High Fat Diet with a High Monounsaturated Fatty Acid and Polyunsaturated/Saturated Fatty Acid Ratio Suppresses Body Fat Accumulation and Weight Gain in Obese Hamsters



Abstract
:1. Introduction
2. Materials and Methods
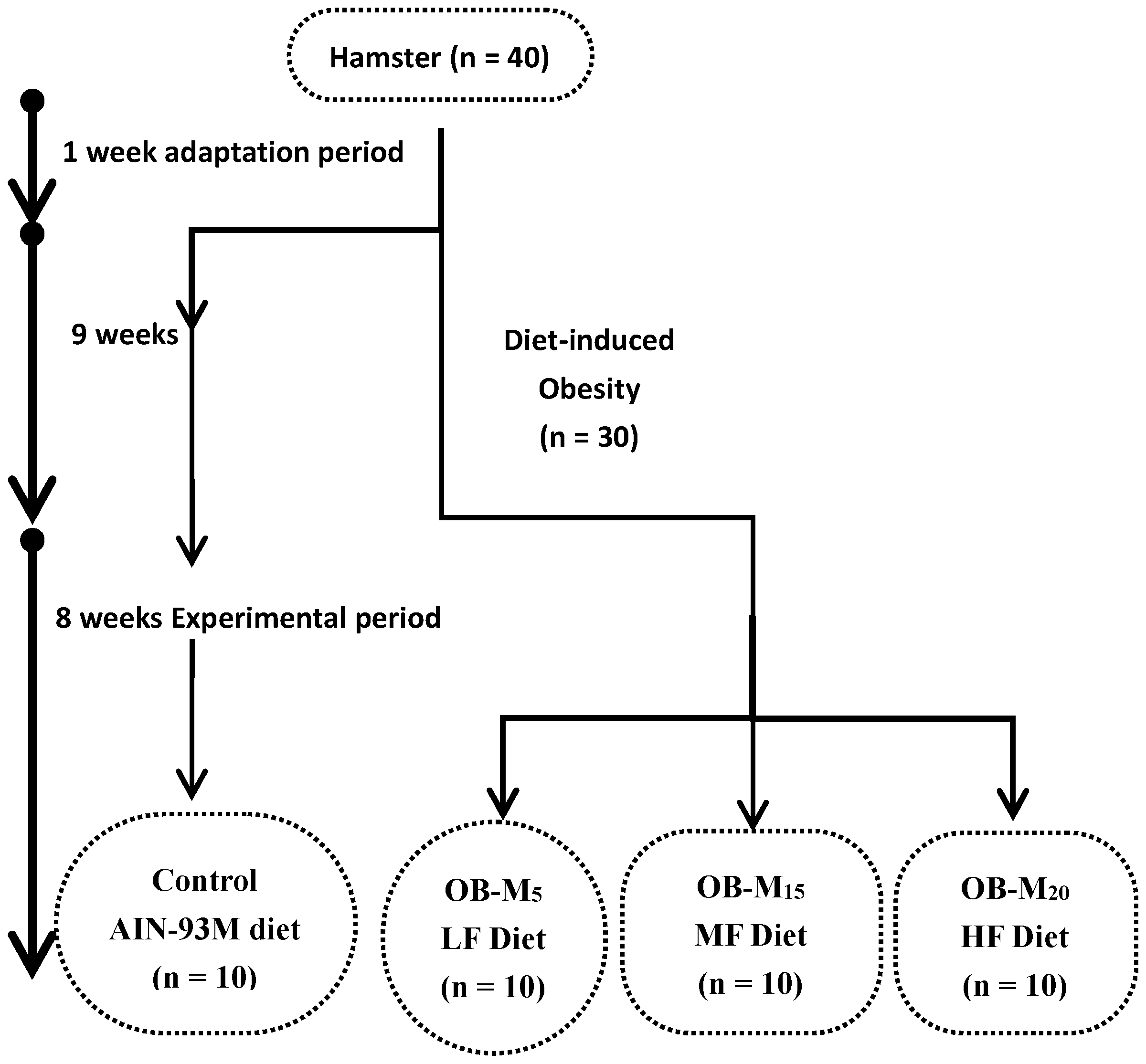
2.1. Animals and the Study Design
2.2. Fatty acid Composition Analysis
2.3. Plasma Measurements
2.4. Hepatic Lipid Contents and Enzyme Activities
2.5. Adipose Tissue LPL Enzyme Assay
2.6. Real-Time Quantitative Polymerase Chain Reaction (qPCR)
2.7. Histological Analyses
2.8. Statistical Analysis
3. Results
3.1. Body and Tissues Weights, Weight Gain, and Energy Consumption
3.2. Plasma Parameter Concentrations and Hepatic Lipid Content
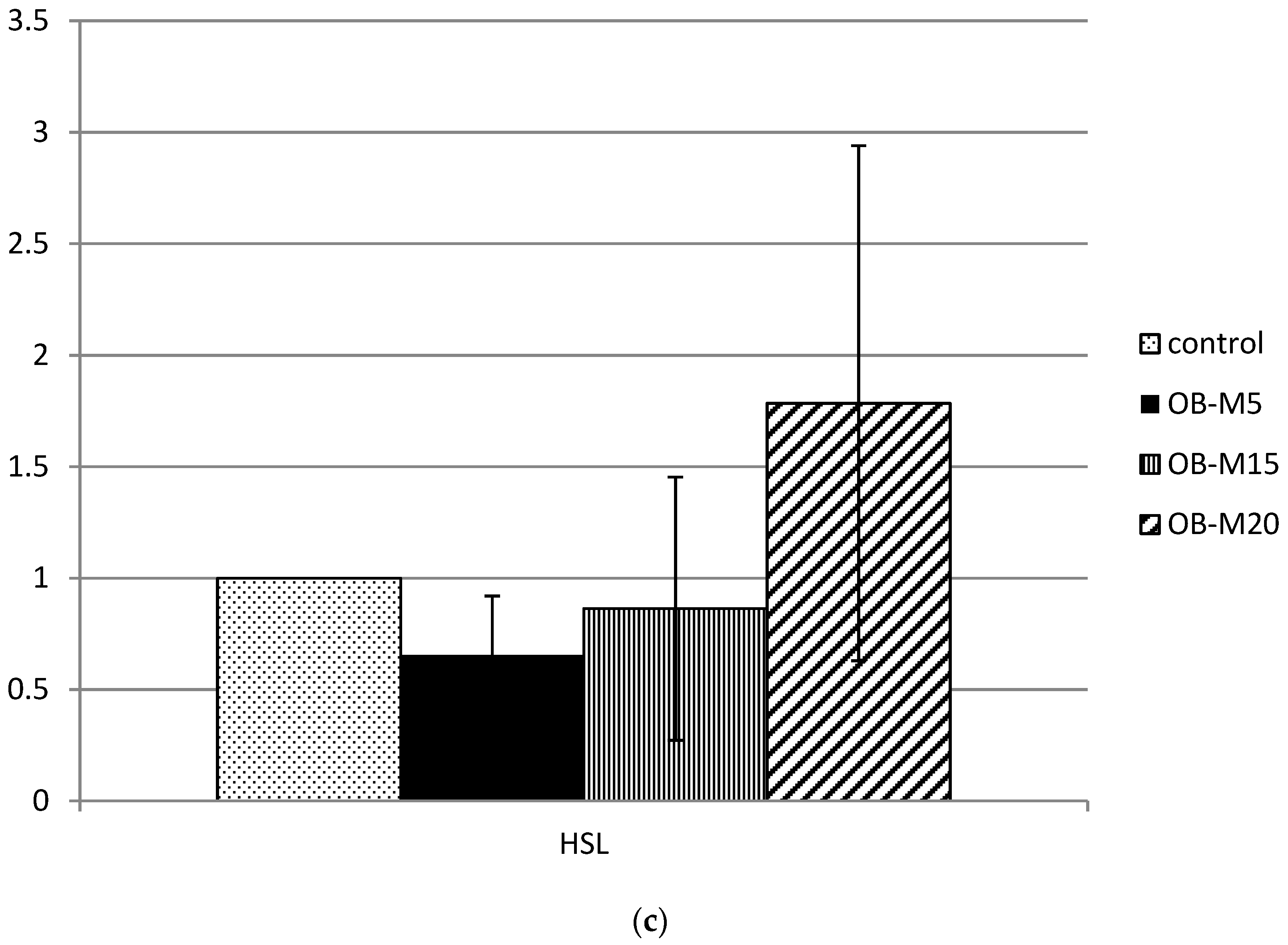
3.3. Hepatic Lipogenic and Lipolysis Enzyme Activities
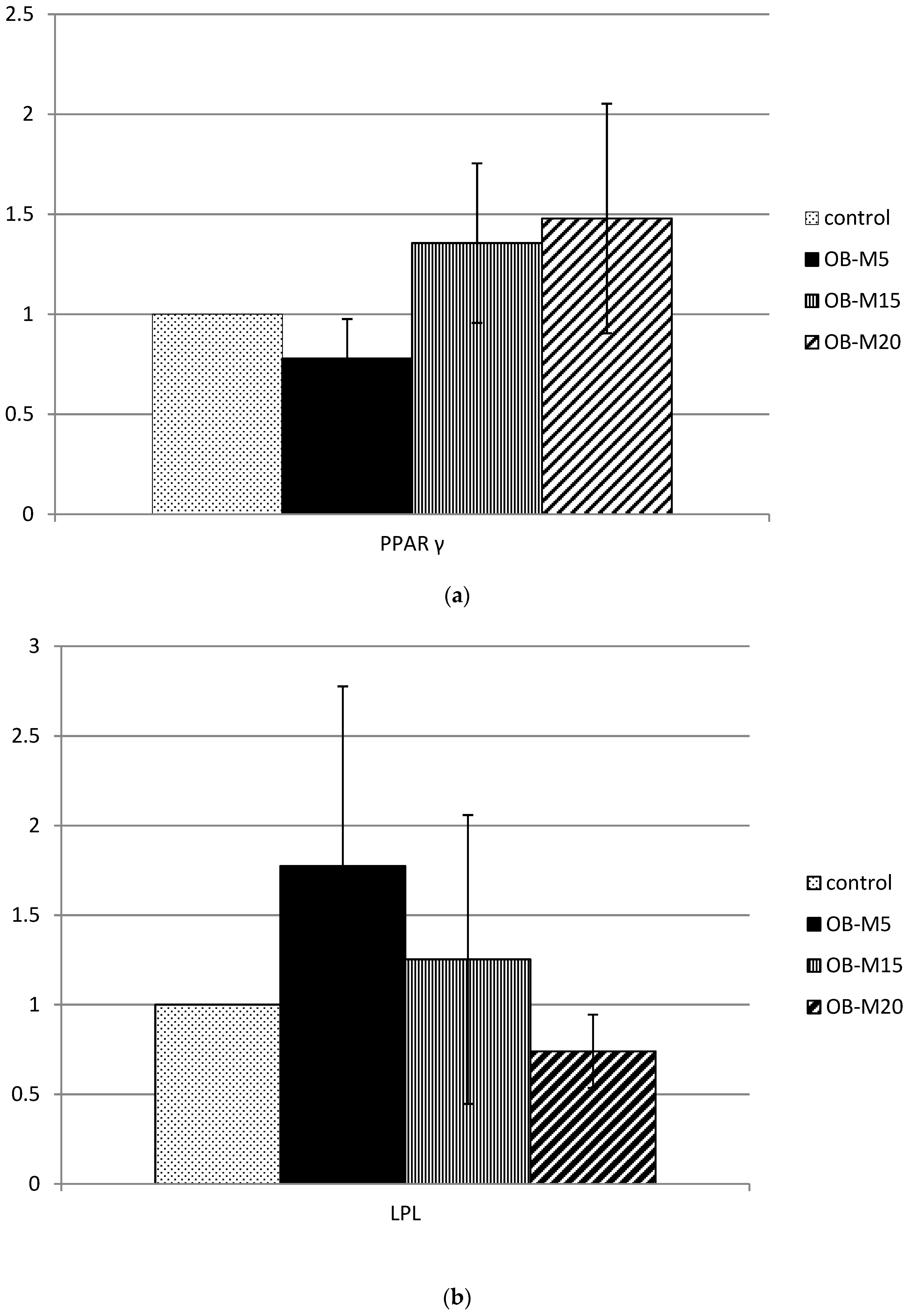
3.4. Adipose Tissue LPL Activities
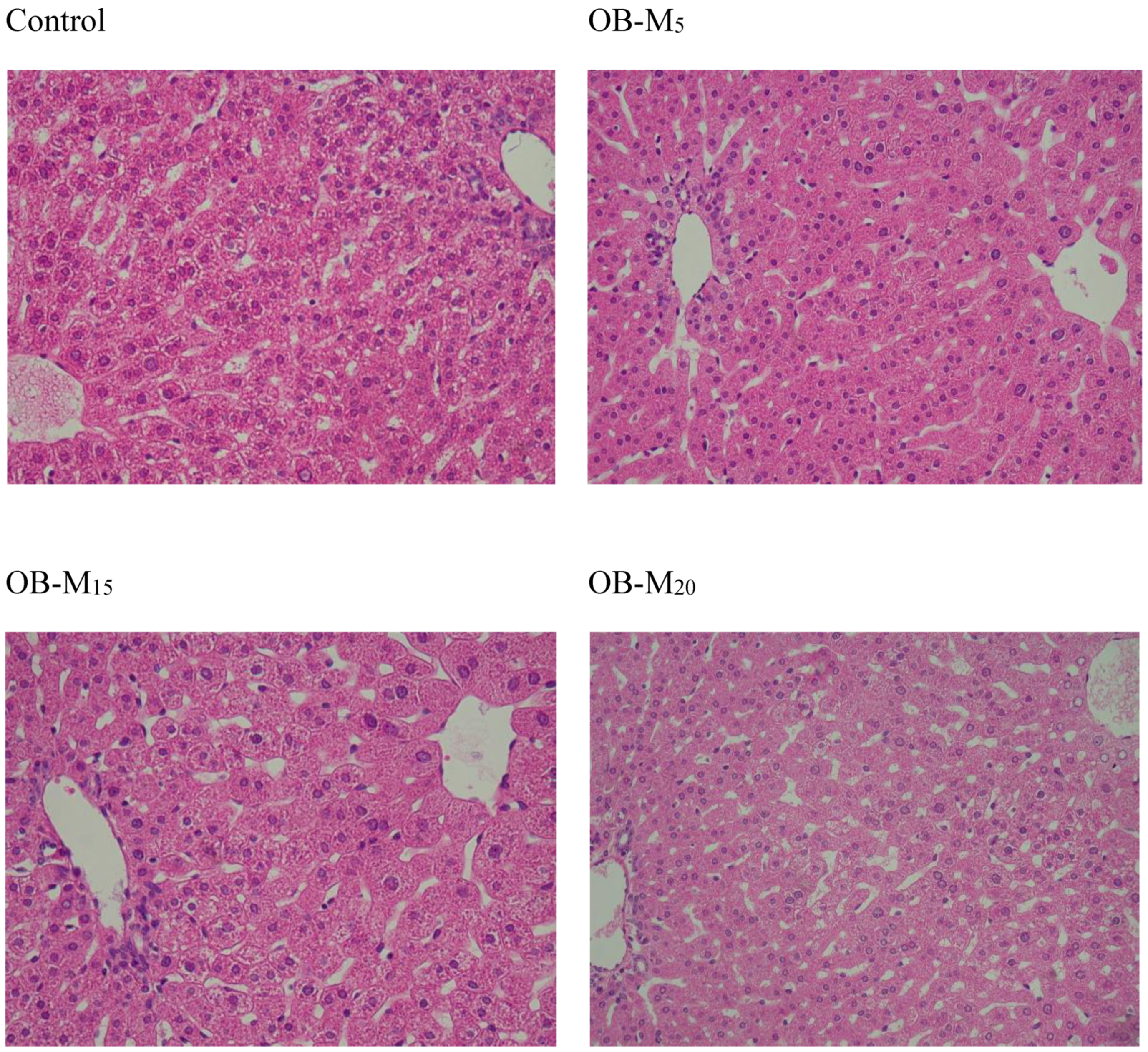
Liver Histological Analyses
3.5. Lipid Metabolism-Related Protein mRNA Expression in Adipose Tissue
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hajer, G.R.; van Haeften, T.W.; Visseren, F.L. Adipose tissue dysfunction in obesity, diabetes, and vascular diseases. Eur. Heart J. 2008, 29, 2959–2971. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.L.; Dive, C.; Renehan, A.G. Biological mechanisms linking obesity and cancer risk: New perspectives. Annu. Rev. Med. 2010, 61, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Choquet, H.; Meyre, D. Genetics of Obesity: What have we Learned? Curr. Genom. 2011, 12, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Schutz, Y.; Flatt, J.P.; Jequier, E. Failure of dietary fat intake to promote fat oxidation: A factor favoring the development of obesity. Am. J. Clin. Nutr. 1989, 50, 307–314. [Google Scholar] [PubMed]
- Hooper, L.; Abdelhamid, A.; Moore, H.J.; Douthwaite, W.; Skeaff, C.M.; Summerbell, C.D. Effect of reducing total fat intake on body weight: Systematic review and meta-analysis of randomised controlled trials and cohort studies. BMJ 2012, 345, e7666. [Google Scholar] [CrossRef] [PubMed]
- Mirza, N.M.; Palmer, M.G.; Sinclair, K.B.; McCarter, R.; He, J.; Ebbeling, C.B.; Ludwig, D.S.; Yanovski, J.A. Effects of a low glycemic load or a low-fat dietary intervention on body weight in obese Hispanic American children and adolescents: A randomized controlled trial. Am. J. Clin. Nutr. 2013, 97, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Harika, R.K.; Eilander, A.; Alssema, M.; Osendarp, S.J.; Zock, P.L. Intake of fatty acids in general populations worldwide does not meet dietary recommendations to prevent coronary heart disease: A systematic review of data from 40 countries. Ann. Nutr. Metab. 2013, 63, 229–238. [Google Scholar] [CrossRef] [PubMed]
- DeSalvo, K.B.; Olson, R.; Casavale, K.O. Dietary Guidelines for Americans. JAMA 2016, 315, 457–458. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Matsuo, T.; Tokuyama, K.; Shimomura, Y.; Suzuki, M. Diet-induced thermogenesis is lower in rats fed a lard diet than in those fed a high oleic acid safflower oil diet, a safflower oil diet or a linseed oil diet. J. Nutr. 1995, 125, 920–925. [Google Scholar] [PubMed]
- Piers, L.S.; Walker, K.Z.; Stoney, R.M.; Soares, M.J.; O’Dea, K. The influence of the type of dietary fat on postprandial fat oxidation rates: Monounsaturated (olive oil) vs saturated fat (cream). Int. J. Obes. Relat. Metab. Disord. 2002, 26, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.J.H.; Jew, S.; AbuMweis, S. The effect of dietary oleic, linoleic, and linolenic acids on fat oxidation and energy expenditure in healthy men. Metabolism 2008, 57, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Bruininx, E.; van den Borne, J.; van Heugten, E.; van Milgen, J.; Verstegen, M.; Gerrits, W. Oxidation of dietary stearic, oleic, and linoleic acids in growing pigs follows a biphasic pattern. J. Nutr. 2001, 141, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- McCloy, U.; Ryan, M.A.; Pencharz, P.B.; Ross, R.J.; Cunnane, S.C. A comparison of the metabolism of eighteen-carbon 13C-unsaturated fatty acids in healthy women. J. Lipid Res. 2004, 45, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Cooper, J.A. Effect of dietary fatty acid composition on substrate utilization and body weight maintenance in humans. Eur. J. Nutr. 2014, 53, 691–710. [Google Scholar] [CrossRef] [PubMed]
- Riccardi, G.; Rivellese, A.A. Dietary treatment of the metabolic syndrome—The optimal diet. Br. J. Nutr. 2000, 83 (Suppl. 1), S143–S148. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.J.; Ridgen, J.E.; Phang, P.T.; Birmingham, C.L. Influence of dietary fat polyunsaturated to saturated ratio on energy substrate utilization in obesity. Metabolism 1992, 41, 396–401. [Google Scholar] [CrossRef]
- Clandinin, M.T.; Wang, L.C.; Rajotte, R.V.; French, M.A.; Goh, Y.K.; Kielo, E.S. Increasing the dietary polyunsaturated fat content alters whole-body utilization of 16:0 and 10:0. Am. J. Clin. Nutr. 1995, 61, 1052–1057. [Google Scholar] [PubMed]
- Liao, F.H.; Liou, T.H.; Chiu, W.C.; Shieh, M.J.; Chien, Y.W. Differential effects of high MUFA with high or low P/S ratio (polyunsaturated to saturated fatty acids) on improving hepatic lipolytic enzymes and mediating PPARgamma related with lipoprotein lipase and hormone-sensitive lipase of white adipose tissue in diet-induced obese hamster. Int. J. Obes. 2010, 34, 1608–1617. [Google Scholar] [CrossRef]
- Liao, F.H.; Liou, T.H.; Shieh, M.J.; Chien, Y.W. Effects of different ratios of monounsaturated and polyunsaturated fatty acids to saturated fatty acids on regulating body fat deposition in hamsters. Nutrition 2010, 26, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.L.; West, K.L. Mechanisms by which dietary fatty acids modulate plasma lipids. J. Nutr. 2005, 135, 2075–2078. [Google Scholar] [PubMed]
- Schoonjans, K.; Martin, G.; Staels, B.; Auwerx, J. Peroxisome proliferator-activated receptors, orphans with ligands and functions. Curr. Opin. Lipidol. 1997, 8, 159–166. [Google Scholar] [CrossRef]
- Harrington, W.W.; Britt, C.S.; Wilson, J.G.; Milliken, N.O.; Binz, J.G.; Lobe, D.C.; Oliver, W.R.; Lewis, M.C.; Ignar, D.M. The Effect of PPARalpha, PPARdelta, PPARgamma, and PPARpan Agonists on Body Weight, Body Mass, and Serum Lipid Profiles in Diet-Induced Obese AKR/J Mice. PPAR Res. 2007, 2007, 97125. [Google Scholar] [CrossRef]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Regulation of adipocyte gene expression and differentiation by peroxisome proliferator activated receptor gamma. Curr. Opin. Genet. Dev. 1995, 5, 571–576. [Google Scholar] [CrossRef]
- Teruel, T.; Hernandez, R.; Rial, E.; Martin-Hidalgo, A.; Lorenzo, M. Rosiglitazone up-regulates lipoprotein lipase, hormone-sensitive lipase and uncoupling protein-1, and down-regulates insulin-induced fatty acid synthase gene expression in brown adipocytes of Wistar rats. Diabetologia 2005, 48, 1180–1188. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, I.J. Lipoprotein lipase and lipolysis: Central roles in lipoprotein metabolism and atherogenesis. J. Lipid Res. 1996, 37, 693–707. [Google Scholar]
- Ahmadian, M.; Wang, Y.; Sul, H.S. Lipolysis in adipocytes. Int. J. Biochem. Cell Biol. 2010, 42, 555–559. [Google Scholar] [CrossRef]
- Shimomura, I.; Bashmakov, Y.; Ikemoto, S.; Horton, J.D.; Brown, M.S.; Goldstein, J.L. Insulin selectively increases SREBP-1c mRNA in the livers of rats with streptozotocin-induced diabetes. Proc. Natl. Acad. Sci. USA 1999, 96, 13656–13661. [Google Scholar] [CrossRef]
- Shimomura, I.; Bashmakov, Y.; Ikemoto, S.; Horton, J.D.; Brown, M.S.; Goldstein, J.L. Sterol regulatory element binding protein-1 expression is suppressed by dietary polyunsaturated fatty acids. A mechanism for the coordinate suppression of lipogenic genes by polyunsaturated fats. J. Biol. Chem. 1999, 274, 23577–23583. [Google Scholar]
- Field, F.J.; Born, E.; Mathur, S.N. Fatty acid flux suppresses fatty acid synthesis in hamster intestine independently of SREBP-1 expression. J. Lipid Res. 2003, 44, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Jump, D.B.; Botolin, D.; Wang, Y.; Xu, J.; Christian, B.; Demeure, O. Fatty acid regulation of hepatic gene transcription. J. Nutr. 2005, 135, 2503–2506. [Google Scholar] [PubMed]
- Pelkman, C.L.; Fishell, V.K.; Maddox, D.H.; Pearson, T.A.; Mauger, D.T.; Kris-Etherton, P.M. Effects of moderate-fat (from monounsaturated fat) and low-fat weight-loss diets on the serum lipid profile in overweight and obese men and women. Am. J. Clin. Nutr. 2004, 79, 204–212. [Google Scholar] [PubMed]
- Yang, J.H.; Chang, J.S.; Chen, C.L.; Yeh, C.L.; Chien, Y.W. Effects of different amounts and types of dietary fatty acids on the body weight, fat accumulation, and lipid metabolism in hamsters. Nutrition 2016, 32, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Morrison, W.R.; Smith, L.M. Preparation of Fatty Acid Methyl Esters and Dimethylacetals from Lipids with Boron Fluoride—Methanol. J. Lipid Res. 1964, 5, 600–608. [Google Scholar] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Wade, G.N. Obesity without overeating in golden hamsters. Physiol. Behav. 1982, 29, 701–707. [Google Scholar] [CrossRef]
- Segal, K.R.; Edaño, A.; Blando, L.; Pi-Sunyer, F.X. Comparison of thermic effects of constant and relative caloric loads in lean and obese men. Am. J. Clin. Nutr. 1990, 51, 14–21. [Google Scholar] [PubMed]
- Schutz, Y.; Bessard, T.; Jequier, E. Diet-induced thermogenesis measured over a whole day in obese and nonobese women. Am. J. Clin. Nutr. 1984, 40, 542–552. [Google Scholar] [PubMed]
- Hall, K.D.; Bemis, T.; Brychta, R.; Chen, K.Y.; Courville, A.; Crayner, E.J.; Goodwin, S.; Guo, J.; Howard, L.; Knuth, N.D.; et al. Calorie for Calorie, Dietary Fat Restriction Results in More Body Fat Loss than Carbohydrate Restriction in People with Obesity. Cell Metab. 2015, 22, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.E.; Lambert, M.H.; Montana, V.G.; Parks, D.J.; Blanchard, S.G.; Brown, P.J.; Sternbach, D.D.; Lehmann, J.M.; Wisely, G.B.; Willson, T.M.; et al. Molecular recognition of fatty acids by peroxisome proliferator-activated receptors. Mol. Cell 1999, 3, 397–403. [Google Scholar] [CrossRef]
- Rezai-Zadeh, K.; Yu, S.; Jiang, Y.; Laque, A.; Schwartzenburg, C.; Morrison, C.D.; Derbenev, A.V.; Zsombok, A.; Münzberg, H. Leptin receptor neurons in the dorsomedial hypothalamus are key regulators of energy expenditure and body weight, but not food intake. Mol. Metab. 2014, 3, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.; Spiers, J.P.; Crowley, V.; Williams, E.; Lithander, F.E. Postprandial adiponectin and gelatinase response to a high-fat versus an isoenergetic low-fat meal in lean, healthy men. Nutrition 2015, 31, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Bueno, A.A.; Oyama, L.M.; de Oliveira, C.; Pisani, L.P.; Ribeiro, E.B.; Silveira, V.L.; Oller do Nascimento, C.M. Effects of different fatty acids and dietary lipids on adiponectin gene expression in 3T3-L1 cells and C57BL/6J mice adipose tissue. Pflüg. Arch. 2008, 455, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Finucane, O.M.; Lyons, C.L.; Murphy, A.M.; Reynolds, C.M.; Klinger, R.; Healy, N.P.; Cooke, A.A.; Coll, R.C.; McAllan, L.; Nilaweera, K.N.; et al. Monounsaturated fatty acid-enriched high-fat diets impede adipose NLRP3 inflammasome-mediated IL-1beta secretion and insulin resistance despite obesity. Diabetes 2015, 64, 2116–2128. [Google Scholar] [CrossRef] [PubMed]
- Bjermo, H.; Iggman, D.; Kullberg, J.; Dahlman, I.; Johansson, L.; Persson, L.; Berglund, J.; Pulkki, K.; Basu, S.; Uusitupa, M.; et al. Effects of n-6 PUFAs compared with SFAs on liver fat, lipoproteins, and inflammation in abdominal obesity: A randomized controlled trial. Am. J. Clin. Nutr. 2012, 95, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Spady, D.K.; Kearney, D.M.; Hobbs, H.H. Polyunsaturated fatty acids up-regulate hepatic scavenger receptor B1 (SR-BI) expression and HDL cholesteryl ester uptake in the hamster. J. Lipid Res. 1999, 40, 1384–1394. [Google Scholar] [PubMed]
- Lopez, S.; Bermudez, B.; Ortega, A.; Varela, L.M.; Pacheco, Y.M.; Villar, J.; Abia, R.; Muriana, F.J. Effects of meals rich in either monounsaturated or saturated fat on lipid concentrations and on insulin secretion and action in subjects with high fasting triglyceride concentrations. Am. J. Clin. Nutr. 2011, 93, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Kruse, M.; von Loeffelholz, C.; Hoffmann, D.; Pohlmann, A.; Seltmann, A.C.; Osterhoff, M.; Hornemann, S.; Pivovarova, O.; Rohn, S.; Jahreis, G.; et al. Dietary rapeseed/canola-oil supplementation reduces serum lipids and liver enzymes and alters postprandial inflammatory responses in adipose tissue compared to olive-oil supplementation in obese men. Mol. Nutr. Food Res. 2015, 59, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Han, L.; Zhu, L.; Yu, Y. Free fatty acids, not triglycerides, are associated with non-alcoholic liver injury progression in high fat diet induced obese rats. Lipids Health Dis. 2016, 15, 27. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, A.M.; Brown, J.L.; Santiago, C.A.; Aad, P.Y.; Spicer, L.J.; Spicer, M.T. High-fat diets promote insulin resistance through cytokine gene expression in growing female rats. J. Nutr. Biochem. 2008, 19, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Van Hees, A.M.; Jocken, J.W.; Essers, Y.; Roche, H.M.; Saris, W.H.; Blaak, E.E. Adipose triglyceride lipase and hormone-sensitive lipase protein expression in subcutaneous adipose tissue is decreased after an isoenergetic low-fat high-complex carbohydrate diet in the metabolic syndrome. Metabolism 2012, 61, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health implications of high dietary omega-6 polyunsaturated Fatty acids. J. Nutr. Metab. 2012, 2012, 539426. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Enos, R.T.; Velázquez, K.T.; McClellan, J.L.; Cranford, T.L.; Walla, M.D.; Murphy, E.A. Reducing the dietary omega-6:omega-3 utilizing alpha-linolenic acid; not a sufficient therapy for attenuating high-fat-diet-induced obesity development nor related detrimental metabolic and adipose tissue inflammatory outcomes. PLoS ONE 2014, 9, e94897. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component (g/100 g of the Diet) | C | OB-M5 | OB-M15 | OB-M20 |
|---|---|---|---|---|
| Casein | 14 | 14 | 14 | 14 |
| Corn starch | 61.07 | 61.07 | 51.07 | 46.07 |
| Sucrose | 10 | 10 | 10 | 10 |
| Cellulose | 5 | 5 | 5 | 5 |
| Soybean oil | 5 | 0 | 0 | 0 |
| Experimental oil | 0 | 5 | 15 | 20 |
| AIN-93 Mineral mix † | 3.5 | 3.5 | 3.5 | 3.5 |
| AIN-93 Vitamin mix ‡ | 1 | 1 | 1 | 1 |
| L-cysteine | 0.18 | 0.18 | 0.18 | 0.18 |
| Choline bitartrate | 0.25 | 0.25 | 0.25 | 0.25 |
| Tert-butylhydroquinone | 0.001 | 0.001 | 0.001 | 0.001 |
| Calorie/g | ||||
| CHO% of energy | ||||
| Protein% of energy | ||||
| Fat% of energy | 11.7 | 11.7 | 31 | 39.1 |
| Fatty Acid % | Soybean Oil | Experimental Mix Oil |
|---|---|---|
| C14-0S | 0.12 | 0.03 |
| C16-0S | 11.25 | 5.76 |
| C18-0S | 3.48 | 1.99 |
| C16-1n7 | 0.11 | 0.24 |
| C18-1n9 | 23.46 | 54.11 |
| C20-1n9 | 0.13 | 0.97 |
| C18-2n6 | 54.87 | 26.32 |
| C20-2n6 | 0.05 | 2.52 |
| C18-3n3 | 6.52 | 7.95 |
| C20-5n3 | 0.02 | 0.08 |
| C22-6n3 | 0 | 0.03 |
| Total SFA | 14.84 | 7.78 |
| Total MUFA | 23.70 | 55.32 |
| Total PUFA | 61.46 | 36.90 |
| P/S ratio | 4.1 | 5.0 |
| S/M/P proportion | 1:1.6:4.1 | 1:7.1:4.7 |
| n-6/n-3 ratio | 8.39 | 3.58 |
| Variables | C | OB-M5 | OB-M15 | OB-M20 |
|---|---|---|---|---|
| Initial body weight (g) | 135.8 ± 1.5 b | 145.7 ± 5.1 a | 146.1 ± 3.5 a | 146.0 ± 3.8 a |
| Final body weight (g) | 153.5 ± 4.4 | 153.2 ± 6.1 | 161.0 ± 4.0 | 164.5 ± 2.3 |
| Liver weight (g) | 5.1 ± 0.3 | 5.5 ± 0.3 | 5.4 ± 0.2 | 5.4 ± 0.2 |
| Epididymal fat weight (g) | 2.7 ± 0.2 | 2.9 ± 0.3 | 3.1 ± 0.2 | 2.8 ± 0.2 |
| Retroperitoneal fat weight (g) | 0.9 ± 0.0 | 1.0 ± 0.1 | 0.9 ± 0.0 | 1.0 ± 0.0 |
| Weight gain (g) | 16.2 ± 3.1 a | 10.6 ± 4.3 b | 14.9 ± 8.1 a | 18.3 ± 1.0 a |
| Food intake (g/day) | 8.6 ± 0.3 a | 8.9 ± 0.2 a | 7.6 ± 0.1 b | 7.5 ± 0.2 b |
| Energy intake (kcal/day) | 33.0 ± 1.0 | 34.3 ± 0.8 | 33.2 ± 0.6 | 34.6 ± 0.9 |
| Feed efficiency (%) | 0.20 ± 0.04 | 0.10 ± 0.04 | 0.10 ± 0.05 | 0.10 ± 0.01 |
| Variables | C | OB-M5 | OB-M15 | OB-M20 |
|---|---|---|---|---|
| Triglycerides (mg/dL) | 111 ± 9.6 | 122 ± 11.4 | 109 ± 9.3 | 101 ± 5.4 |
| Cholesterol (mg/dL) | 74.2 ± 3.4 ab | 83.9 ± 6.3 a | 67.4 ± 2.7 bc | 63.0 ± 2.3 c |
| HDL-C (mg/dL) | 54.4 ± 2.5 b | 63.9 ± 5.0 a | 52.7 ± 2.3 b | 50.2 ± 2.4 b |
| LDL-C (mg/dL) | 4.8 ± 0.5 a | 4.3 ± 0.3 ab | 3.1 ± 0.2 c | 3.4 ± 0.2 bc |
| TC/HDL-C | 1.37 ± 0.02 a | 1.31 ± 0.01 ab | 1.30 ± 0.02 b | 1.28 ± 0.02 b |
| Glucose (mg/dL) | 250 ± 29.8 | 277 ± 23.9 | 276 ± 18.5 | 251 ± 20.6 |
| FFA (mmol/L) | 1.6 ± 0.1 | 1.8 ± 0.2 | 1.5 ± 0.1 | 1.5 ± 0.1 |
| Insulin (μg/L) | 1.4 ± 0.3 b | 1.8 ± 0.2 ab | 2.0 ± 0.1 a | 2.2 ± 0.2 a |
| Leptin (ng/mL) | 870 ± 574 | 1153 ± 562 | 1243 ± 676 | 835 ± 571 |
| Adiponectin (ng/mL) | 1.2 ± 0.2 | 1.5 ± 0.2 | 1.7 ± 0.2 | 1.5 ± 0.2 |
| Variables | C | OB-M5 | OB-M15 | OB-M20 |
|---|---|---|---|---|
| Liver tissue | ||||
| Liver TG (mg/g liver) | 12.6 ± 0.1 | 12.7 ± 0.2 | 12.8 ± 0.2 | 13.6 ± 0.9 |
| Liver TC (mg/g liver) | 10.4 ± 1.0 | 10.4 ± 0.9 | 10.7 ± 0.6 | 10.9 ± 1.2 |
| Liver FFA (mmol/g liver) | 1.79 ± 0.03 d | 1.87 ± 0.03 c | 1.97 ± 0.02 b | 2.07 ± 0.03 a |
| FAS (nmole/min/mg protein) | 2.51 ± 0.53 b | 4.99 ± 0.86 a | 6.96 ± 0.93 a | 5.64 ± 0.77 a |
| ACC (nmole/min/mg protein) | 2.85 ± 0.65 b | 5.13 ± 0.78 ab | 8.10 ± 2.05 a | 3.40 ± 0.84 b |
| ACO (nmole/min/mg protein) | 0.41 ± 0.01 c | 1.05 ± 0.19 b | 1.58 ± 0.26 a | 1.13 ± 0.26 b |
| CPT-1 (nmole/min/mg protein) | 0.56 ± 0.16 | 0.44 ± 0.19 | 0.50 ± 0.03 | 0.44 ± 0.01 |
| LPL activities in adipose tissues | ||||
| Epididymal fat (mU/mL) | 7.7 ± 1.2 | 7.5 ± 0.7 | 7.6 ± 0.8 | 10.1 ± 0.9 |
| Retroperitoneal fat (mU/mL) | 11.0 ± 1.1 | 10.3 ± 1.0 | 11.0 ± 1.5 | 10.5 ± 1.5 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.-C.; Lin, S.-H.; Chang, J.-S.; Chien, Y.-W. High Fat Diet with a High Monounsaturated Fatty Acid and Polyunsaturated/Saturated Fatty Acid Ratio Suppresses Body Fat Accumulation and Weight Gain in Obese Hamsters. Nutrients 2017, 9, 1148. https://doi.org/10.3390/nu9101148
Yang S-C, Lin S-H, Chang J-S, Chien Y-W. High Fat Diet with a High Monounsaturated Fatty Acid and Polyunsaturated/Saturated Fatty Acid Ratio Suppresses Body Fat Accumulation and Weight Gain in Obese Hamsters. Nutrients. 2017; 9(10):1148. https://doi.org/10.3390/nu9101148
Chicago/Turabian StyleYang, Suh-Ching, Shyh-Hsiang Lin, Jung-Su Chang, and Yi-Wen Chien. 2017. "High Fat Diet with a High Monounsaturated Fatty Acid and Polyunsaturated/Saturated Fatty Acid Ratio Suppresses Body Fat Accumulation and Weight Gain in Obese Hamsters" Nutrients 9, no. 10: 1148. https://doi.org/10.3390/nu9101148
APA StyleYang, S. -C., Lin, S. -H., Chang, J. -S., & Chien, Y. -W. (2017). High Fat Diet with a High Monounsaturated Fatty Acid and Polyunsaturated/Saturated Fatty Acid Ratio Suppresses Body Fat Accumulation and Weight Gain in Obese Hamsters. Nutrients, 9(10), 1148. https://doi.org/10.3390/nu9101148