Inositol Hexaphosphate and Inositol Inhibit Colorectal Cancer Metastasis to the Liver in BALB/c Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Lines and Cell Cultures
2.3. Animals and Treatment
2.4. Liver Metastasis Animal Model
2.5. Grouping and Treatment of Animals
2.6. Treatment of Experimental Animals
2.7. Western Blot Analysis
2.8. Immunohistochemical Analysis
2.9. Real-Time PCR
2.10. Statistical Analysis
3. Results
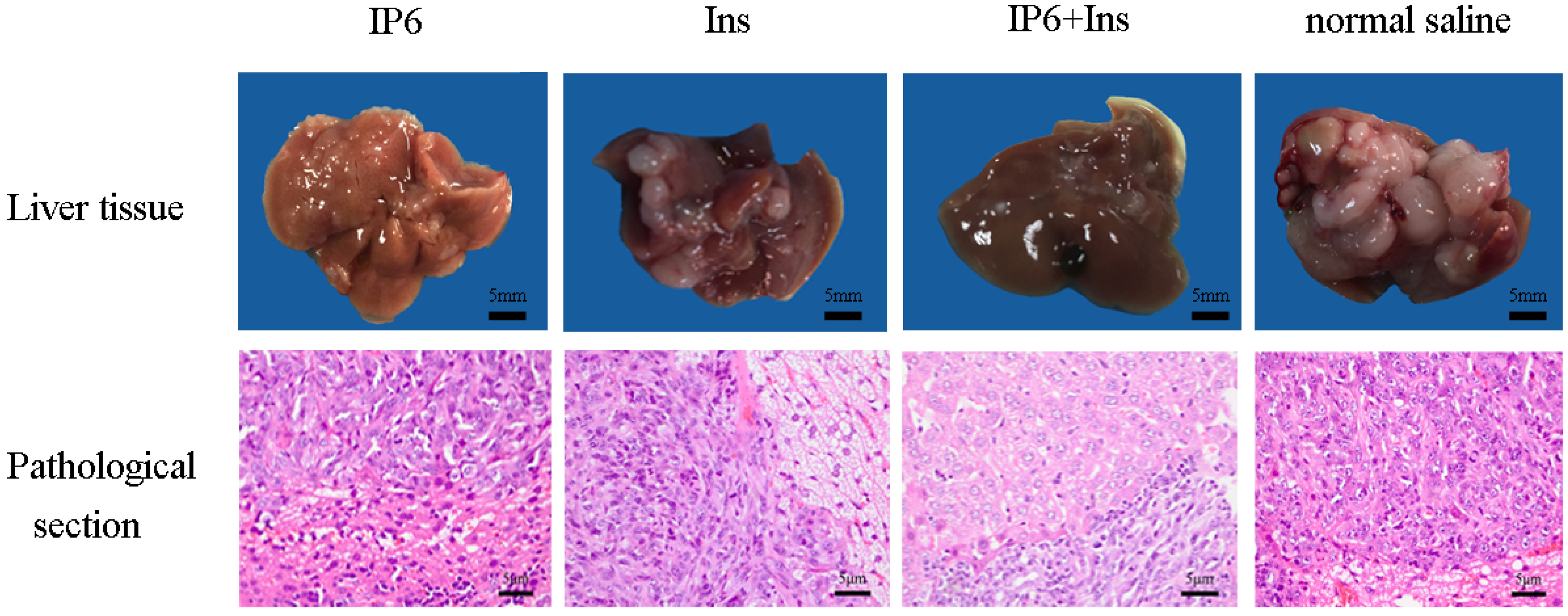
3.1. Occurrence of Liver Metastasis in CRC in BALB/c Mice
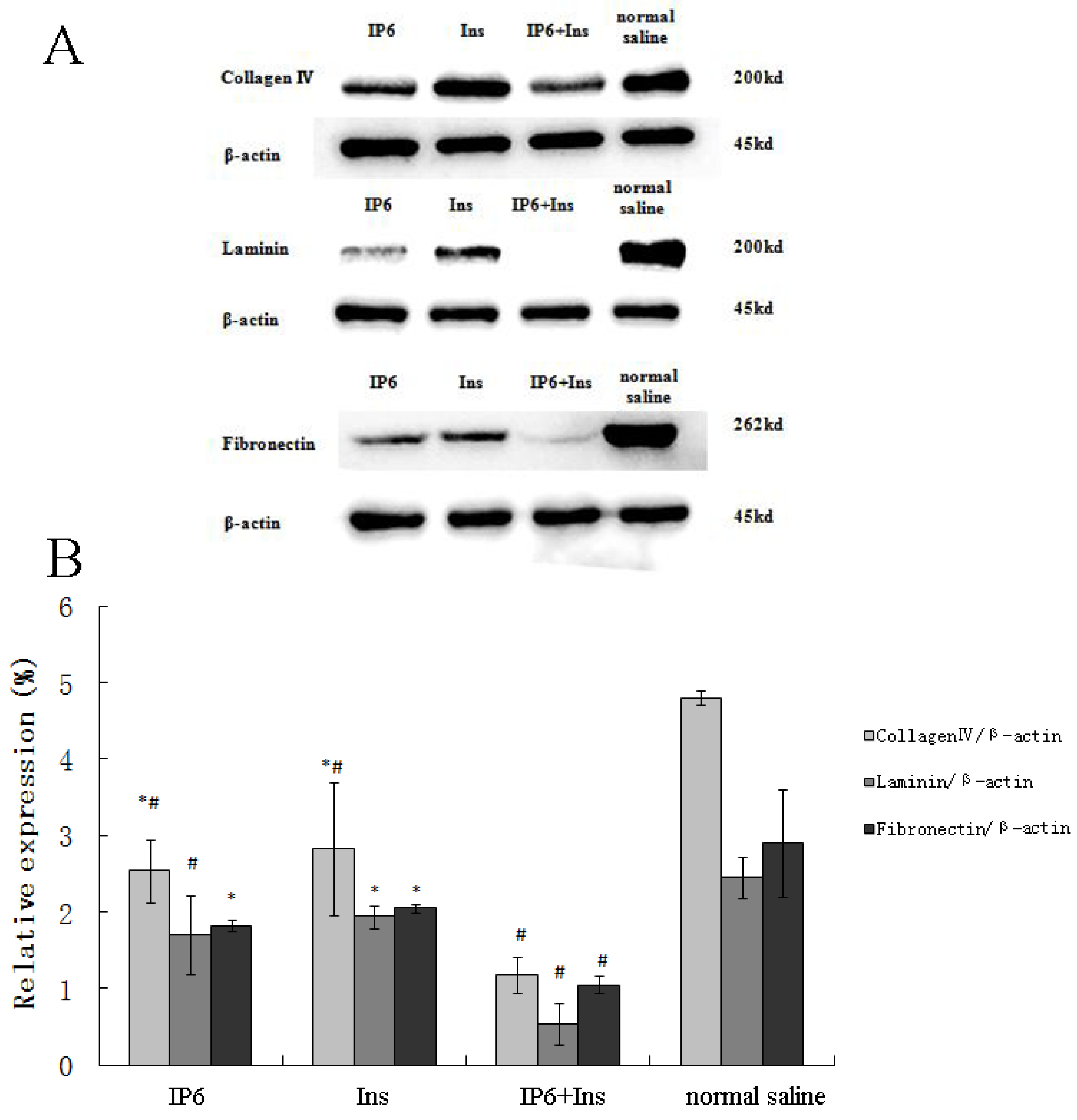
3.2. Protein Expression of Collagen IV, LN and FN in Liver Tissue
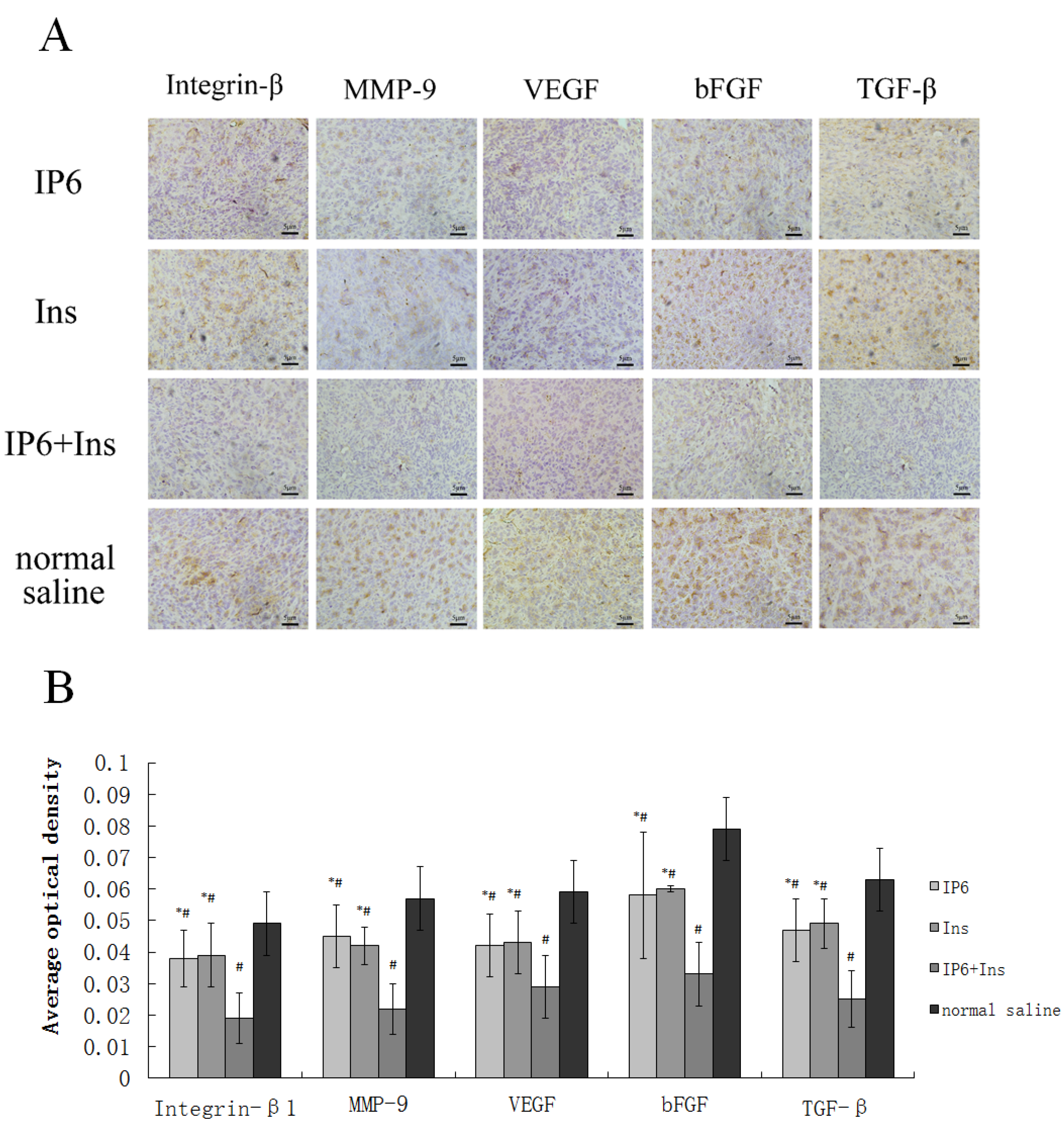
3.3. Immunohistochemical Analyses of Integrin-β1, MMP-9, VEGF, bFGF, and TGF-β Protein Expression in Liver Tissue
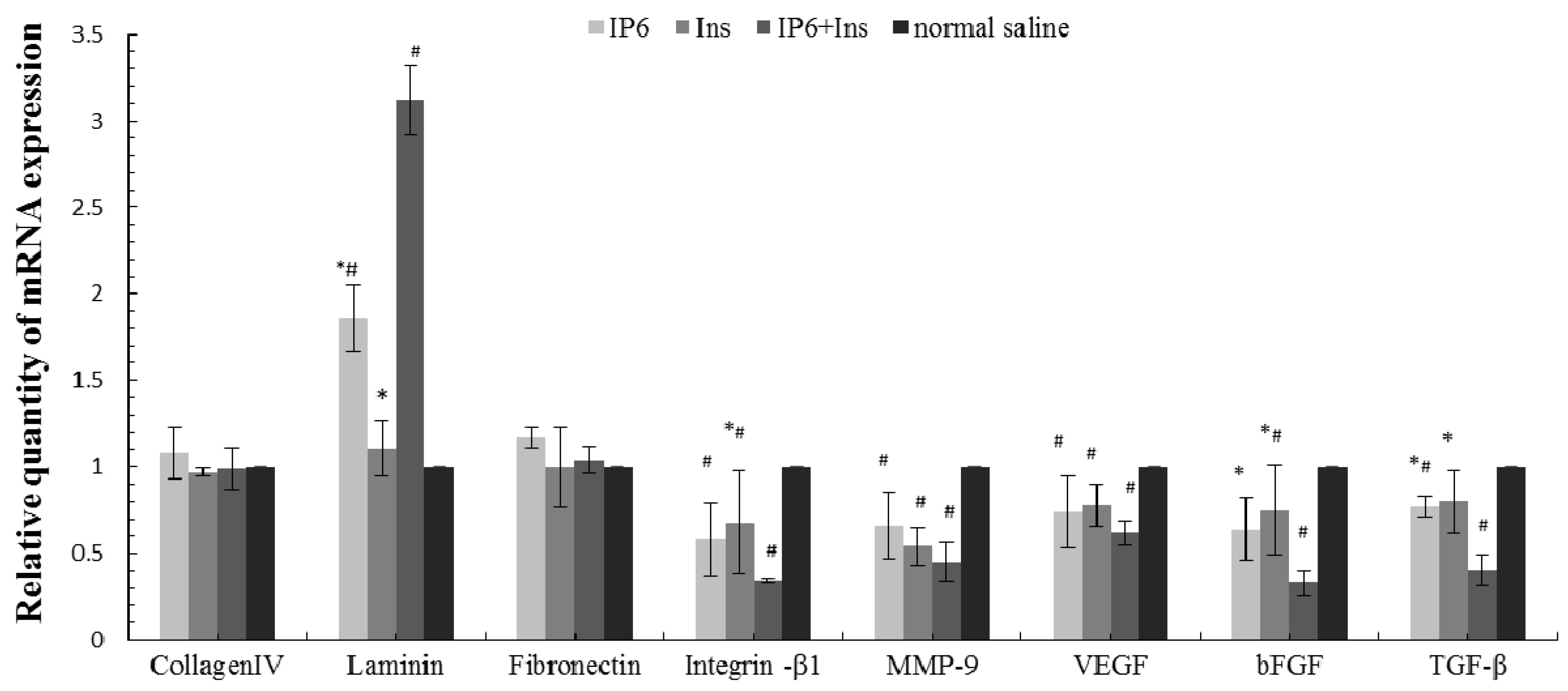
3.4. Real-Time PCR Analyses of Collagen IV, LN, FN, Integrin-β1, MMP-9, VEGF, bFGF, and TGF-β mRNA Expression in Liver Tissue
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 1, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Mantke, R.; Schmidt, U.; Wolff, S.; Kube, R.; Lippert, H. Incidence of synchronous liver metastases in patients with colorectal cancer in relationship to clinico-pathologic characteristics. Results of a German prospective multicentre observational study. Eur. J. Surg. Oncol. 2012, 3, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Cameron, J.L.; Riall, T.S.; Coleman, J.; Belcher, K.A. One thousand consecutive pancreaticoduodenectomies. Ann. Surg. 2006, 1, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Deliliers, G.L.; Servida, F.; Fracchiolla, N.S.; Ricci, C.; Borsotti, C.; Colombo, G.; Soligo, D. Effect of inositol hexaphosphate (IP6) on human normal and leukaemic haematopoietic cells. Br. J. Haematol. 2002, 117, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Schlemmer, U.; Frolich, W.; Prieto, R.M.; Grases, F. Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 2009, 53, S330–S375. [Google Scholar] [CrossRef] [PubMed]
- Nawrocka-Musial, D.; Latocha, M. [Phytic acid—Anticancer nutriceutic]. Pol. Merkur. Lek. 2012, 193, 43–47. [Google Scholar]
- Hirose, M.; Ozaki, K.; Takaba, K.; Fukushima, S.; Shirai, T.; Ito, N. Modifying effects of the naturally occurring antioxidants gamma-oryzanol, phytic acid, tannic acid and n-tritriacontane-16, 18-dione in a rat wide-spectrum organ carcinogenesis model. Carcinogenesis 1991, 10, 1917–1921. [Google Scholar] [CrossRef]
- Vucenik, I.; Tantivejkul, K.; Zhang, Z.S.; Cole, K.E.; Saied, I.; Shamsuddin, A.M. IP6 in treatment of liver cancer. I. IP6 inhibits growth and reverses transformed phenotype in HepG2 human liver cancer cell line. Anticancer Res. 1998, 18, 4083–4090. [Google Scholar] [PubMed]
- Vucenik, I.; Zhang, Z.S.; Shamsuddin, A.M. IP6 in treatment of liver cancer. II. Intra-tumoral injection of IP6 regresses pre-existing human liver cancer xenotransplanted in nude mice. Anticancer Res. 1998, 18, 4091–4096. [Google Scholar] [PubMed]
- Lee, H.J.; Lee, S.A.; Choi, H. Dietary administration of inositol and/or inositol-6-phosphate prevents chemically-induced rat hepatocarcinogenesis. Asian Pac. J. Cancer Prev. 2005, 1, 41–47. [Google Scholar]
- Dang, N.T.; Mukai, R.; Yoshida, K.; Ashida, H. D-pinitol and myo-inositol stimulate translocation of glucose transporter 4 in skeletal muscle of C57BL/6 mice. Biosci. Biotechnol. Biochem. 2010, 5, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Setoguchi, T.; Katayama, T. Effects of dietary myo-inositol, d-chiro-inositol and l-chiro-inositol on hepatic lipids with reference to the hepatic myo-inositol status in rats fed on 1,1,1-trichloro-2,2-bis (p-chlorophenyl) ethane. Biosci. Biotechnol. Biochem. 2006, 11, 2766–2770. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, A.; Giordano, D.; Corrado, F.; Pintaudi, B.; Interdonato, M.L.; Vieste, G.D.; Benedetto, A.D.; D’Anna, R. One-year effects of myo-inositol supplementation in postmenopausal women with metabolic syndrome. Climacteric 2012, 5, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Shamsuddin, A.M.; Ullah, A.; Chakravarthy, A.K. Inositol and inositol hexaphosphate suppress cell proliferation and tumor formation in CD-1 mice. Carcinogenesis 1989, 8, 1461–1463. [Google Scholar] [CrossRef]
- Vucenik, I.; Yang, G.Y.; Shamsuddin, A.M. Inositol hexaphosphate and inositol inhibit DMBA-induced rat mammary cancer. Carcinogenesis 1995, 5, 1055–1058. [Google Scholar] [CrossRef]
- Vucenik, I.; Tomazic, V.J.; Fabian, D.; Shamsuddin, A.M. Antitumor activity of phytic acid (inositol hexaphosphate) in murine transplanted and metastatic fibrosarcoma, a pilot study. Cancer Lett. 1992, 1, 9–13. [Google Scholar] [CrossRef]
- Bacic, I.; Druzijanic, N.; Karlo, R.; Skific, I.; Jagic, S. Efficacy of IP6 + inositol in the treatment of breast cancer patients receiving chemotherapy: Prospective, randomized, pilot clinical study. J. Exp. Clin. Cancer Res. 2010, 12. [Google Scholar] [CrossRef] [PubMed]
- Vucenik, I.; Shamsuddin, A.M. Protection against cancer by dietary IP6 and inositol. Nutr. Cancer 2006, 2, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Esin, E.; Yalcin, S. Maintenance strategy in metastatic colorectal cancer: A systematic review. Cancer Treat. Rev. 2016, 42, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, F.; de Vita, F.; Colucci, G.; Pisconti, S. Maintenance therapy in colon cancer. Cancer Treat. Rev. 2010, 36, S42–S45. [Google Scholar] [CrossRef]
- Kitadai, Y.; Sasaki, T.; Kuwai, T.; Nakamura, T.; Bucana, C.D.; Fidler, I.J. Targeting the expression of platelet-derived growth factor receptor by reactive stroma inhibits growth and metastasis of human colon carcinoma. Am. J. Pathol. 2006, 6, 2054–2065. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Song, Y.; Lan, K.T. Effects of Phytic acid on colorectal carcinogenesis and natural killer cells activity in rats. Acta Mech. Sin. 2004, 4, 308–310, 314. [Google Scholar]
- Liu, G.; Song, Y.; Cui, L.; Wen, Z.; Lu, X. Inositol hexaphosphate suppresses growth and induces apoptosis in HT-29 colorectal cancer cells in culture: PI3K/Akt pathway as a potential target. Int. J. Clin. Exp. Pathol. 2015, 2, 1402–1410. [Google Scholar]
- Liu, M.; Meng, X.F.; Song, Y.; Yang, F.G. Effects of phytic acid on apoptosis of human heptoma cell line HepG2 induced through mitochondrial pathway. Chin. J. Public Health 2014, 10, 1285–1288. [Google Scholar]
- Song, Y.; Zhang, Z.; Ma, B.L. Effect of inositol hexaphosphate (IP6) on cell cycle of HT-29 human colon carcinoma cell line. Acta Mech. Sin. 2007, 3, 246–249. [Google Scholar]
- Li, X.; Song, Y. Effects of phytic acid on cell cycle and the related protein expression in human hepatoma cell line HepG2 in vitro. Acta Nutr. Sin. 2011, 5, 476–479. [Google Scholar]
- Tian, Y.; Song, Y. Effects of inositol hexaphosphate on proliferation of HT-29 human colon carcinoma cell line. J. World J. Gastroenterol. 2006, 26, 4137–4142. [Google Scholar] [CrossRef]
- Yang, W.P.; Song, Y.; Meng, X.F. Effect of inositol hexaphosphate on multiplication of liver cancer cell HepG2. Acta Acad. Med. Qingdao Univ. 2011, 4, 293–295. [Google Scholar]
- Yang, W.P.; Leng, C.L.; Song, Y. Growth inhibition and mechanism of phytic acid on HepG2 cells. Chin. J. Cancer Prev. Treat. 2013, 13, 982–985. [Google Scholar]
- Liu, X.Y.; Song, Y.; Zhang, X.N. The experimental study of cell-induced differentiation by inositol hexaphosphate in HT-29 human colon carcinoma cell line. Acta Nutr. Sin. 2009, 4, 388–391. [Google Scholar]
- Song, L.; Leng, C.L.; Song, Y. Effect of inositol hexaphosphate on apoptosis of HT-29 human colon carcinoma cells and expression of apoptosis related gene. Chin. J. Public Health 2010, 4, 419–420. [Google Scholar]
- Meng, X.F.; Song, Y.; Yang, W.P. Apoptosis of heptoma cell line HepG2 induced by IP6 and its effects on expressions of proteins Bcl-2 and Bax. Acta Acad. Med. Qingdao Univ. 2011, 4, 311–313. [Google Scholar]
- Song, Y.; Li, M. Inhibition effect of IP6 on subcutaneous transplantation of colon cancer in nude mice. Chin. J. Cancer Prev. Treat. 2011, 23, 1821–1824. [Google Scholar]
- Herszenyi, L.; Hritz, I.; Lakatos, G.; Varga, M.Z.; Tulassay, Z. The behavior of matrix metalloproteinases and their inhibitors in colorectal cancer. Int. J. Mol. Sci. 2012, 10, 13240–13263. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Ngoc, K.V.; Cheung, K.J.; Brenot, A.; Shamir, E.R.; Gray, R.S.; Hines, W.C.; Yaswen, P.; Werb, Z.; Ewald, A.J. ECM microenvironment regulates collective migration and local dissemination in normal and malignant mammary epithelium. Proc. Natl. Acad. Sci. USA 2012, 39, E2595–E2604. [Google Scholar] [CrossRef] [PubMed]
- Naba, A.; Clauser, K.R.; Whittaker, C.A.; Carr, S.A.; Tanabe, K.K.; Hynes, R.O. Extracellular matrix signatures of human primary metastatic colon cancers and their metastases to liver. BMC Cancer 2014, 14, 518. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.K.; Ma, Q.; Chen, S.; Sha, H.; Ding, S. Relationship of fibronectin and CD44v6 expression with invasive growth and metastasis of liver cancer. Cancer Invest. 2009, 3, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Aoki, S.; Nakanishi, Y.; Akimoto, S.; Moriya, Y.; Yoshimura, K.; Kitajima, M.; Sakamoto, M.; Hirohashi, S. Prognostic significance of laminin-5 gamma2 chain expression in colorectal carcinoma: Immunohistochemical analysis of 103 cases. Dis. Colon Rectum 2002, 11, 1520–1527. [Google Scholar] [CrossRef]
- Spenle, C.; Lefebvre, O.; Lacroute, J.; Mechine-Neuville, A.; Barreau, F.; Blottiere, H.M.; Duclos, B.; Arnold, C.; Hussenet, T.; Hemmerlé, J.; et al. The laminin response in inflammatory bowel disease: Protection or malignancy? PLoS ONE 2014, 10, e111336. [Google Scholar] [CrossRef] [PubMed]
- Naba, A.; Clauser, K.R.; Lamar, J.M.; Carr, S.A.; Hynes, R.O. Extracellular matrix signatures of human mammary carcinoma identify novel metastasis promoters. Elife 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Shamsuddin, A.M.; Yang, G.Y.; Vucenik, I. Novel anti-cancer functions of IP6: Growth inhibition and differentiation of human mammary cancer cell lines in vitro. Anticancer Res. 1996, 16, 3287–3292. [Google Scholar] [PubMed]
- Tantivejkul, K.; Vucenik, I.; Shamsuddin, A.M. Inositol hexaphosphate (IP6) inhibits key events of cancer metastasis: I. In vitro studies of adhesion, migration and invasion of MDA-MB 231 human breast cancer cells. Anticancer Res. 2003, 23, 3671–3679. [Google Scholar] [PubMed]
- Zhang, J.; Gao, J.; Tan, X.; Wang, M.; Qin, R. Effects of down-regulation of integrin-beta1 expression on migration and hepatic metastasis of human colon carcinoma. J. Huazhong Univ. Sci. Technol. Med. Sci. 2010, 4, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Kolanus, W.; Seed, B. Integrins and inside-out signal transduction: Converging signals from PKC and PIP3. Curr. Opin. Cell Biol. 1997, 5, 725–731. [Google Scholar] [CrossRef]
- Ferry, S.; Matsuda, M.; Yoshida, H.; Hirata, M. Inositol hexakisphosphate blocks tumor cell growth by activating apoptotic machinery as well as by inhibiting the Akt/NFkappaB-mediated cell survival pathway. Carcinogenesis 2002, 12, 2031–2041. [Google Scholar] [CrossRef]
- Tantivejkul, K.; Vucenik, I.; Shamsuddin, A.M. Inositol hexaphosphate (IP6) inhibits key events of cancer metastasis: II. Effects on integrins and focal adhesions. Anticancer Res. 2003, 23, 3681–3689. [Google Scholar] [PubMed]
- Jensen, S.A.; Vainer, B.; Bartels, A.; Brunner, N.; Sorensen, J.B. Expression of matrix metalloproteinase 9 (MMP-9) and tissue inhibitor of metalloproteinases 1 (TIMP-1) by colorectal cancer cells and adjacent stroma cells—Associations with histopathology and patients outcome. Eur. J. Cancer 2010, 18, 3233–3242. [Google Scholar] [CrossRef] [PubMed]
- Ogata, Y.; Ookita, A.; Kakegawa, T. Significance of MMP-9 and TIMP-1 production during liver metastasis in colorectal cancer. Nihon Rinsho 1995, 7, 1811–1815. [Google Scholar]
- Thompson, L.U.; Zhang, L. Phytic acid and minerals: Effect on early markers of risk for mammary and colon carcinogenesis. Carcinogenesis 1991, 11, 2041–2045. [Google Scholar] [CrossRef]
- Kapral, M.; Wawszczyk, J.; Jurzak, M.; Dymitruk, D.; Weglarz, L. Evaluation of the expression of metalloproteinases 2 and 9 and their tissue inhibitors in colon cancer cells treated with phytic acid. Acta. Pol. Pharm. 2010, 6, 625–629. [Google Scholar]
- Bourboulia, D.; Stetler-Stevenson, W.G. Matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs): Positive and negative regulators in tumor cell adhesion. Semin. Cancer Biol. 2010, 3, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Vucenik, I.; Passaniti, A.; Vitolo, M.I.; Tantivejkul, K.; Eggleton, P.; Shamsuddin, A.M. Anti-angiogenic activity of inositol hexaphosphate (IP6). Carcinogenesis 2004, 11, 2115–2123. [Google Scholar] [CrossRef] [PubMed]
- Min, B.S.; Kim, N.K.; Jeong, H.C.; Chung, H.C. High levels of serum VEGF and TIMP-1 are correlated with colon cancer liver metastasis and intrahepatic recurrence after liver resection. Oncol. Lett. 2012, 1, 123–130. [Google Scholar]
- Sahni, A.; Francis, C.W. Vascular endothelial growth factor binds to fibrinogen and fibrin and stimulates endothelial cell proliferation. Blood 2000, 12, 3772–3778. [Google Scholar]
- Zittermann, S.I.; Issekutz, A.C. Basic fibroblast growth factor (bFGF, FGF-2) potentiates leukocyte recruitment to inflammation by enhancing endothelial adhesion molecule expression. Am. J. Pathol. 2006, 3, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Maehara, Y.; Kakeji, Y.; Kabashima, A.; Emi, Y.; Watanabe, A.; Akazawa, K.; Baba, H.; Kohnoe, S.; Sugimachi, K. Role of transforming growth factor-beta 1 in invasion and metastasis in gastric carcinoma. J. Clin. Oncol. 1999, 2, 607–614. [Google Scholar]
- Kapral, M.; Wawszczyk, J.; Hollek, A.; Weglarz, L. Induction of the expression of genes encoding TGF-beta isoforms and their receptors by inositol hexaphosphate in human colon cancer cells. Acta Pol. Pharm. 2013, 2, 357–363. [Google Scholar]
- Takahashi, T.; Ueno, H.; Shibuya, M. VEGF activates protein kinase C-dependent, but Ras-independent Raf-MEK-MAP kinase pathway for DNA synthesis in primary endothelial cells. Oncogene 1999, 13, 2221–2230. [Google Scholar] [CrossRef] [PubMed]
- Di, Y.; Zhang, Y.; Yang, H.; Wang, A.; Chen, X. The mechanism of CCN1-enhanced retinal neovascularization in oxygen-induced retinopathy through PI3K/Akt-VEGF signaling pathway. Drug Des. Dev. Ther. 2015, 9, 2463–2473. [Google Scholar]
- Bekhite, M.M.; Finkensieper, A.; Binas, S.; Muller, J.; Wetzker, R.; Figulla, H.R.; Sauer, H.; Wartenberg, M. VEGF-mediated PI3K class IA and PKC signaling in cardiomyogenesis and vasculogenesis of mouse embryonic stem cells. J. Cell Sci. 2011, 124, 1819–1830. [Google Scholar] [CrossRef] [PubMed]
- Marks, G.; Aydos, R.D.; Fagundes, D.J.; Pontes, E.R.; Takita, L.C.; Amaral, E.G.; Rossini, A.; Ynouye, C.M. Modulation of transforming growth factor beta2 (TGF-beta2) by inositol hexaphosphate in colon carcinogenesis in rats. Acta Cir. Bras. 2006, 21, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Cuschieri, J.; Maier, R.V. Mitogen-activated protein kinase (MAPK). Crit. Care Med. 2005, 12, S417–S419. [Google Scholar] [CrossRef]
- Bando, H.; Ikuno, Y.; Hori, Y.; Sayanagi, K.; Tano, Y. Mitogen-activated protein kinase (MAPK) and phosphatidylinositol-3 kinase (PI3K) pathways differently regulate retinal pigment epithelial cell-mediated collagen gel contraction. Exp. Eye Res. 2006, 3, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ji, Q.; Fan, Z.; Li, Q. Cellular signaling pathways implicated in metastasis of colorectal cancer and the associated targeted agents. Future Oncol. 2015, 21, 2911–2922. [Google Scholar] [CrossRef] [PubMed]
- Maffucci, T.; Piccolo, E.; Cumashi, A.; Iezzi, M.; Riley, A.M.; Saiardi, A.; Godage, H.Y.; Rossi, C.; Broggini, M.; Iacobelli, S.; et al. Inhibition of the phosphatidylinositol 3-kinase/Akt pathway by inositol pentakisphosphate results in antiangiogenic and antitumor effects. Cancer Res. 2005, 18, 8339–8349. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Ma, W.Y.; Hecht, S.S.; Dong, Z. Inositol hexaphosphate inhibits cell transformation and activator protein 1 activation by targeting phosphatidylinositol-3′ kinase. Cancer Res. 1997, 14, 2873–2878. [Google Scholar]
- Vucenik, I.; Ramakrishna, G.; Tantivejkul, K.; Anderson, L.M.; Ramljak, D. Inositol hexaphosphate (IP6) blocks proliferation of human breast cancer cells through a PKCdelta-dependent increase in p27Kip1 and decrease in retinoblastoma protein (pRb) phosphorylation. Breast Cancer Res. Treat. 2005, 1, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Vucenik, I.; Ramljak, D. The contradictory role of PKCdelta in cellular signaling. Breast Cancer Res. Treat. 2006, 1, 1–2. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Product Size (bp) |
|---|---|---|
| β-actin | F 5′-TTCCTTCTTGGGTATGGAAT-3′ | 174 |
| R 5′-GAGCAATGATCTTGATCTTC-3′ | ||
| Collagen IV | F 5′-CCTCCAGGTTTCCCTACTCC-3′ | 123 |
| R 5′-GGCTCCATCTCTTCCACTTG-3′ | ||
| Laminin | F 5′-AGTACCAGGAGGAAGCAGCA-3′ | 126 |
| R 5′-TCTAAGCATCGCAAGGGAGT-3′ | ||
| Fibronectin | F 5′-ACCACCCAGAACTACGATGC-3′ | 124 |
| R 5′-CATGCTGCTTATCCCACTGA-3′ | ||
| Integrin-β1 | F 5′-CAGTGAATGGCAACAATGAAG-3′ | 133 |
| R 5′-ATCAGCAGCAAGGCAAGG-3′ | ||
| Matrix metalloproteinase 9 | F 5′-TCTACTGGGCGTTAGGGACA-3′ | 112 |
| R 5′-TCGGGAGAGAGAGGAGTCTG-3 | ||
| VEGF | F 5′-CCCTTCGTCCTCTCCTTACC-3′ | 118 |
| R 5′-AAGCCACTCACACACACAGC-3′ | ||
| bFGF | F 5′-CGGTCACGGAAATACTCC-3′ | 120 |
| R 5′-GCTCTTAGCAGACATTGGAAG-3′ | ||
| TGF-β | F 5′-ATTCCTGGCGTTACCTTGG-3′ | 120 |
| R 5′-AGCCCTGTATTCCGTCTCCT-3′ |
| Group | Number | Tumor Weight (g) | Inhibition Rate (%) |
|---|---|---|---|
| IP6 | 9 | 0.541 ± 0.349 *,# | 53.52 |
| Ins | 9 | 0.557 ± 0.421 *,# | 52.15 |
| IP6 + Ins | 9 | 0.319 ± 0.117 # | 72.59 |
| Normal saline | 11 | 1.164 ± 0.558 | - |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, M.; Song, Y.; Wen, Z.; Lu, X.; Cui, L. Inositol Hexaphosphate and Inositol Inhibit Colorectal Cancer Metastasis to the Liver in BALB/c Mice. Nutrients 2016, 8, 286. https://doi.org/10.3390/nu8050286
Fu M, Song Y, Wen Z, Lu X, Cui L. Inositol Hexaphosphate and Inositol Inhibit Colorectal Cancer Metastasis to the Liver in BALB/c Mice. Nutrients. 2016; 8(5):286. https://doi.org/10.3390/nu8050286
Chicago/Turabian StyleFu, Min, Yang Song, Zhaoxia Wen, Xingyi Lu, and Lianhua Cui. 2016. "Inositol Hexaphosphate and Inositol Inhibit Colorectal Cancer Metastasis to the Liver in BALB/c Mice" Nutrients 8, no. 5: 286. https://doi.org/10.3390/nu8050286
APA StyleFu, M., Song, Y., Wen, Z., Lu, X., & Cui, L. (2016). Inositol Hexaphosphate and Inositol Inhibit Colorectal Cancer Metastasis to the Liver in BALB/c Mice. Nutrients, 8(5), 286. https://doi.org/10.3390/nu8050286