLoS ONE 2014, 9, e113974. [Google Scholar] [CrossRef] [PubMed]
- Luster, A.D.; Alon, R.; von Andrian, U.H. Immune cell migration in inflammation: Present and future therapeutic targets. Nat. Immunol. 2005, 6, 1182–1190. [Google Scholar] [CrossRef] [PubMed]
- Cialdella-Kam, L.; Nieman, D.C.; Sha, W.; Meaney, M.P.; Knab, A.M.; Shanely, R.A. Dose-response to 3 months of quercetin-containing supplements on metabolite and quercetin conjugate profile in adults. Br. J. Nutr. 2013, 109, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Rudkowska, I.; Paradis, A.M.; Thifault, E.; Julien, P.; Tchernof, A.; Couture, P.; Lemieux, S.; Barbier, O.; Vohl, M.C. Transcriptomic and metabolomic signatures of an n-3 polyunsaturated fatty acids supplementation in a normolipidemic/normocholesterolemic caucasian population. J. Nutr. Biochem. 2013, 24, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Bouwens, M.; van de Rest, O.; Dellschaft, N.; Bromhaar, M.G.; de Groot, L.C.; Geleijnse, J.M.; Muller, M.; Afman, L.A. Fish-oil supplementation induces antiinflammatory gene expression profiles in human blood mononuclear cells. Am. J. Clin. Nutr. 2009, 90, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Stahl, F.; Mutz, K.O.; Scheper, T.; Hahn, A.; Schuchardt, J.P. Transcriptome-based identification of antioxidative gene expression after fish oil supplementation in normo- and dyslipidemic men. Nutr. Metab. (Lond.) 2012, 9, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, S.; Stahl, F.; Mutz, K.-O.; Scheper, T.; Hahn, A.; Schuchardt, J.P. Different gene expression profiles in normo- and dyslipidemic men after fish oil supplementation: Results from a randomized controlled trial. Lipids Health Dis. 2012, 11, 105–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skulas-Ray, A.C. Omega-3 fatty acids and inflammation: A perspective on the challenges of evaluating efficacy in clinical research. Prostaglandins Other Lipid Mediat. 2015, 116–117, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gallego, J.; Garcia-Mediavilla, M.V.; Sanchez-Campos, S.; Tunon, M.J. Fruit polyphenols, immunity and inflammation. Br. J. Nutr. 2010, 104 (Suppl. S3), S15–S27. [Google Scholar] [CrossRef] [PubMed]
- Van Duynhoven, J.; Vaughan, E.E.; Jacobs, D.M.; Kemperman, R.A.; van Velzen, E.J.; Gross, G.; Roger, L.C.; Possemiers, S.; Smilde, A.K.; Dore, J.; et al. Metabolic fate of polyphenols in the human superorganism. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4531–4538. [Google Scholar] [CrossRef] [PubMed]
- Roowi, S.; Stalmach, A.; Mullen, W.; Lean, M.E.; Edwards, C.A.; Crozier, A. Green tea flavan-3-ols: Colonic degradation and urinary excretion of catabolites by humans. J. Agric. Food Chem. 2010, 58, 1296–1304. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Clifford, M.N. Colonic metabolites of berry polyphenols: The missing link to biological activity? Br. Nutr. 2010, 104 (Suppl. S3), S48–S66. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Q-Mix (n = 24) | Placebo (n = 24) | p † | |||
|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||
| Age (year) | 55.3 | 7.3 | 56.9 | 9.3 | 0.4919 |
| Height (m) | 1.64 | 0.06 | 1.63 | 0.06 | 0.5007 |
| Body mass (kg) | |||||
| Pre-study | 89.5 | 14.1 | 82.5 | 14.5 | 0.5349 |
| Post-study | 90.7 | 14.2 | 83.3 | 14.6 | |
| BMI (kg/m2) | 33.3 | 5.9 | 31.0 | 5.4 | 0.1692 |
| Body fat (%) | 37.0 | 4.7 | 35.3 | 5.7 | 0.2974 |
| Inflammatory Markers | Time | Q-Mix (n = 24) | Placebo (n = 24) | p † | ||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | |||
| GM-CSF (pg/mL) | Pre | 0.73 | 0.58 | 0.94 | 1.32 | 0.3367 |
| Post | 0.78 | 0.56 | 0.90 | 1.14 | ||
| IFN-γ (pg/mL) | Pre | 1.16 | 0.82 | 1.13 | 0.64 | 0.2288 |
| Post | 1.51 | 1.09 | 1.22 | 0.54 | ||
| IL-1β (pg/mL) | Pre | 0.16 | 0.17 | 0.20 | 0.24 | 0.5825 |
| Post | 0.18 | 0.19 | 0.25 | 0.25 | ||
| IL-2 (pg/mL) | Pre | 0.72 | 0.68 | 0.61 | 0.48 | 0.8645 |
| Post | 0.69 | 0.69 | 0.60 | 0.44 | ||
| IL-6 (pg/mL) | Pre | 1.47 | 0.97 | 1.63 | 1.69 | 0.1527 |
| Post | 1.35 | 0.74 | 1.78 | 1.72 | ||
| IL-8 (pg/mL) | Pre | 3.58 | 2.54 | 3.70 | 2.35 | 0.4471 |
| Post | 3.63 | 2.00 | 4.09 | 2.44 | ||
| IL-10 (pg/mL) | Pre | 4.06 | 9.77 | 2.20 | 2.86 | 0.6386 |
| Post | 4.48 | 12.02 | 2.36 | 2.53 | ||
| IL-12p70 (pg/mL) | Pre | 1.78 | 1.95 | 1.72 | 2.17 | 0.6003 |
| Post | 1.81 | 2.20 | 1.95 | 2.02 | ||
| TNF-α (pg/mL) | Pre | 5.52 | 1.83 | 5.63 | 2.40 | 0.5146 |
| Post | 5.32 | 1.58 | 5.64 | 2.23 | ||
| CRP (mg/L) | Pre | 4.64 | 5.14 | 4.69 | 7.29 | 0.268 |
| Post | 5.60 | 4.27 | 6.81 | 8.64 | ||
| Oxidative Stress Marker | ||||||
| F2-isoprostanes (pg/mL) | Pre | 98.6 | 28.2 | 117.4 | 34.3 | 0.2727 |
| Post | 96.4 | 26.0 | 106.1 | 23.0 | ||
| Dietary Intake | Time | Q-Mix (n = 24) | Placebo (n = 24) | p † | ||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | |||
| Energy (kJ/day) | Pre | 8363 | 2161 | 8016 | 1727 | 0.3626 |
| Post | 8081 | 2112 | 7223 | 2348 | ||
| Carbohydrates (% total energy) | Pre | 50 | 9.0 | 54 | 7.7 | 0.2988 |
| Post | 51 | 8.6 | 52 | 8.0 | ||
| Protein (% total energy) | Pre | 15 | 3.6 | 16 | 2.8 | 0.0990 |
| Post | 14 | 3.4 | 17 | 3.3 | ||
| Fat (% total energy) | Pre | 35 | 7.8 | 32 | 7.8 | 0.5048 |
| Post | 34 | 7.4 | 32 | 6.8 | ||
| Fiber (g/day) | Pre | 18 | 7.7 | 20 | 11.1 | 0.2078 |
| Post | 19 | 9.2 | 18 | 8.5 | ||
| Lipid Panel | ||||||
| Cholesterol (mg/dL) | Pre | 201 | 32.3 | 210 | 47.2 | 0.4144 |
| Post | 199 | 35.5 | 213 | 51.6 | ||
| LDL-Cholesterol (mg/dL) | Pre | 127 | 29.2 | 131 | 44.4 | 0.4218 |
| Post | 125 | 31.1 | 133 | 51.3 | ||
| HDL-Cholesterol (mg/dL) | Pre | 51.1 | 11.9 | 54.7 | 11.4 | 0.3122 |
| Post | 52.9 | 13.1 | 58.1 | 11.8 | ||
| Triglycerides (mg/dL) | Pre | 116 | 34.7 | 124 | 51.2 | 0.6909 |
| Post | 109 | 40.5 | 112 | 37.5 | ||
| Comprehensive Metabolic Panel | ||||||
| Alkaline phosphatase (IU/L) | Pre | 72.6 | 14.2 | 74.5 | 13.9 | 0.1086 |
| Post | 73.2 | 14.0 | 78.5 | 14.6 | ||
| Aspartate aminotransferase (AST; IU/L) | Pre | 21.5 | 5.9 | 25.3 | 9.5 | 0.3035 |
| Post | 22.2 | 7.2 | 24.1 | 9.6 | ||
| Blood Urea Nitrogen (BUN; mg/dL) | Pre | 10.3 | 2.7 | 12.2 | 4.1 | 0.7283 |
| Post | 10.7 | 2.7 | 12.3 | 3.8 | ||
| Creatinine (mg/dL) | Pre | 0.816 | 0.14 | 0.836 | 0.24 | 0.6786 |
| Post | 0.812 | 0.13 | 0.846 | 0.24 | ||
| Glucose (mg/dL) | Pre | 93.8 | 9.7 | 97.0 | 10.2 | 0.6234 |
| Post | 95.2 | 11.3 | 99.7 | 14.3 | ||
| Sodium (mEq/L) | Pre | 139 | 1.8 | 139 | 2.6 | 0.7400 |
| Post | 140 | 1.9 | 140 | 2.5 | ||
| Total Bilirubin (mg/dL) | Pre | 0.688 | 0.18 | 0.667 | 0.18 | 0.6036 |
| Post | 0.679 | 0.20 | 0.692 | 0.21 | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cialdella-Kam, L.; Nieman, D.C.; Knab, A.M.; Shanely, R.A.; Meaney, M.P.; Jin, F.; Sha, W.; Ghosh, S. A Mixed Flavonoid-Fish Oil Supplement Induces Immune-Enhancing and Anti-Inflammatory Transcriptomic Changes in Adult Obese and Overweight Women—A Randomized Controlled Trial. Nutrients 2016, 8, 277. https://doi.org/10.3390/nu8050277
Cialdella-Kam L, Nieman DC, Knab AM, Shanely RA, Meaney MP, Jin F, Sha W, Ghosh S. A Mixed Flavonoid-Fish Oil Supplement Induces Immune-Enhancing and Anti-Inflammatory Transcriptomic Changes in Adult Obese and Overweight Women—A Randomized Controlled Trial. Nutrients. 2016; 8(5):277. https://doi.org/10.3390/nu8050277
Chicago/Turabian StyleCialdella-Kam, Lynn, David C. Nieman, Amy M. Knab, R. Andrew Shanely, Mary Pat Meaney, Fuxia Jin, Wei Sha, and Sujoy Ghosh. 2016. "A Mixed Flavonoid-Fish Oil Supplement Induces Immune-Enhancing and Anti-Inflammatory Transcriptomic Changes in Adult Obese and Overweight Women—A Randomized Controlled Trial" Nutrients 8, no. 5: 277. https://doi.org/10.3390/nu8050277
APA StyleCialdella-Kam, L., Nieman, D. C., Knab, A. M., Shanely, R. A., Meaney, M. P., Jin, F., Sha, W., & Ghosh, S. (2016). A Mixed Flavonoid-Fish Oil Supplement Induces Immune-Enhancing and Anti-Inflammatory Transcriptomic Changes in Adult Obese and Overweight Women—A Randomized Controlled Trial. Nutrients, 8(5), 277. https://doi.org/10.3390/nu8050277