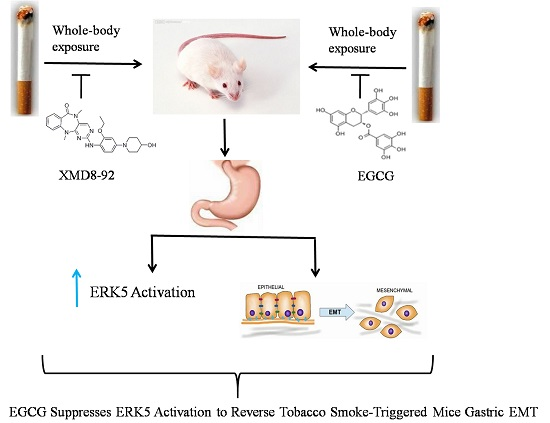
EGCG Suppresses ERK5 Activation to Reverse Tobacco Smoke-Triggered Gastric Epithelial-Mesenchymal Transition in BALB/c Mice

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Section
2.1. Chemicals and Reagents
2.2. Mice and Tobacco Smoke Exposure
2.3. In Vivo Delivery of Specific ERK5 Inhibitor
2.4. EGCG Treatment of Mice
2.5. Western Blot Analysis
2.6. Quantitative Real-Time PCR
2.7. Immunohistochemistry
2.8. Statistical Analysis
3. Results
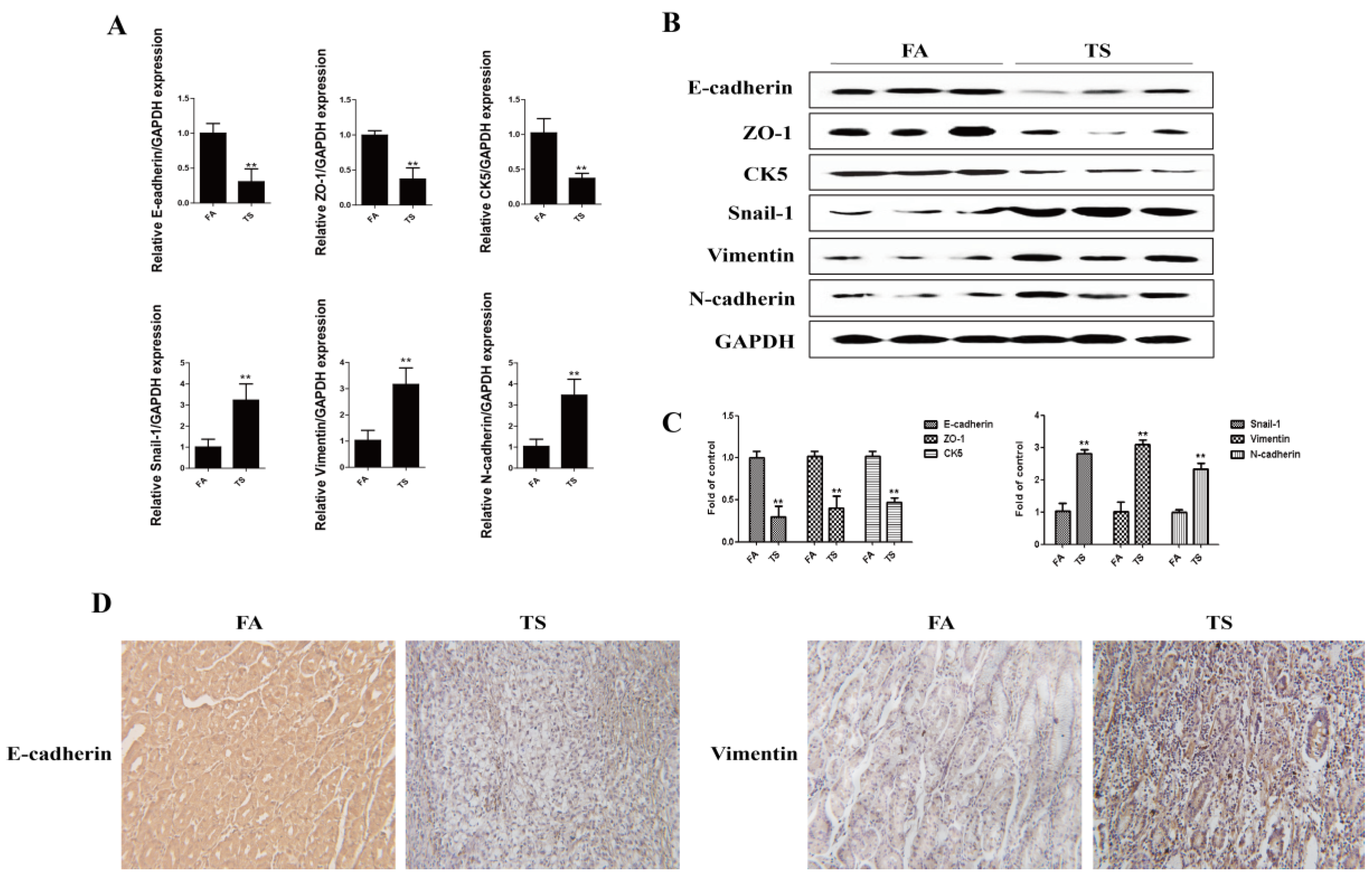
3.1. Tobacco Smoke Induced Epithelial-Mesenchymal Transition Changes in Gastric Tissues of Mice
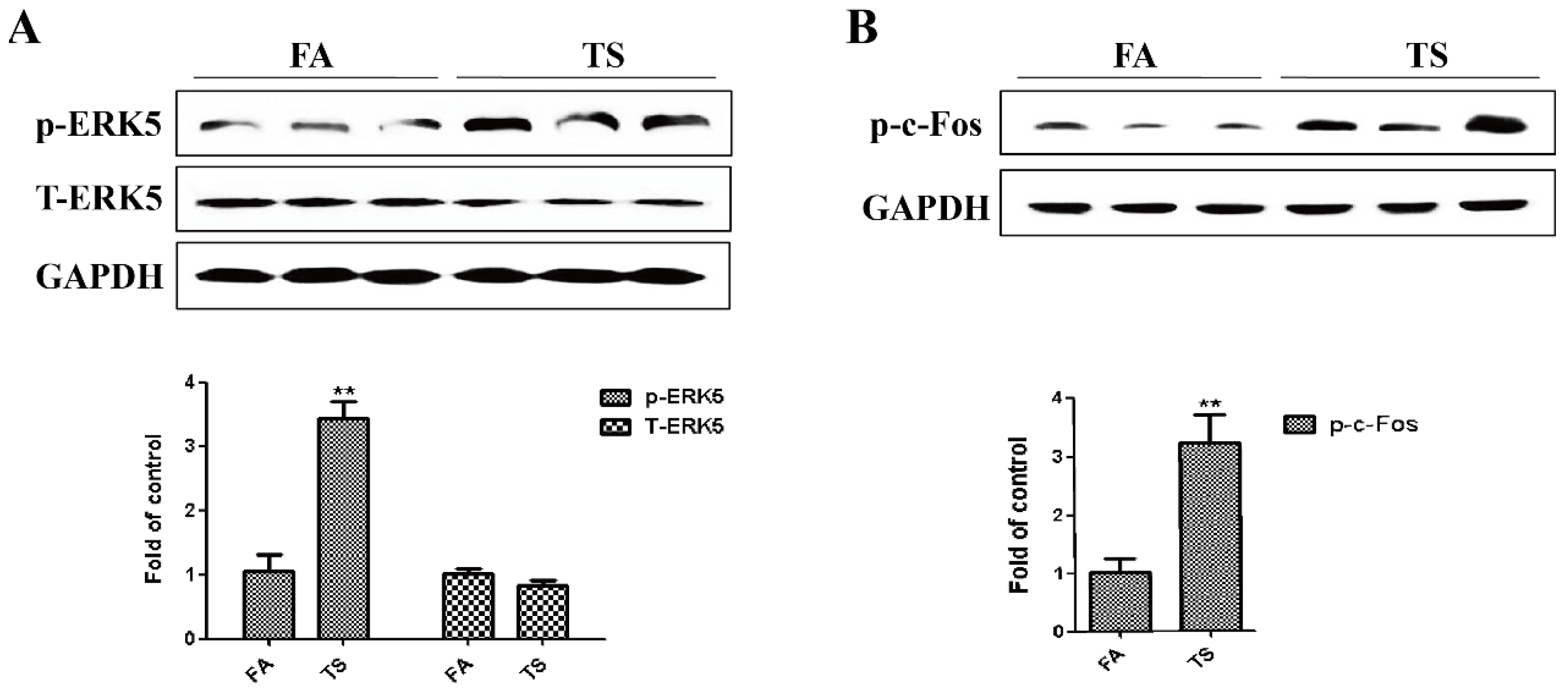
3.2. Tobacco Smoke-Induced Gastric Epithelial-Mesenchymal Transition Was Associated with ERK5 Activation
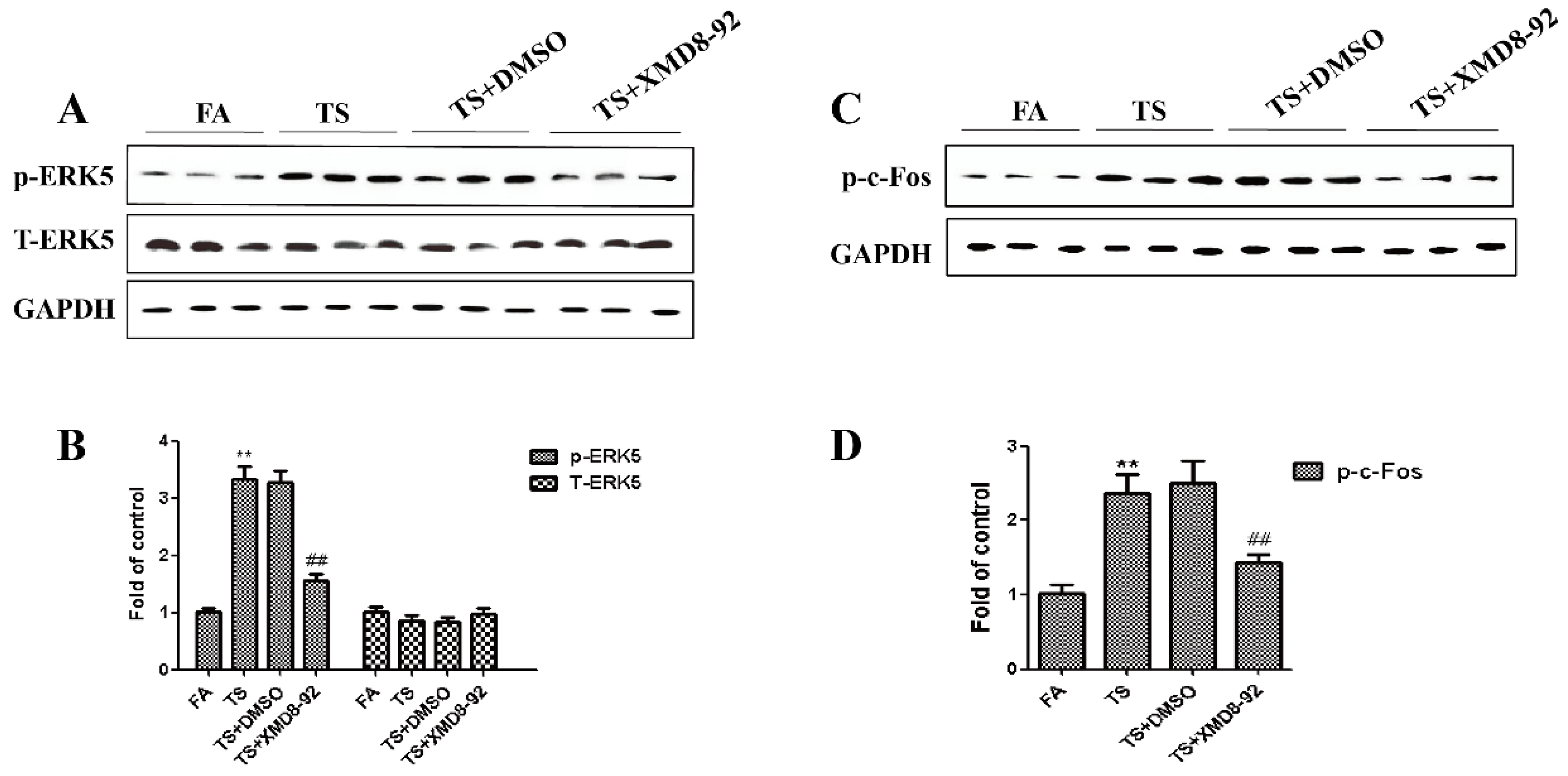
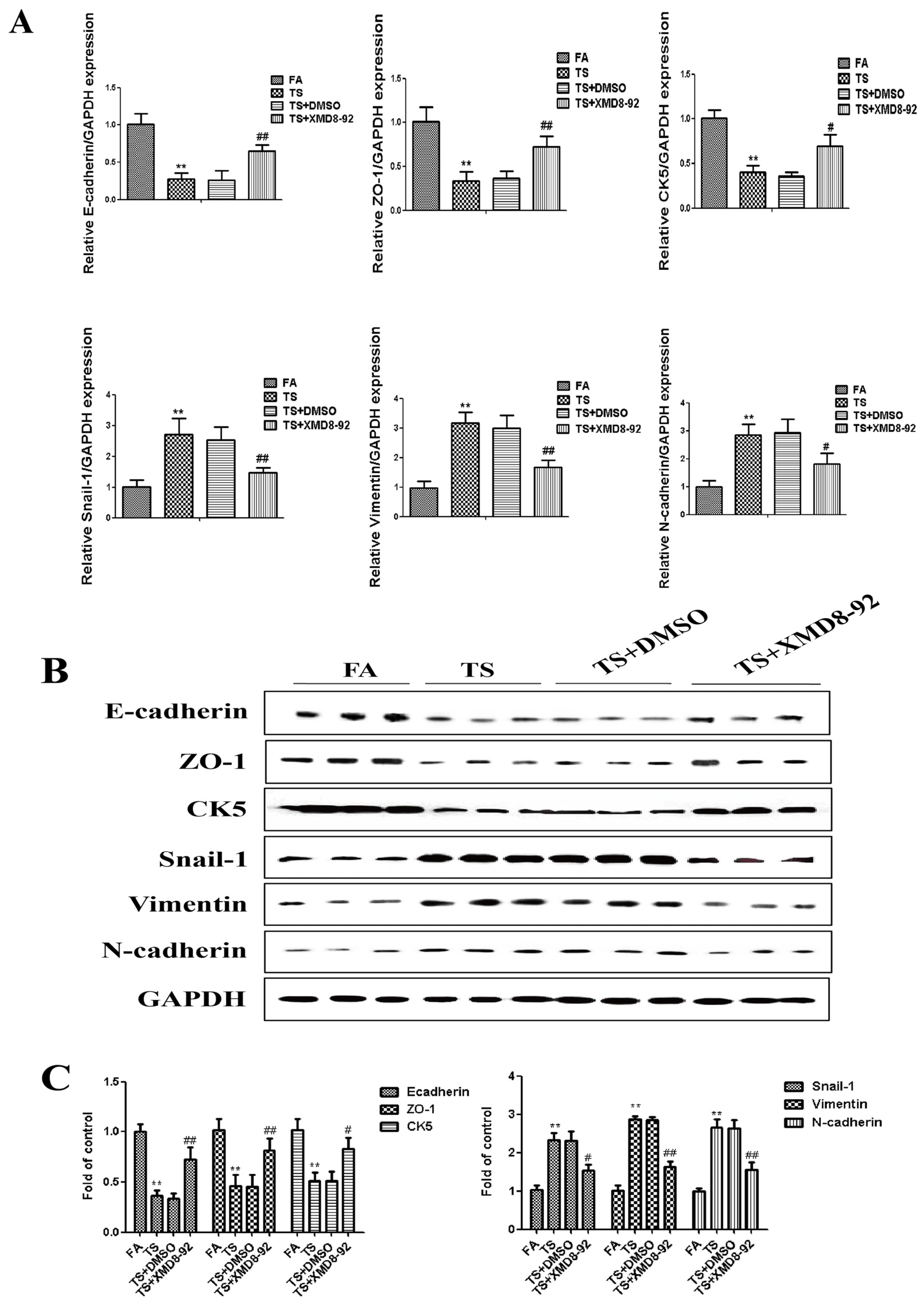
3.3. ERK5 Suppression Reversed Tobacco Smoke-Triggered Gastric Epithelial-Mesenchymal Transition
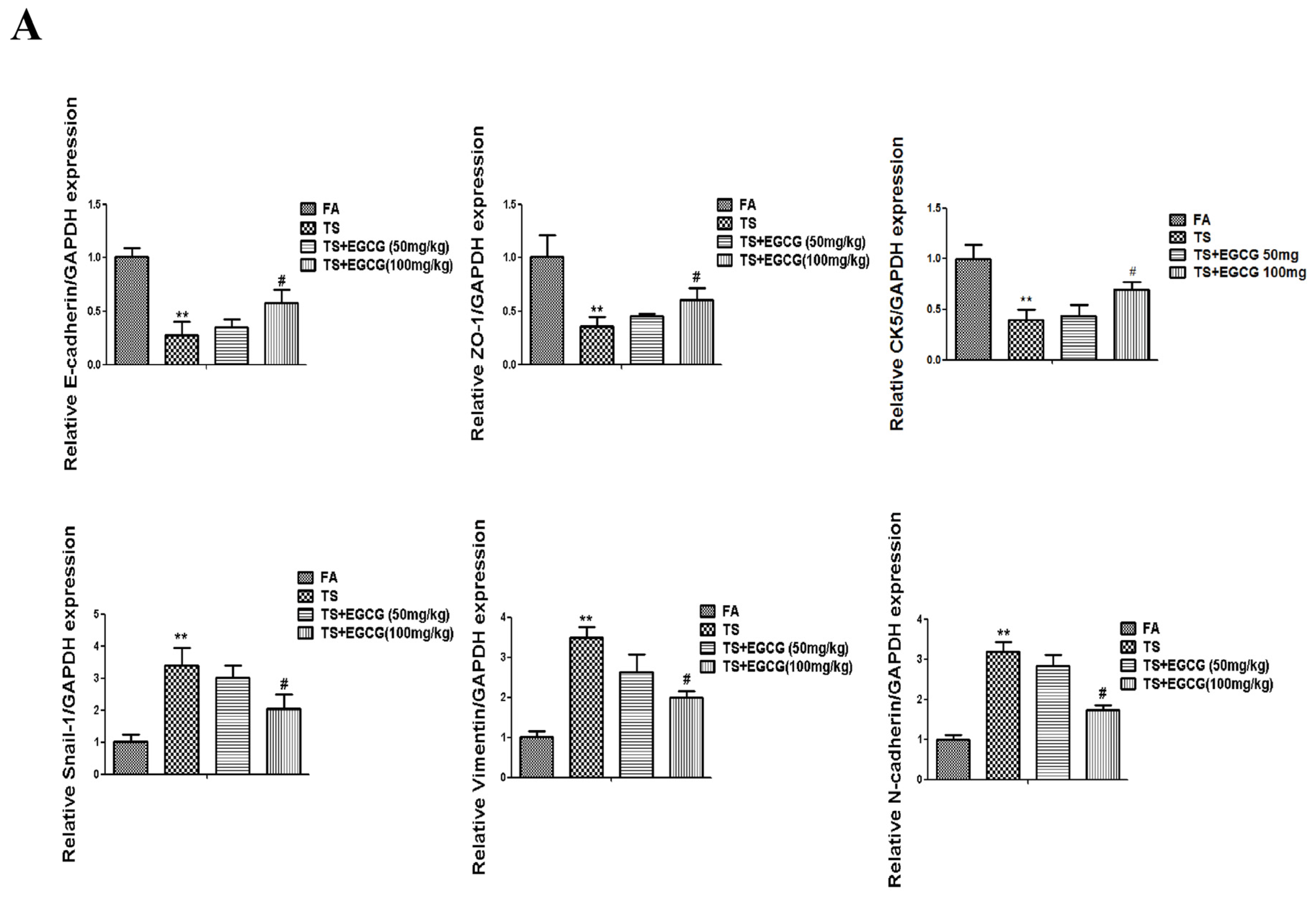
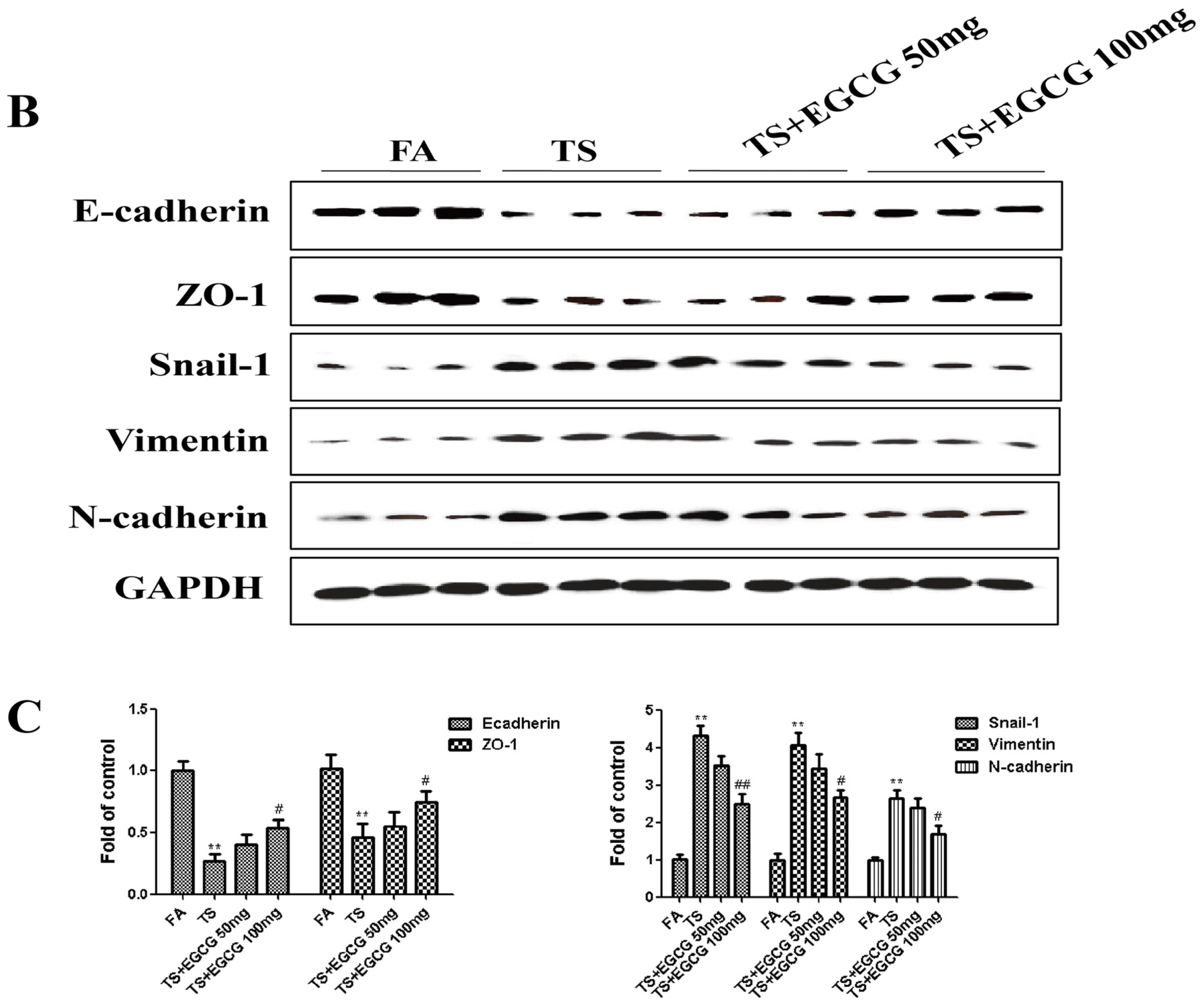
3.4. EGCG Attenuated Tobacco Smoke- Induced Gastric Epithelial-Mesenchymal Transition in the Stomach of Mice
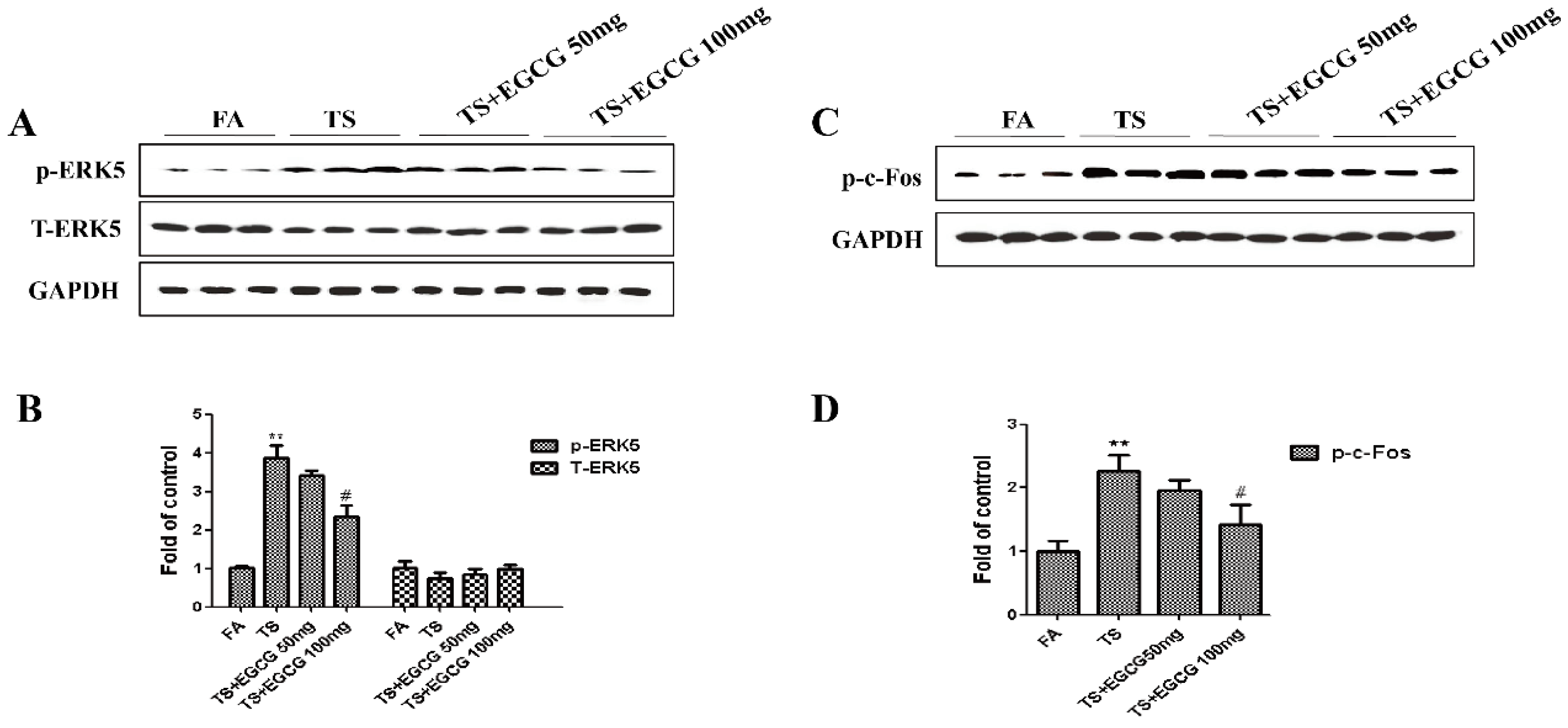
3.5. EGCG Reversed Tobacco Smoke-Induced ERK5 Activation in the Stomach of Mice
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mahipal, A.; Choi, M.; Kim, R. Second-line treatment of advanced gastric cancer: Where do we stand? J. Natl. Compr. Canc. Netw. 2015, 13, 1281–1289. [Google Scholar] [PubMed]
- Qiu, L.X.; He, J.; Cheng, L.; Zhou, F.; Wang, M.Y.; Sun, M.H.; Zhou, X.Y.; Li, J.; Guo, W.J.; Wang, Y.N.; et al. Genetic variant of PRKAA1 and gastric cancer risk in an Eastern Chinese population. Oncotarget 2015, 6, 42661–42666. [Google Scholar] [PubMed]
- Chen, W.; Zheng, R.; Zhang, S.; Zhao, P.; Zeng, H.; Zou, X. Report of cancer incidence and mortality in China, 2010. Ann. Transl. Med. 2014, 2, 61. [Google Scholar] [PubMed]
- Zhu, X.; Li, J. Gastric carcinoma in China: Current status and future perspectives (Review). Oncol. Lett. 2010, 1, 407–412. [Google Scholar] [PubMed]
- Chen, Y.S.; Chen, J.G.; Zhu, J.; Zhang, Y.H.; Ding, L.L. Long-term survival trends of gastric cancer patients between 1972 and 2011 in Qidong. Chin. J. Cancer 2015, 34, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.F.; Wu, R.; Xie, W.; Geng, H.; Zhao, L.; Xie, C.F.; Wu, J.S.; Geng, S.S.; Li, X.T.; Zhu, M.M.; et al. Curcumin suppresses MAPK pathways to reverse tobacco smoke-induced gastric epithelial-mesenchymal transition in mice. Phytother. Res. 2015, 29, 1665–1671. [Google Scholar] [CrossRef] [PubMed]
- Malakar, M.; Devi, K.R.; Phukan, R.K.; Kaur, T.; Deka, M.; Puia, L.; Baruah, D.; Mahanta, J.; Narain, K. CYP2E1 genetic polymorphism with dietary, tobacco, alcohol habits, H. pylori infection status and susceptibility to stomach cancer in Mizoram, India. Asian. Pac. J. Cancer Prev. 2014, 15, 8815–8822. [Google Scholar] [CrossRef] [PubMed]
- Li, L.F.; Chan, R.L.; Lu, L.; Shen, J.; Zhang, L.; Wu, W.K.; Wang, L.; Hu, T.; Li, M.X.; Cho, C.H. Cigarette smoking and gastrointestinal diseases: The causal relationship and underlying molecular mechanisms (review). Int. J. Mol. Med. 2014, 34, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Shin, V.Y.; Jin, H.; Ng, E.K.; Cheng, A.S.; Chong, W.W.; Wong, C.Y.; Leung, W.K.; Sung, J.J.; Chu, K.M. NF-kappaβ targets miR-16 and miR-21 in gastric cancer: Involvement of prostaglandin E receptors. Carcinogenesis 2011, 32, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Phukan, R.K.; Zomawia, E.; Narain, K.; Hazarika, N.C.; Mahanta, J. Tobacco use and stomach cancer in Mizoram, India. Cancer Epidemiol. Biomarkers Prev. 2005, 14, 1892–1896. [Google Scholar] [CrossRef] [PubMed]
- Babhadiashar, N.; Sotoudeh, M.; Azizi, E.; Bashiri, J.; Didevar, R.; Malekzadeh, R.; Ghahremani, M.H. Correlation between cigarette smoking and urine cotinine level in gastric cancer patients. Iran. J. Pharm. Res. 2014, 13, 313–318. [Google Scholar] [PubMed]
- Sjodahl, K.; Lu, K.; Nilsen, T.I.; Ye, W.; Hveem, K.; Vatten, L.; Lagergren, J. Smoking and alcohol drinking in relation to risk of gastric cancer: A population-based, prospective cohort study. Int. J. Cancer 2007, 120, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ren, J.W.; Wong, C.C.; Wu, W.K.; Ren, S.X.; Shen, J.; Chan, R.L.; Cho, C.H. Effects of cigarette smoke and its active components on ulcer formation and healing in the gastrointestinal mucosa. Curr. Med. Chem. 2012, 19, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Dong, D.; Sun, L.; Zhang, G.; Sun, L. Prognostic significance of the epithelial-to-mesenchymal transition markers e-cadherin, vimentin and twist in bladder cancer. Int. Braz. J. Urol. 2014, 40, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hu, G.; Chen, D.; Gong, A.Y.; Soori, G.S.; Dobleman, T.J.; Chen, X.M. Suppression of SCARA5 by snail1 is essential for EMT-associated cell migration of A549 cells. Oncogenesis 2013, 2, e73. [Google Scholar] [CrossRef] [PubMed]
- Shin, V.Y.; Jin, H.C.; Ng, E.K.; Sung, J.J.; Chu, K.M.; Cho, C.H. Activation of 5-lipoxygenase is required for nicotine mediated epithelial-mesenchymal transition and tumor cell growth. Cancer Lett. 2010, 292, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gallup, M.; Zlock, L.; Basbaum, C.; Finkbeiner, W.E.; McNamara, N.A. Cigarette smoke disrupts the integrity of airway adherens junctions through the aberrant interaction of p120-catenin with the cytoplasmic tail of MUC1. J. Pathol. 2013, 229, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, H.; Borok, Z.; Davies, K.J.; Ursini, F.; Forman, H.J. Cigarette smoke extract stimulates epithelial-mesenchymal transition through Src activation. Free Radic. Biol. Med. 2012, 52, 1437–1442. [Google Scholar] [CrossRef] [PubMed]
- Drew, B.A.; Burow, M.E.; Beckman, B.S. MEK5/ERK5 pathway: The first fifteen years. Biochim. Biophys. Acta 2012, 1825, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, S.; Nishida, E. MAPK signalling: ERK5 versus ERK1/2. EMBO Rep. 2006, 7, 782–786. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Fearns, C.; Eliceiri, B.; Yang, Y.; Lee, J.D. Big mitogen-activated protein kinase 1/extracellular signal-regulated kinase 5 signaling pathway is essential for tumor-associated angiogenesis. Cancer Res. 2005, 65, 7699–7706. [Google Scholar] [PubMed]
- Wang, X.; Tournier, C. Regulation of cellular functions by the ERK5 signalling pathway. Cell Signal. 2006, 18, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Nitschke, A.M.; Xiong, W.; Zhang, Q.; Tang, Y.; Bloch, M.; Elliott, S.; Zhu, Y.; Bazzone, L.; Yu, D.; et al. Proteomic analysis of tumor necrosis factor-alpha resistant human breast cancer cells reveals a MEK5/Erk5-mediated epithelial-mesenchymal transition phenotype. Breast. Cancer Res. 2008, 10, R105. [Google Scholar] [CrossRef] [PubMed]
- Antoon, J.W.; Martin, E.C.; Lai, R.; Salvo, V.A.; Tang, Y.; Nitzchke, A.M.; Elliott, S.; Nam, S.Y.; Xiong, W.; Rhodes, L.V.; et al. MEK5/ERK5 signaling suppresses estrogen receptor expression and promotes hormone-independent tumorigenesis. PLoS ONE 2013, 8, e69291. [Google Scholar] [CrossRef] [PubMed]
- Madak-Erdogan, Z.; Ventrella, R.; Petry, L.; Katzenellenbogen, B.S. Novel roles for ERK5 and cofilin as critical mediators linking ERalpha-driven transcription, actin reorganization, and invasiveness in breast cancer. Mol. Cancer Res. 2014, 12, 714–727. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Wu, Y.; Wehrli, B.; Chakrabarti, S.; Chakraborty, C. Modulation of ERK5 is a novel mechanism by which Cdc42 regulates migration of breast cancer cells. J. Cell. Biochem. 2015, 116, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Woo, C.H.; Massett, M.P.; Shishido, T.; Itoh, S.; Ding, B.; McClain, C.; Che, W.; Vulapalli, S.R.; Yan, C.; Abe, J. ERK5 activation inhibits inflammatory responses via peroxisome proliferator-activated receptor delta (PPARdelta) stimulation. J. Biol. Chem. 2006, 281, 32164–32174. [Google Scholar] [CrossRef] [PubMed]
- Cerezo-Guisado, M.I.; Zur, R.; Lorenzo, M.J.; Itoh, S.; Ding, B.; McClain, C.; Che, W.; Vulapalli, S.R.; Yan, C.; Abe, J. Implication of Akt, ERK1/2 and alternative p38MAPK signalling pathways in human colon cancer cell apoptosis induced by green tea EGCG. Food Chem. Toxicol. 2015, 84, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Wang, W. Green tea polyphenol EGCG suppresses osteosarcoma cell growth through upregulating miR-1. Tumour. Biol. 2016, 37, 4373–4382. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.O.; Chan, K.P.; Yang, Y.P.; Qin, Y.J.; Li, W.Y.; Chan, S.O.; Wang, C.C.; Pang, C.P. Effects of EGCG content in green tea extract on pharmacokinetics, oxidative status and expression of inflammatory and apoptotic genes in the rat ocular tissues. J. Nutr. Biochem. 2015, 26, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, A.; Sarkar, J.; Chakraborti, T.; Chakraborti, S. Role of Spm-Cer-S1P signalling pathway in MMP-2 mediated U46619-induced proliferation of pulmonary artery smooth muscle cells: Protective role of epigallocatechin-3-gallate. Cell Biochem. Funct. 2015, 33, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.H.; Shabrmi, F.M.A.; Allemailem, K.S.; Aly, S.M.; Khan, M.A. Implications of green tea and its constituents in the prevention of cancer via the modulation of cell signalling pathway. Biomed. Res. Int. 2015, 2015, 925640. [Google Scholar] [CrossRef] [PubMed]
- Pi, X.; Garin, G.; Xie, L.; Zheng, Q.; Wei, H.; Abe, J.; Yan, C.; Berk, B.C. BMK1/ERK5 is a novel regulator of angiogenesis by destabilizing hypoxia inducible factor 1alpha. Circ. Res. 2005, 96, 1145–1151. [Google Scholar] [CrossRef] [PubMed]
- Rovida, E.; Navari, N.; Caligiuri, A.; DelloSbarba, P.; Marra, F. ERK5 differentially regulates PDGF-induced proliferation and migration of hepatic stellate cells. J. Hepatol. 2008, 48, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Spiering, D.; Schmolke, M.; Ohnesorge, N.; Schmidt, M.; Goebeler, M.; Wegener, J.; Wixler, V.; Ludwig, S. MEK5/ERK5 signaling modulates endothelial cell migration and focal contact turnover. J. Biol. Chem. 2009, 284, 24972–24980. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Xie, W.; Wu, R.; Geng, H.; Zhao, L.; Xie, C.; Li, X.; Huang, C.; Zhu, J.; Zhu, M.; et al. ERK5 negatively regulates tobacco smoke-induced pulmonary epithelial-mesenchymal transition. Oncotarget 2015, 6, 19605–19618. [Google Scholar] [CrossRef] [PubMed]
- Takagaki, A.; Nanjo, F. Effects of metabolites produced from (−)-epigallocatechingallate by rat intestinal bacteria on angiotensin i-converting enzyme activity and blood pressure in spontaneously hypertensive rats. J. Agric. Food Chem. 2015, 63, 8262–8266. [Google Scholar] [CrossRef] [PubMed]
- Rietveld, A.; Wiseman, S. Antioxidant effects of tea: Evidence from human clinical trials. J. Nutr. 2003, 133, 3285S–3292S. [Google Scholar] [PubMed]
- Jin, X.; Liu, M.; Chen, Z.; Mao, R.; Xiao, Q.; Gao, H.; Wei, M. Separation and purification of epigallocatechin-3-gallate (EGCG) from green tea using combined macroporous resin and polyamide column chromatography. J. Chromatogr. B 2015, 1002, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Forester, S.C.; Lambert, J.D. The catechol-O-methyltransferase inhibitor, tolcapone, increases the bioavailability of unmethylated (−)-epigallocatechin-3-gallate in mice. J. Funct. Foods 2015, 17, 183–188. [Google Scholar] [CrossRef] [PubMed]







© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, L.; Chen, J.; Tang, H.; Bai, L.; Lu, C.; Wang, K.; Li, M.; Yan, Y.; Tang, L.; Wu, R.; et al. EGCG Suppresses ERK5 Activation to Reverse Tobacco Smoke-Triggered Gastric Epithelial-Mesenchymal Transition in BALB/c Mice. Nutrients 2016, 8, 380. https://doi.org/10.3390/nu8070380
Lu L, Chen J, Tang H, Bai L, Lu C, Wang K, Li M, Yan Y, Tang L, Wu R, et al. EGCG Suppresses ERK5 Activation to Reverse Tobacco Smoke-Triggered Gastric Epithelial-Mesenchymal Transition in BALB/c Mice. Nutrients. 2016; 8(7):380. https://doi.org/10.3390/nu8070380
Chicago/Turabian StyleLu, Ling, Jia Chen, Hua Tang, Ling Bai, Chun Lu, Kehuan Wang, Manli Li, Yinmei Yan, Ling Tang, Rui Wu, and et al. 2016. "EGCG Suppresses ERK5 Activation to Reverse Tobacco Smoke-Triggered Gastric Epithelial-Mesenchymal Transition in BALB/c Mice" Nutrients 8, no. 7: 380. https://doi.org/10.3390/nu8070380
APA StyleLu, L., Chen, J., Tang, H., Bai, L., Lu, C., Wang, K., Li, M., Yan, Y., Tang, L., Wu, R., Ye, Y., Jin, L., & Liang, Z. (2016). EGCG Suppresses ERK5 Activation to Reverse Tobacco Smoke-Triggered Gastric Epithelial-Mesenchymal Transition in BALB/c Mice. Nutrients, 8(7), 380. https://doi.org/10.3390/nu8070380