Chronic High Dose Zinc Supplementation Induces Visceral Adipose Tissue Hypertrophy without Altering Body Weight in Mice


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Study
2.2. Insulin Tolerance Test and Glucose Tolerance Test
2.3. Zinc Content Analysis
2.4. Serum Metabolites Profile Analysis
2.5. Serum Hormone Levels Measurement
2.6. Fat Tissue Histology Staining
2.7. Cell Culture
2.8. RNA Extraction and Real-Time PCR
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
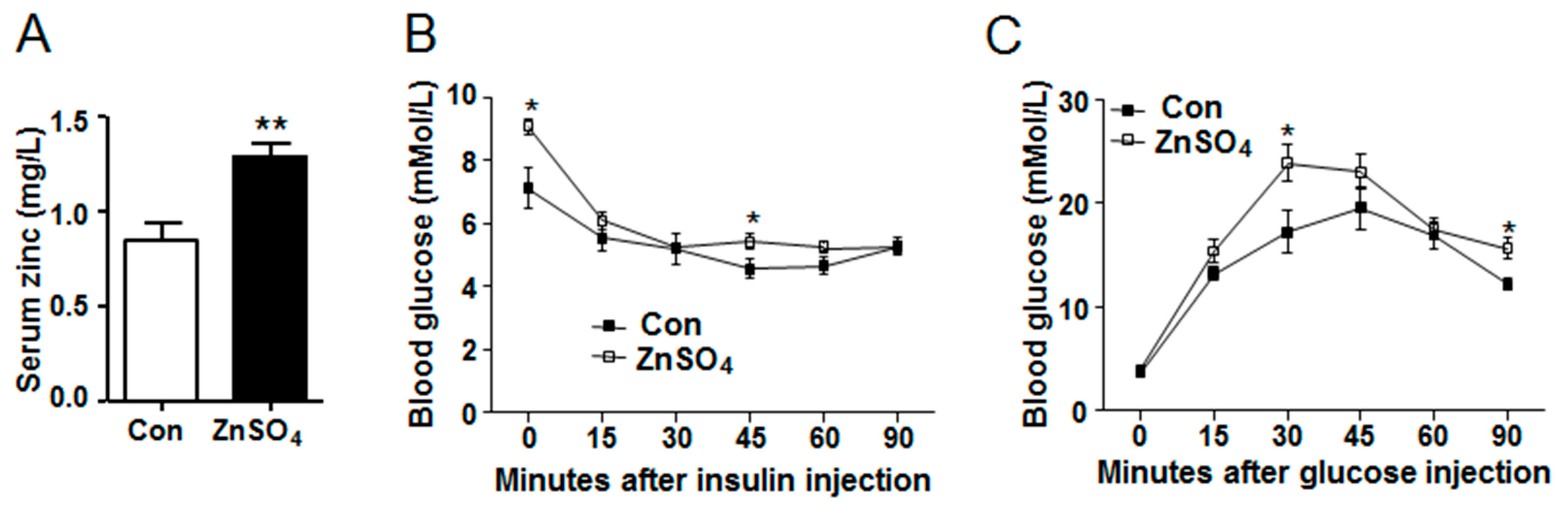
3.1. Chronic Zinc Supplementation Induced Glucose Intolerance
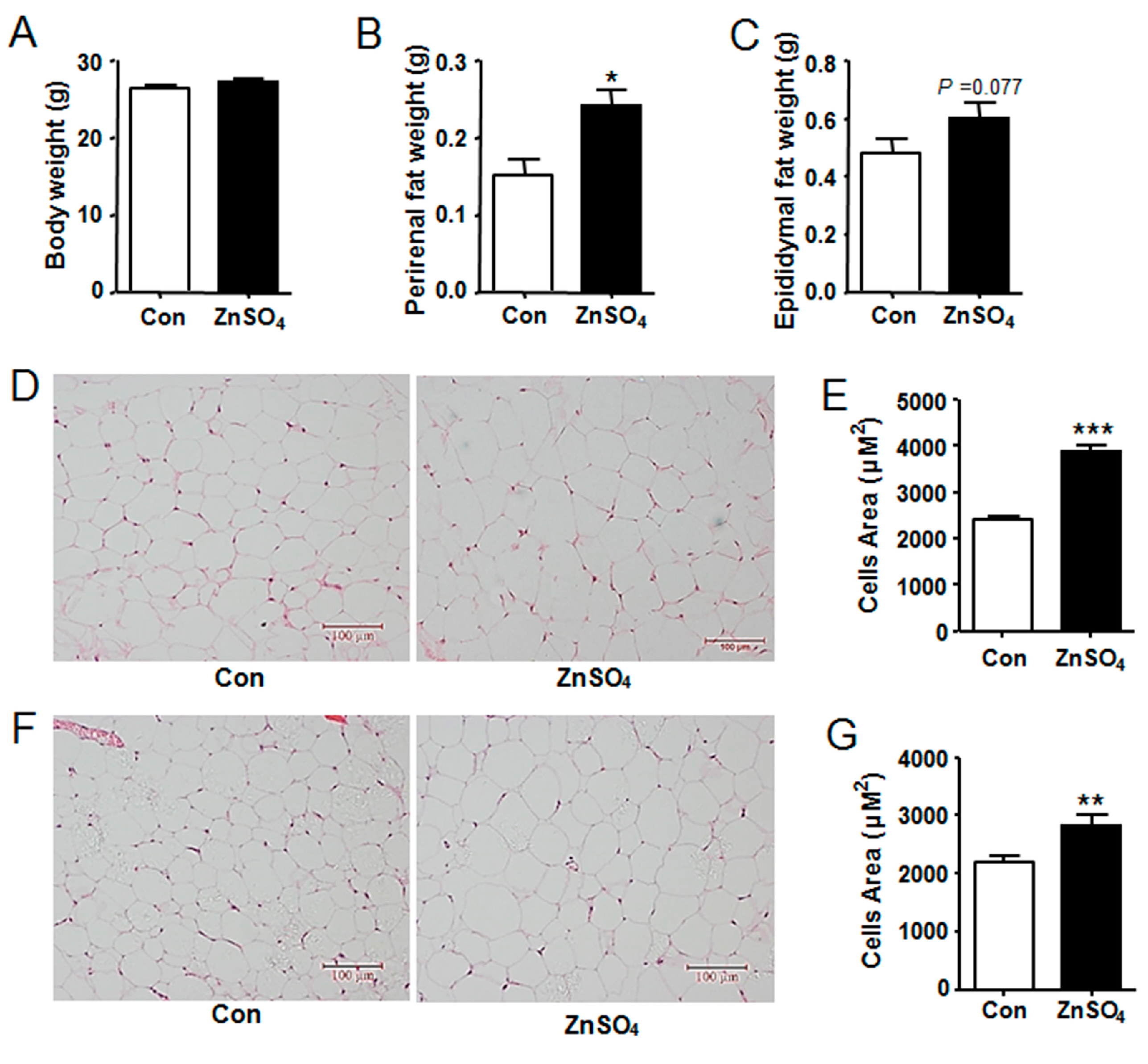
3.2. Chronic Zinc Supplementation Induced Visceral Adipose Tissue Hypertrophy
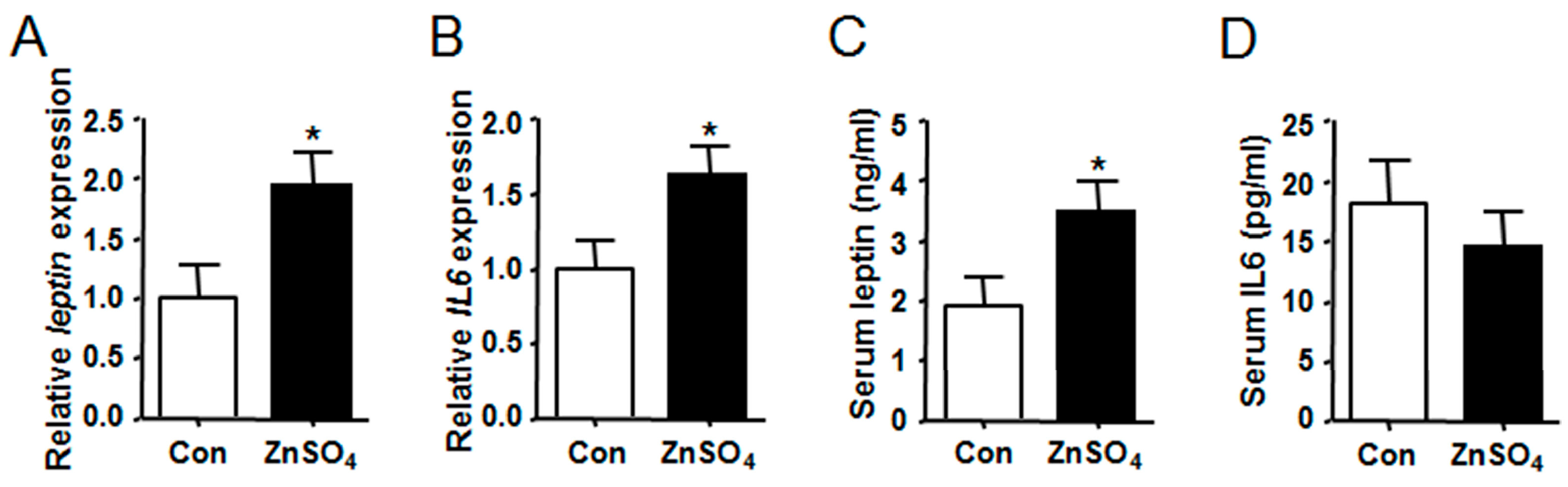
3.3. Chronic Zinc Supplementation Stimulated the Expression and Secretion of Leptin in Visceral Adipose Tissue
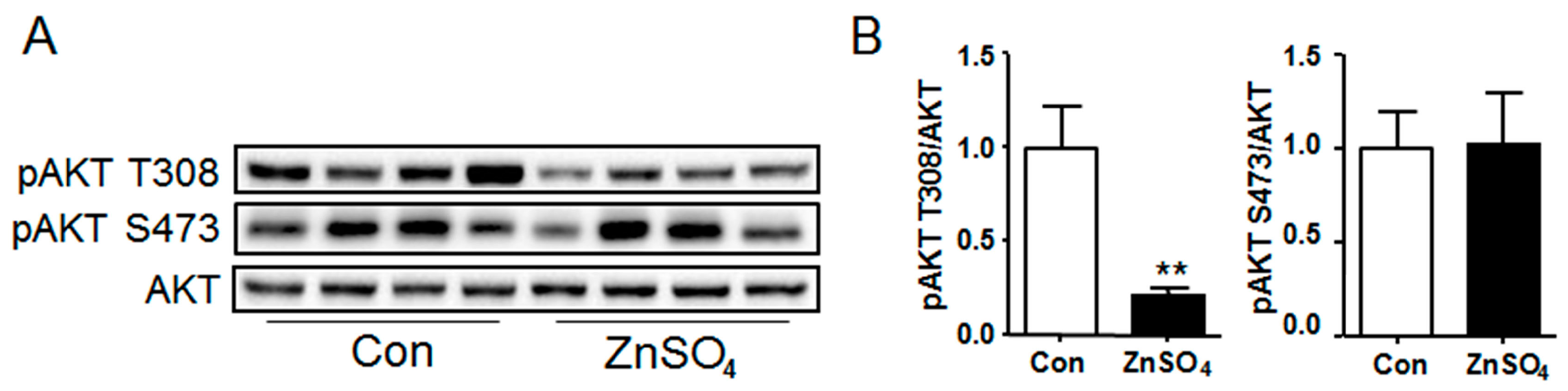
3.4. Chronic Zinc Supplementation Attenuated AKT Phosphorylation in Perirenal Adipose Tissue
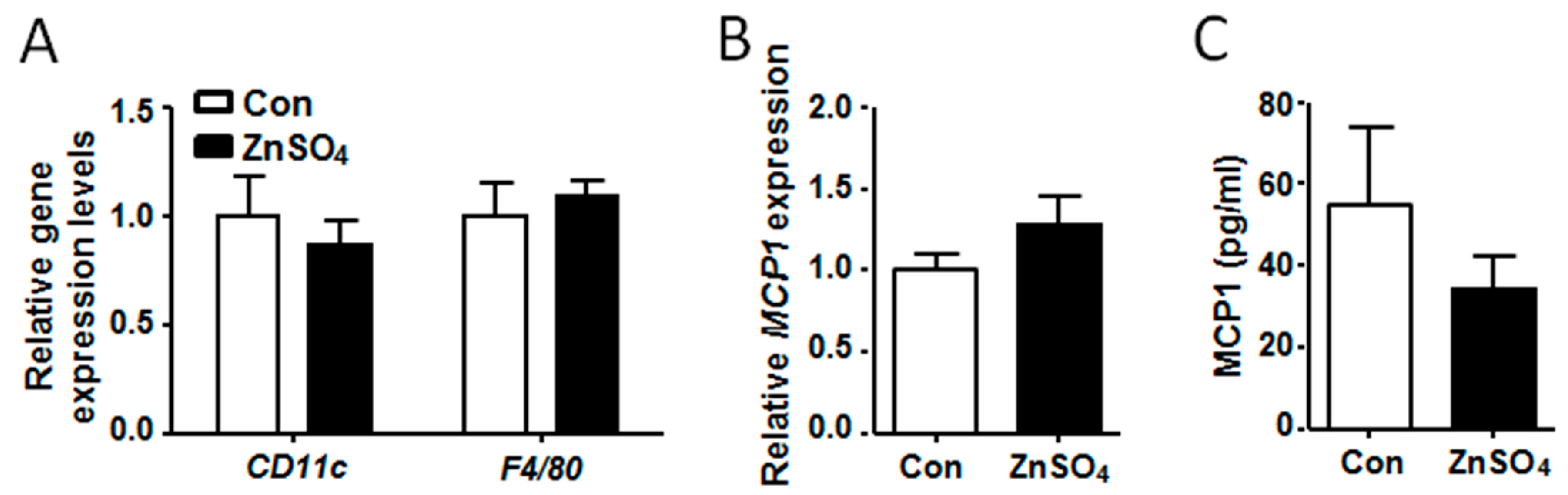
3.5. Chronic Zinc Supplementation Did Not Alter Macrophage Content in the Adipose Tissue
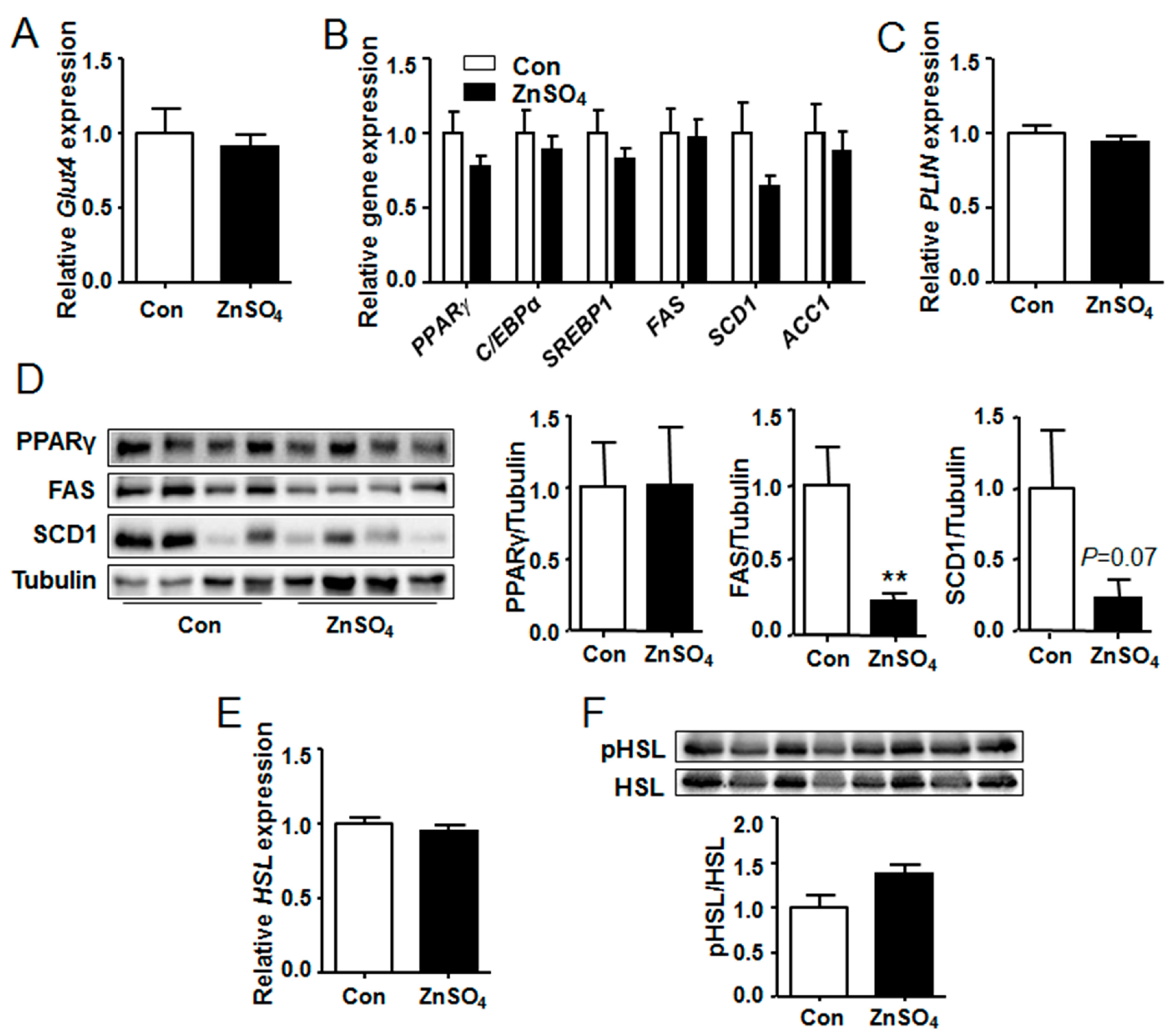
3.6. The Protein Levels of FAS and SCD1 Were Decreased in the Perirenal Adipose Tissue of Chronic Zinc-Supplemented Mice
3.7. Chronic Zinc Supplementation Did Not Alter the Metabolic Profiles of Serum Lipids
4. Discussion
4.1. Correlation of Chronic Zinc Supplementation with Systemic Glucose Clearance
4.2. Correlation of Chronic Zinc Supplementation with Adipose Accumulation in Visceral Adipose Tissue
4.3. Correlation of Chronic Zinc Supplementation with Adipokines and Cytokines
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Myers, S.; Shastri, M.D.; Adulcikas, J.; Sohal, S.S.; Norouzi, S. Zinc and gastrointestinal disorders: A role for the zinc transporters Zips and ZnTs. Curr. Pharm. Des. 2017, 23, 2328–2332. [Google Scholar] [CrossRef] [PubMed]
- Hojyo, S.; Fukada, T. Roles of Zinc Signaling in the Immune System. J. Immunol. Res. 2016, 2016, 6762343. [Google Scholar] [CrossRef] [PubMed]
- Nissensohn, M.; Sanchez-Villegas, A.; Fuentes Lugo, D.; Henriquez Sanchez, P.; Doreste Alonso, J.; Pena Quintana, L.; Ruano, C.; Lowe, N.L.; Hall Moran, V.; Skinner, A.L.; et al. Effect of Zinc Intake on Growth in Infants: A Meta-analysis. Crit. Rev. Food Sci. Nutr. 2016, 56, 350–363. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, P.; Benedetti, G.; Albarede, F.; Miossec, P. Zinc and its role in immunity and inflammation. Autoimmun. Rev. 2015, 14, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, A.J. Zinc, aging, and immunosenescence: An overview. Pathobiol. Aging Age Relat. Dis. 2015, 5, 25592. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Zhao, Y.; Sun, X.; Song, Z.; McClain, C.J.; Zhou, Z. Dietary zinc deficiency exaggerates ethanol-induced liver injury in mice: Involvement of intrahepatic and extrahepatic factors. PLoS ONE 2013, 8, e76522. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.J.; Bao, S.; Bolin, E.R.; Burris, D.L.; Xu, X.; Sun, Q.; Killilea, D.W.; Shen, Q.; Ziouzenkova, O.; Belury, M.A.; et al. Zinc deficiency augments leptin production and exacerbates macrophage infiltration into adipose tissue in mice fed a high-fat diet. J. Nutr. 2013, 143, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Fung, E.B.; Gildengorin, G.; Talwar, S.; Hagar, L.; Lal, A. Zinc status affects glucose homeostasis and insulin secretion in patients with thalassemia. Nutrients 2015, 7, 4296–4307. [Google Scholar] [CrossRef] [PubMed]
- Cruz, K.J.; Morais, J.B.; de Oliveira, A.R.; Severo, J.S.; Marreiro, D.D. The Effect of Zinc Supplementation on Insulin Resistance in Obese Subjects: A Systematic Review. Biol. Trace Elem. Res. 2017, 176, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Umrani, R.D.; Paknikar, K.M. Zinc oxide nanoparticles show antidiabetic activity in streptozotocin-induced Type 1 and 2 diabetic rats. Nanomed. Lond. 2014, 9, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Bao, W.; Zhang, Y.; Rong, Y.; Wang, X.; Jin, Y.; Song, Y.; Yao, P.; Sun, C.; Hu, F.B.; et al. Interactions between zinc transporter-8 gene (SLC30A8) and plasma zinc concentrations for impaired glucose regulation and type 2 diabetes. Diabetes 2014, 63, 1796–1803. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Siddique, K.U.; Ashfaq, F.; Ali, W.; Reddy, H.D.; Mishra, A. Effect of high-dose zinc supplementation with oral hypoglycemic agents on glycemic control and inflammation in type-2 diabetic nephropathy patients. J. Nat. Sci. Biol. Med. 2013, 4, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Zhong, W.; Liu, J.; Song, Z.; McClain, C.J.; Kang, Y.J.; Zhou, Z. Zinc supplementation reverses alcohol-induced steatosis in mice through reactivating hepatocyte nuclear factor-4alpha and peroxisome proliferator-activated receptor-alpha. Hepatology 2009, 50, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, C.; Yang, S.H.; Kim, J.G.; Jeon, T.I.; Yoon, B.H.; Lee, J.Y.; Lee, E.Y.; Choi, S.G.; Hwang, S.G. Zinc-chelated Vitamin C Stimulates Adipogenesis of 3T3-L1 Cells. Asian-Australas. J. Anim. Sci. 2013, 26, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.B.; Wang, M.S.; Wang, Z.S.; Zhou, A.M.; Zhang, X.M.; Dong, X.W.; Peng, Q.H. Supplementation dietary zinc levels on growth performance, carcass traits, and intramuscular fat deposition in weaned piglets. Biol. Trace Elem. Res. 2014, 161, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Kusminski, C.M.; Scherer, P.E. Adipose tissue remodeling and obesity. J. Clin. Invest. 2011, 121, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- McTernan, P.G.; Harte, A.L.; Anderson, L.A.; Green, A.; Smith, S.A.; Holder, J.C.; Barnett, A.H.; Eggo, M.C.; Kumar, S. Insulin and rosiglitazone regulation of lipolysis and lipogenesis in human adipose tissue in vitro. Diabetes 2002, 51, 1493–1498. [Google Scholar] [CrossRef] [PubMed]
- Kuryszko, J.; Slawuta, P.; Sapikowski, G. Secretory function of adipose tissue. Pol. J. Vet. Sci. 2016, 19, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.C.; Lee, J. Cellular and molecular players in adipose tissue inflammation in the development of obesity-induced insulin resistance. Biochim. Biophys. Acta 2014, 1842, 446–462. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Lun, M.; Wang, M.; Senyo, S.E.; Guillermier, C.; Patwari, P.; Steinhauser, M.L. Loss of white adipose hyperplastic potential is associated with enhanced susceptibility to insulin resistance. Cell Metab. 2014, 20, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, K.; Rockstroh, D.; Wagner, I.V.; Weise, S.; Tauscher, R.; Schwartze, J.T.; Loffler, D.; Buhligen, U.; Wojan, M.; Till, H.; et al. Evidence of early alterations in adipose tissue biology and function and its association with obesity-related inflammation and insulin resistance in children. Diabetes 2015, 64, 1249–1261. [Google Scholar] [CrossRef] [PubMed]
- White, U.A.; Stephens, J.M. Transcriptional factors that promote formation of white adipose tissue. Mol. Cell. Endocrinol. 2010, 318, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, T.; Li, J.; Wang, S.; Qiu, F.; Yu, H.; Zhang, Y.; Wang, T. Effects of Natural Products on Fructose-Induced Nonalcoholic Fatty Liver Disease (NAFLD). Nutrients 2017, 9, 96. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Guan, Y.; Shi, X.; Ding, H.; Song, Y.; Li, C.; Liu, R.; Liu, G. Effects of high zinc levels on the lipid synthesis in rat hepatocytes. Biol. Trace Elem. Res. 2013, 154, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Jiao, P.; Helou, Y.; Li, Y.; He, Q.; Walters, M.S.; Salomon, A.; Xu, H. Mitogen-activated protein kinase phosphatase 3 (MKP-3)-deficient mice are resistant to diet-induced obesity. Diabetes 2014, 63, 2924–2934. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Huang, X.; Jiang, D.; Hua, L.; Zhuo, Y.; Wu, D. Endoplasmic Reticulum Stress Inducer Tunicamycin Alters Hepatic Energy Homeostasis in Mice. Int. J. Mol. Sci. 2017, 18, 1710. [Google Scholar] [CrossRef]
- Feng, B.; Jiao, P.; Nie, Y.; Kim, T.; Jun, D.; van Rooijen, N.; Yang, Z.; Xu, H. Clodronate liposomes improve metabolic profile and reduce visceral adipose macrophage content in diet-induced obese mice. PLoS ONE. 2011, 6, e24358. [Google Scholar] [CrossRef] [PubMed]
- Marchesini, G.; Bugianesi, E.; Ronchi, M.; Flamia, R.; Thomaseth, K.; Pacini, G. Zinc supplementation improves glucose disposal in patients with cirrhosis. Metabolism 1998, 47, 792–798. [Google Scholar] [CrossRef]
- Quarterman, J.; Mills, C.F.; Humphries, W.R. The reduced secretion of, and sensitivity to insulin in zinc-deficient rats. Biochem. Biophys. Res. Commun. 1966, 25, 354–358. [Google Scholar] [CrossRef]
- Shisheva, A.; Gefel, D.; Shechter, Y. Insulinlike effects of zinc ion in vitro and in vivo. Preferential effects on desensitized adipocytes and induction of normoglycemia in streptozocin-induced rats. Diabetes 1992, 41, 982–988. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, S. Effect of zinc supplementation on insulin resistance and metabolic risk factors in obese Korean women. Nutr. Res. Pract. 2012, 6, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Lobene, A.J.; Kindler, J.M.; Jenkins, N.T.; Pollock, N.K.; Laing, E.M.; Grider, A.; Lewis, R.D. Zinc Supplementation Does Not Alter Indicators of Insulin Secretion and Sensitivity in Black and White Female Adolescents. J. Nutr. 2017, 147, 1296–1300. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. Mechanisms for insulin resistance: Common threads and missing links. Cell 2012, 148, 852–871. [Google Scholar] [CrossRef] [PubMed]
- Rothman, D.L.; Magnusson, I.; Katz, L.D.; Shulman, R.G.; Shulman, G.I. Quantitation of hepatic glycogenolysis and gluconeogenesis in fasting humans with 13C NMR. Science 1991, 254, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Stofkova, A. Leptin and adiponectin: From energy and metabolic dysbalance to inflammation and autoimmunity. Endocr. Regul. 2009, 43, 157–168. [Google Scholar] [CrossRef] [PubMed]
- DePaoli, A.M. 20 years of leptin: Leptin in common obesity and associated disorders of metabolism. J. Endocrinol. 2014, 223, T71–T81. [Google Scholar] [CrossRef] [PubMed]
- Baltaci, A.K.; Mogulkoc, R.; Halifeoglu, I. Effects of zinc deficiency and supplementation on plasma leptin levels in rats. Biol. Trace Elem. Res. 2005, 104, 41–46. [Google Scholar] [CrossRef]
- Ott, E.S.; Shay, N.F. Zinc deficiency reduces leptin gene expression and leptin secretion in rat adipocytes. Exp. Biol. Med. 2001, 226, 841–846. [Google Scholar] [CrossRef]
- Argani, H.; Mahdavi, R.; Ghorbani-haghjo, A.; Razzaghi, R.; Nikniaz, L.; Gaemmaghami, S.J. Effects of zinc supplementation on serum zinc and leptin levels, BMI, and body composition in hemodialysis patients. J. Trace Elem. Med. Biol. 2014, 28, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest. 2003, 112, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Abella, V.; Scotece, M.; Conde, J.; Pino, J.; Gonzalez-Gay, M.A.; Gomez-Reino, J.J.; Mera, A.; Lago, F.; Gomez, R.; Gualillo, O. Leptin in the interplay of inflammation, metabolism and immune system disorders. Nat. Rev. Rheumatol. 2017, 13, 100–109. [Google Scholar] [CrossRef] [PubMed]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Jiang, D.; Zhu, Y.; Fang, Z.; Che, L.; Lin, Y.; Xu, S.; Li, J.; Huang, C.; Zou, Y.; et al. Chronic High Dose Zinc Supplementation Induces Visceral Adipose Tissue Hypertrophy without Altering Body Weight in Mice. Nutrients 2017, 9, 1138. https://doi.org/10.3390/nu9101138
Huang X, Jiang D, Zhu Y, Fang Z, Che L, Lin Y, Xu S, Li J, Huang C, Zou Y, et al. Chronic High Dose Zinc Supplementation Induces Visceral Adipose Tissue Hypertrophy without Altering Body Weight in Mice. Nutrients. 2017; 9(10):1138. https://doi.org/10.3390/nu9101138
Chicago/Turabian StyleHuang, Xiaohua, Dandan Jiang, Yingguo Zhu, Zhengfeng Fang, Lianqiang Che, Yan Lin, Shengyu Xu, Jian Li, Chao Huang, Yuanfeng Zou, and et al. 2017. "Chronic High Dose Zinc Supplementation Induces Visceral Adipose Tissue Hypertrophy without Altering Body Weight in Mice" Nutrients 9, no. 10: 1138. https://doi.org/10.3390/nu9101138
APA StyleHuang, X., Jiang, D., Zhu, Y., Fang, Z., Che, L., Lin, Y., Xu, S., Li, J., Huang, C., Zou, Y., Li, L., Wu, D., & Feng, B. (2017). Chronic High Dose Zinc Supplementation Induces Visceral Adipose Tissue Hypertrophy without Altering Body Weight in Mice. Nutrients, 9(10), 1138. https://doi.org/10.3390/nu9101138