High Dietary Fructose Intake on Cardiovascular Disease Related Parameters in Growing Rats



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Diet
2.2. Body Weight, Food Consumption, and Fat Mass
2.3. Analysis of Blood Parameters
2.4. Analysis of Abdominal Aorta
2.5. Quantitative Real-Time Polymerase Chain Reaction (PCR)
2.6. Statistical Analysis
3. Results
3.1. Body Weights, Food Intakes, Calorie Intakes, and Food Efficiency Ratios
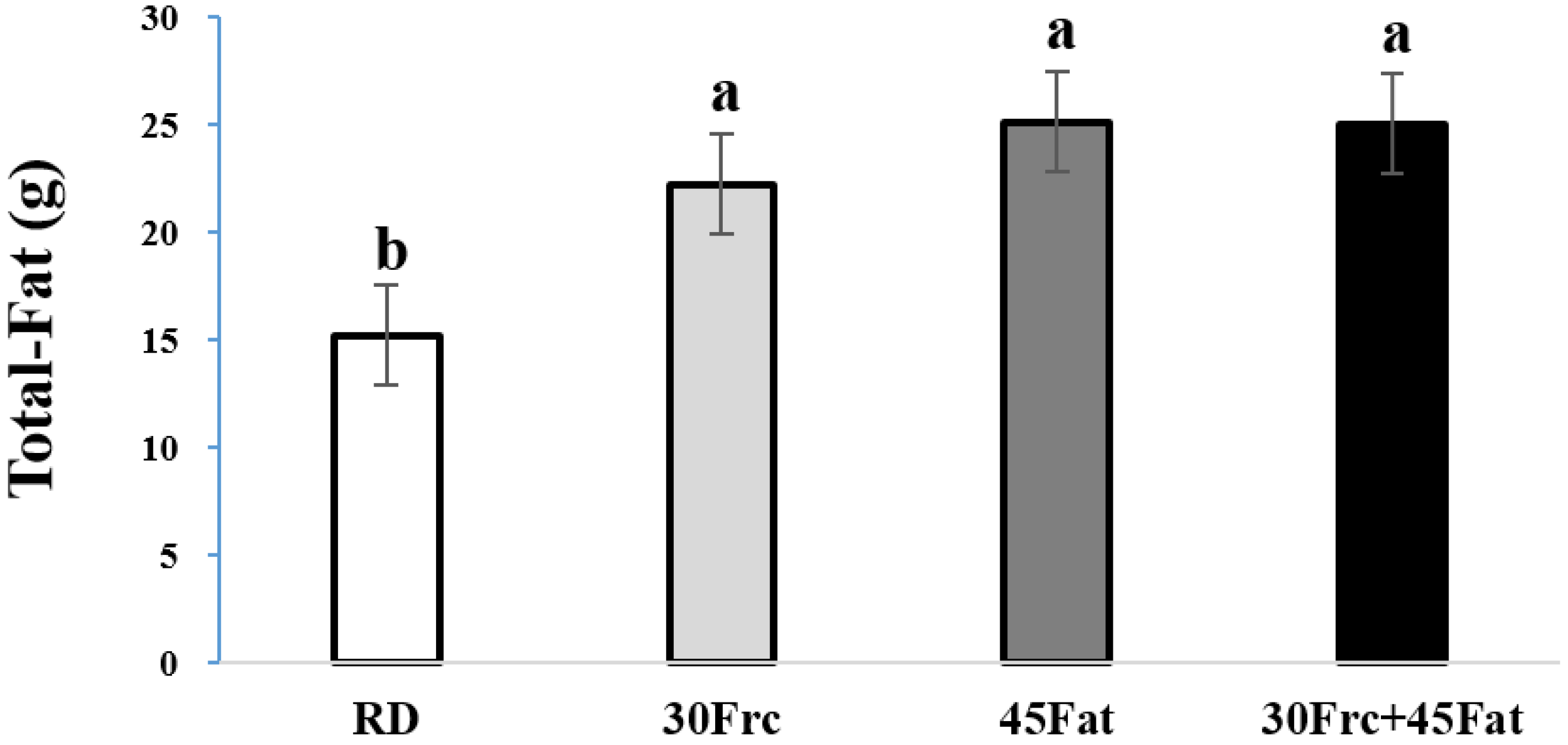
3.2. Total-Fat Weights
3.3. Serum Levels of Glucose and Insulin, HOMA-IR, and QUICKI
3.4. Serum Levels of Lipid Profiles
3.5. Serum Levels of Pro-Inflammatory Cytokines
3.6. Analysis of Abdominal Aorta Wall Thickness and Lumen Diameter
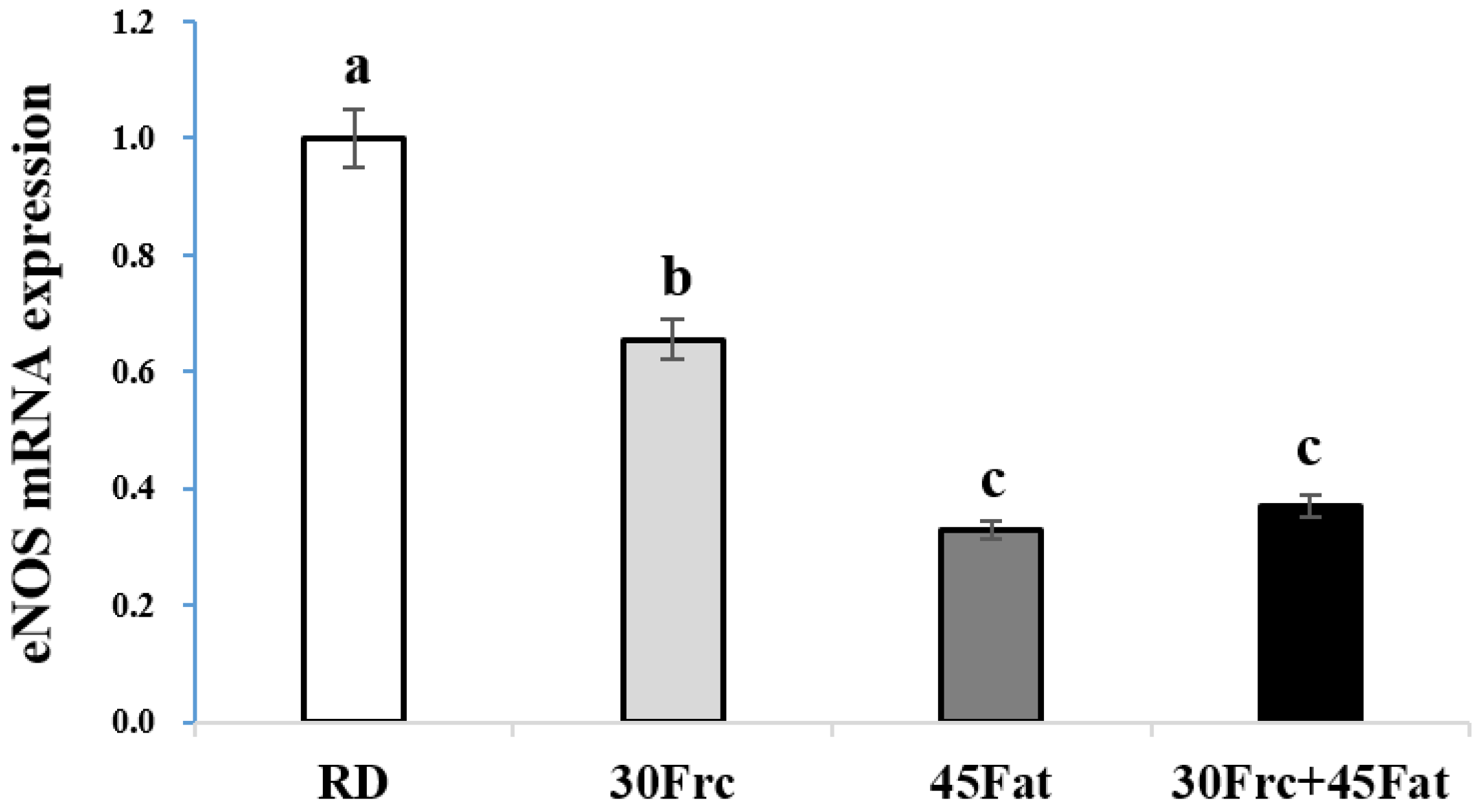
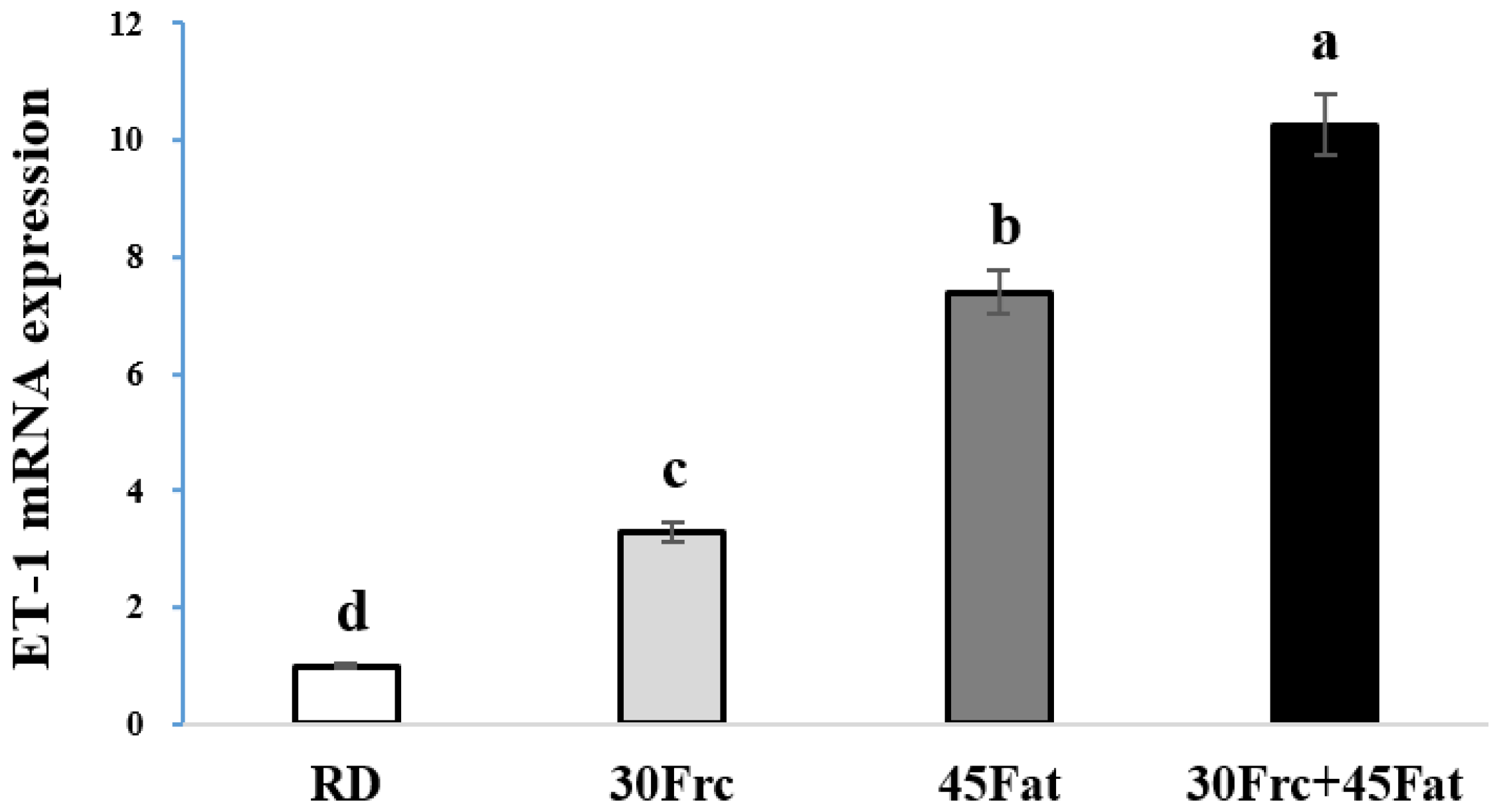
3.7. Abdominal Aortic eNOS and ET-1 mRNA Expression Measured by qRTPC
4. Discussion
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Kim, S.D.; Moon, H.; Park, J.S.; Lee, Y.C.; Shin, G.Y.; Jo, H.B.; Kim, B.S.; Kim, J.H.; Chae, Y.Z. Macromineral intake in non-alcoholic beverages for children and adolescents: Using the fourth Korea national health and nutrition examination survey (KNHANES IV, 2007–2009). Korean J. Nutr. 2013, 46, 50–60. [Google Scholar] [CrossRef]
- The Korea National Health and Nutrition Examination Survey (KNHNES). Dietary Intake Survey of Infant, Children and Adolescents; The Korea National Health and Nutrition Examination Survey: Cheongju, Korea, 2011.
- Korea Health Industry Development Institute (KHIDI). In-Depth Analysis on the Dietary Intake Survey of Infant, Children and Adolescents (II); Korea Health Industry Development Institute: Cheongju, Korea, 2010. [Google Scholar]
- Bandini, L.G.; Vu, D.; Must, A.; Cyr, H.; Goldberg, A.; Dietz, W.H. Comparison of high-calorie, low-nutrient-dense food consumption among obese and non-obese adolescents. Obes. Res. 1999, 7, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Berkey, C.S.; Rockett, H.R.; Field, A.E.; Gillman, M.W.; Colditz, G.A. Sugar-added beverages and adolescent weight change. Obes. Res. 2004, 12, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Hu, H.; Zharikov, S.; Tuttle, K.R.; Short, R.A.; Glushakova, O.; Ouyang, X.; Feig, D.I.; Block, E.R.; Herrera-Acosta, J.; et al. A causal role for uric acid in fructose-induced metabolic syndrome. Am. J. Physiol. Ren. Physiol. 2006, 290, F625–F631. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.S.; Nallasamy, P.; Muniyandi, P.; Periyasami, V.; Carani, V.A. Genistein improves liver function and attenuates non-alcoholic fatty liver disease in a rat model of insulin resistance. J. Diabetes 2009, 1, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Shih, C.; Lin, C.; Lin, W.; Wu, J. Momordica charantia extract on insulin resistance and the skeletal muscle GLUT4 protein in fructose-fed rats. J. Ethnopharmacol. 2009, 123, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Bocarsly, M.E.; Powell, E.S.; Avena, N.M.; Hoebel, B.G. High-fructose corn syrup causes characteristics of obesity in rats: Increased body weight, body fat and triglyceride levels. Pharmacol. Biochem. Behav. 2010, 97, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Crescenzo, R.; Bianco, F.; Coppola, P.; Mazzoli, A.; Valiante, S.; Liverini, G.; Iossa, S. Adipose tissue remodeling in rats exhibiting fructose-induced obesity. Eur. J. Nutr. 2014, 53, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muniyappa, R.; Montagnani, M.; Koh, K.K.; Quon, M.J. Cardiovascular actions of insulin. Endocr. Rev. 2007, 28, 463–491. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Arikawa, E.; Galipeau, D.; Battell, M.; McNeill, J.H. Androgens are necessary for the development of fructose-induced hypertension. Hypertension 2004, 43, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Behr-Roussel, D.; Oudot, A.; Compagnie, S.; Gorny, D.; Le Coz, O.; Bernabe, J.; Wayman, C.; Alexandre, L.; Giuliano, F. Impact of a long-term sildenafil treatment on pressor response in conscious rats with insulin resistance and hypertriglyceridemia. Am. J. Hypertens. 2008, 21, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Endemann, D.H.; Schiffrin, E.L. Endothelial dysfunction. J. Am. Soc. Nephrol. 2004, 15, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Austin, M.A.; King, M.C.; Vranizan, K.M.; Krauss, R.M. Atherogenic lipoprotein phenotype: A proposed genetic marker for coronary heart disease risk. Circulation 1990, 82, 495–506. [Google Scholar] [CrossRef] [PubMed]
- De Castro, U.G.; dos Santos, R.A.; Silva, M.E.; de Lima, W.G.; Campagnole-Santos, M.J.; Alzamora, A.C. Age-dependent effect of high-fructose and high-fat diets on lipid metabolism and lipid accumulation in liver and kidney of rats. Lipids Health Dis. 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kadowaki, T. Physiological and pathophysiological roles of adiponectin and adiponectin receptors in the integrated regulation of metabolic and cardiovascular diseases. Int. J. Obes. 2008, 32, S13–S18. [Google Scholar] [CrossRef] [PubMed]
- Ross, R. The pathogenesis of atherosclerosis: A perspective for the 1990s. Nature 1993, 362, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Sinitskaya, N.; Gourmelen, S.; Schuster-Klein, C.; Guardiola-Lemaitre, B.; Pevet, P.; Challet, E. Increasing the fat-to-carbohydrate ratio in a high-fat diet prevents the development of obesity but not a prediabetic state in rats. Clin. Sci. 2007, 113, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Matveyenko, A.V.; Gurlo, T.; Daval, M.; Butler, A.E.; Butler, P.C. Successful versus failed adaptation to high-fat diet-induced insulin resistance: The role of IAPP-induced beta-cell endoplasmic reticulum stress. Diabetes 2009, 58, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Ouguerram, K.; Nguyen, P. Lipid profile and insulin sensitivity in rats fed with high-fat or high-fructose diets. Br. J. Nutr. 2011, 106, S206–S210. [Google Scholar]
- Tran, L.T.; Yuen, V.G.; McNeill, J.H. The fructose-fed rat: A review on the mechanisms of fructose-induced insulin resistance and hypertension. Mol. Cell. Biochem. 2009, 332, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.; Adeli, K. Dietary fructose and the metabolic syndrome. Curr. Opin. Gastroenterol. 2008, 24, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulos, T.J.; Lowndes, J.; Zukley, L.; Melanson, K.J.; Nguyen, V.; Huffman, A.; Rippe, J.M. The effect of high-fructose corn syrup consumption on triglycerides and uric acid. J. Nutr. 2009, 139, 1242S–1245S. [Google Scholar] [CrossRef] [PubMed]
- Shulman, G.I. Cellular mechanisms of insulin resistance. J. Clin. Investig. 2000, 106, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Blakely, S.R.; Hallfrisch, J.; Reiser, S.; Prather, E.S. Long-term effects of moderate fructose feeding on glucose tolerance parameters in rats. J. Nutr. 1981, 111, 307–314. [Google Scholar] [PubMed]
- Tappy, L.; Le, K.A. Metabolic effects of fructose and the worldwide increase in obesity. Physiol. Rev. 2010, 90, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Basciano, H.; Federico, L.; Adeli, K. Fructose, insulin resistance, and metabolic dyslipidemia. Nutr. Metab. 2005, 2, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iida, T.; Yamada, T.; Hayashi, N.; Okuma, K.; Izumori, K.; Ishii, R.; Matsuo, T. Reduction of abdominal fat accumulation in rats by 8-week ingestion of a newly developed sweetener made from high fructose corn syrup. Food Chem. 2013, 138, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Catena, C.; Giacchetti, G.; Novello, M.; Colussi, G.; Cavarape, A.; Sechi, L.A. Cellular mechanisms of insulin resistance in rats with fructose-induced hypertension. Am. J. Hypertens. 2003, 16, 973–978. [Google Scholar] [CrossRef]
- Kho, M.C.; Lee, Y.J.; Ahn, Y.M.; Choi, Y.H.; Kim, A.Y.; Kang, D.G.; Lee, H.S. Effects of ethanol extract of gastrodia elata blume on high-fructose induced metabolic syndrome. FASEB J. 2013, 27, 1108–1113. [Google Scholar]
- Dandona, P.; Aljada, A.; Bandyopadhyay, A. Inflammation: The link between insulin resistance, obesity and diabetes. Trends Immunol. 2004, 25, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–999. [Google Scholar] [CrossRef] [PubMed]
- Rudin, E.; Barzilai, N. Inflammatory peptides derived from adipose tissue. Immun. Ageing 2005, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yudkin, J.S.; Stehouwer, C.D.; Emeis, J.J.; Coppack, S.W. C-reactive protein in healthy subjects: Associations with obesity, insulin resistance, and endothelial dysfunction: A potential role for cytokines originating from adipose tissue? Arterioscler. Thromb. Vasc. Biol. 1999, 19, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.H.; Dawson, J.D.; Riley, W.A.; Lauer, R.M. Carotid intimal-medial thickness is related to cardiovascular risk factors measured from childhood through middle age: The Muscatine study. Circulation 2001, 104, 2815–2819. [Google Scholar] [CrossRef] [PubMed]
- Berenson, G.S.; Srinivasan, S.R.; Bao, W.; Newman, W.P.; Tracy, R.E.; Wattigney, W.A. Association between multiple cardiovascular risk factors and atherosclerosis in children and young adults. N. Engl. J. Med. 1998, 338, 1650–1656. [Google Scholar] [CrossRef] [PubMed]
- McGill, H.C., Jr.; McMahan, C.A.; Zieske, A.W.; Malcom, G.T.; Tracy, R.E.; Strong, J.P. Effects of nonlipid risk factors on atherosclerosis in youth with a favorable lipoprotein profile. Circulation 2001, 103, 1546–1550. [Google Scholar] [CrossRef] [PubMed]
- McGill, H.C., Jr.; McMahan, C.A.; Herderick, E.E.; Tracy, R.E.; Malcom, G.T.; Zieske, A.W.; Strong, J.P. Effects of coronary heart disease risk factors on atherosclerosis of selected regions of the aorta and right coronary artery. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Bonetti, P.O.; Lerman, L.O.; Lerman, A. Endothelial dysfunction: A marker of atherosclerotic risk. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Pepine, C.J. Clinical implications of endothelial dysfunction. Clin. Cardiol. 1998, 21, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Brunner, H.; Cockcroft, J.R.; Deanfield, J.; Donald, A.; Ferrannini, E.; Halcox, J.; Kiowski, W.; Lüscher, T.F.; Mancia, G.; Natali, A. Endothelial function and dysfunction. Part II: Association with cardiovascular risk factors and diseases. A statement by the working group on endothelins and endothelial factors of the european society of hypertension. J. Hypertens. 2005, 23, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Davignon, J.; Ganz, P. Role of endothelial dysfunction in atherosclerosis. Circulation 2004, 109, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Dawson, V.; Brahmbhatt, H.; Mong, J.; Dawson, T. Expression of inducible nitric oxide synthase causes delayed neurotoxicity in primary mixed neuronal-glial cortical cultures. Neuropharmacology 1994, 33, 1425–1430. [Google Scholar] [CrossRef]
- Rees, D.D.; Palmer, R.M.; Moncada, S. Role of endothelium-derived nitric oxide in the regulation of blood pressure. Proc. Natl. Acad. Sci. USA 1989, 86, 3375–3378. [Google Scholar] [CrossRef] [PubMed]
- Oliver, F.J.; de la Rubia, G.; Feener, E.P.; Lee, M.E.; Loeken, M.R.; Shiba, T.; Quertermous, T.; King, G.L. Stimulation of endothelin-1 gene expression by insulin in endothelial cells. J. Biol. Chem. 1991, 266, 23251–23256. [Google Scholar] [PubMed]
- Verma, S.; Bhanot, S.; McNeill, J.H. Effect of chronic endothelin blockade in hyperinsulinemic hypertensive rats. Am. J. Physiol. 1995, 269, H2017–H2021. [Google Scholar] [PubMed]
- White, J.S. Challenging the fructose hypothesis: New perspectives on fructose consumption and metabolism. Adv. Nutr. 2013, 4, 246–256. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RD | 30Frc | 45Fat | 30Frc + 45Fat | |||||
|---|---|---|---|---|---|---|---|---|
| % | g | kcal | g | kcal | g | kcal | g | kcal |
| Protein | 20 | 20 | 20 | 20 | 24 | 20 | 24 | 20 |
| Carbohydrate | 64 | 64 | 64 | 64 | 41 | 35 | 41 | 35 |
| Fat | 7 | 16 | 7 | 16 | 24 | 45 | 24 | 45 |
| Total | 100 | 100 | 100 | 100 | ||||
| kcal/gm | 4.0 | 4.0 | 4.8 | 4.8 | ||||
| Ingredient | g | kcal | g | kcal | g | kcal | g | kcal |
| Casein, 80 Mesh | 200 | 800 | 200 | 800 | 200 | 800 | 200 | 800 |
| l-Cystine | 3 | 12 | 3 | 12 | 3 | 12 | 3 | 12 |
| Corn Starch | 397.5 | 1590 | 229.5 | 918 | 137 | 548 | 0 | 0 |
| Maltodextrin 10 | 132 | 528 | 100 | 400 | 100 | 400 | 37 | 148 |
| Sucrose | 100 | 400 | 0 | 0 | 100 | 400 | 0 | 0 |
| Fructose | 0 | 0 | 300 | 1200 | 0 | 0 | 300 | 1200 |
| Cellulose | 50 | 0 | 50 | 0 | 50 | 0 | 50 | 0 |
| Soybean Oil | 70 | 630 | 70 | 630 | 26 | 234 | 26 | 234 |
| t-Butylhydroquinone | 0.014 | 0 | 0.014 | 0 | 0.014 | 0 | 0.014 | 0 |
| Lard | 0 | 0 | 0 | 0 | 174 | 1566 | 174 | 1566 |
| Mineral Mix | 35 | 0 | 35 | 0 | 35 | 0 | 35 | 0 |
| Vitamin Mix | 10 | 40 | 10 | 40 | 10 | 40 | 10 | 40 |
| Choline Bitartrate | 2.5 | 0 | 2.5 | 0 | 2.5 | 0 | 2.5 | 0 |
| Total | 1000 | 4000 | 1000 | 4000 | 837.5 | 4000 | 837.5 | 4000 |
| RD (n = 8) | 30Frc (n = 8) | 45Fat (n = 8) | 30Frc + 45Fat (n = 8) | |
|---|---|---|---|---|
| Initial body weight (g) | 74.6 ± 2.6 | 74.9 ± 5.0 | 74.5 ± 4.71 | 74.4 ± 4.1 |
| Final body weight (g) | 251.3 ± 9.6 b | 263.3 ± 15.0 a,b | 278.1 ± 22.1 a | 277.2 ± 16.6 a |
| Weight gain (g) | 177.2 ± 9.8 b | 188.0 ± 13.4 a,b | 204.3 ± 22.6 a | 201.9 ± 17.7 a |
| Food intake (g/day) | 12.5 ± 0.8 a | 13.2 ± 0.9 a | 11.3 ± 0.9 b | 11.5 ± 0.8 b |
| Calorie intake (kcal/day) | 49.8 ± 2.9 | 52.4 ± 4.4 | 53.8 ± 4.6 | 54.7 ± 4.7 |
| Food efficiency ratio | 0.05 ± 0.00 | 0.05 ± 0.00 | 0.05 ± 0.01 | 0.05 ± 0.00 |
| RD (n = 8) | 30Frc (n = 8) | 45Fat (n = 8) | 30Frc + 45Fat (n = 8) | |
|---|---|---|---|---|
| Glucose (mg/dL) | 143.6 ± 10.3 | 142.0 ± 9.2 | 141.4 ± 14.9 | 136.6 ± 15.8 |
| Insulin (ng/mL) | 1.2 ± 1.4 | 2.2 ± 1.7 | 0.9± 0.3 | 1.3 ± 0.9 |
| HOMA-IR | 8.1 ± 9.1 | 12.7 ± 11.7 | 5.9± 2.4 | 8.3 ± 6.2 |
| QUICKI | 0.5 ± 0.07 | 0.4 ± 0.07 | 0.5± 0.05 | 0.5 ± 0.07 |
| RD (n = 8) | 30Frc (n = 8) | 45Fat (n = 8) | 30Frc + 45Fat (n = 8) | |
|---|---|---|---|---|
| TG (mg/dL) | 71.4 ± 19.8 b | 108.6 ± 25.2 a | 88.6 ± 26.9 a,b | 93.9 ± 24.1 a,b |
| Total-C (mg/dL) | 107.9 ± 19.7 | 114.6 ± 17.5 | 97.8 ± 15.4 | 96.3 ± 18.4 |
| HDL-C (mg/dL) | 98.1 ± 12.4 | 103.8 ± 10.9 | 92.1 ± 13.4 | 91.8 ± 14.7 |
| AI | 0.09 ± 0.07 | 0.10 ± 0.05 | 0.06 ± 0.03 | 0.05 ± 0.05 |
| RD (n = 8) | 30Frc (n = 8) | 45Fat (n = 8) | 30Frc + 45Fat (n = 8) | |
|---|---|---|---|---|
| TNF-α (pg/mL) | 55.1 ± 4.9 | 51.5 ± 5.5 | 47.0 ± 4.8 | 48.3 ± 10.9 |
| IL-6 (pg/mL) | 11.6 ± 4.2 | 18.2 ± 11.6 | 12.6 ± 4.0 | 15.5 ± 8.3 |
| PAI-1 (pg/mL) | 100.8 ± 13.3 | 101.5 ± 9.4 | 96.0 ± 4.8 | 99.4 ± 20.5 |
| RD (n = 8) | 30Frc (n = 8) | 45Fat (n = 8) | 30Frc + 45Fat (n = 8) | |
|---|---|---|---|---|
| Wall thickness (μm) | 18.5 ± 0.5 b | 23.6 ± 0.9 a | 23.7 ± 2.8 a | 22.5 ± 2.2 a |
| Lumen diameter (μm) | 306.8 ± 54.4 | 248.4 ± 25.1 | 243.9 ± 29.4 | 266.9 ± 29.0 |
| Wall thickness/ Lumen ratio (μm/μm) | 0.061 ± 0.009 b | 0.096 ± 0.001 a | 0.098 ± 0.014 a | 0.085 ± 0.005 a |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, S.; Ahn, H.; Park, Y.K. High Dietary Fructose Intake on Cardiovascular Disease Related Parameters in Growing Rats. Nutrients 2017, 9, 11. https://doi.org/10.3390/nu9010011
Yoo S, Ahn H, Park YK. High Dietary Fructose Intake on Cardiovascular Disease Related Parameters in Growing Rats. Nutrients. 2017; 9(1):11. https://doi.org/10.3390/nu9010011
Chicago/Turabian StyleYoo, SooYeon, Hyejin Ahn, and Yoo Kyoung Park. 2017. "High Dietary Fructose Intake on Cardiovascular Disease Related Parameters in Growing Rats" Nutrients 9, no. 1: 11. https://doi.org/10.3390/nu9010011
APA StyleYoo, S., Ahn, H., & Park, Y. K. (2017). High Dietary Fructose Intake on Cardiovascular Disease Related Parameters in Growing Rats. Nutrients, 9(1), 11. https://doi.org/10.3390/nu9010011