Probiotic Modulation of Innate Cell Pathogen Sensing and Signaling Events


Abstract
:1. Introduction
2. Probiotic Modulation of Intestinal Epithelial Cells
2.1. Modulation of Barrier Function
2.2. Modulation of Mucosal Immunity
2.3. Probiotic Modulation of IEC TLR Expression Regulates Mucosal Intestinal Immunity and Tolerance
3. Probiotic Modulation of DCs
3.1. Activation of DCs
3.2. Probiotic Modulation of NK Cells via Cross-Talk with DCs
4. Probiotic Modulation of Neutrophils
5. Probiotic Modulation of Macrophage Signaling Pathways Influences Cytokine Production
5.1. Modulation of Pro-Inflammatory Cytokines
5.2. Inflammasomes
5.3. Modulation of the IL-12/IL-10 Balance
5.4. Lipid Rafts—CD14, TLR2 and TLR4
5.5. Regulation by miRNA
5.6. Probiotics as Vehicles of Endotoxin Tolerisation
6. Secreted Proteins, Lipids, Polysaccharides and Metabolites
7. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- FAO/WHO. Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria; World Health Organization: Basel, Switzerland, 2001. [Google Scholar]
- Borcher, A.T.; Selmi, C.; Meyers, F.J.; Keen, C.L.; Gershwin, M.E. Probiotics and immunity. J. Gastroenterol. 2009, 44, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A.D. Probiotics, prebiotics and immunomodulation of gut mucosal defences: Homeostasis and immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef] [PubMed]
- Saxelin, M.; Tynkkynen, S.; Mattila-Sandholm, T.; M de Vos, W. Probiotic and other functional microbes: From markets to mechanisms. Curr. Opin. Biotechnol. 2005, 16, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Atris, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Vastano, V.; Pagano, A.; Fusco, A.; Merola, G.; Sacco, M.; Donnarumma, G. The Lactobacillus plantarum Eno A1 enolase is involved in immunostimulation of Caco-2 cells and in biofilm development. Adv. Exp. Med. Biol. 2016, 897, 33–44. [Google Scholar] [PubMed]
- Zyrek, A.A.; Cichon, C.; Helms, S.; Enders, C.; Sonnenborn, U.; Schmidt, M.A. Molecular mechanisms underlying the probiotic effects of Escherichia coli Nissle 1917 involve ZO-2 and PKCzeta redistribution resulting in tight junction and epithelial barrier repair. Cell Microbiol. 2007, 9, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Schlee, M.; Harder, J.; Koten, B.; Stange, E.F.; Wehkamp, J.; Fellermann, K. Probiotic lactobacilli and VSL#3 induce enterocyte β-defensin 2. Clin. Exp. Immunol. 2008, 151, 528–535. [Google Scholar] [PubMed]
- Liu, H.; Hou, C.; Wang, G.; Jia, H.; Yu, H.; Zeng, X.; Thacker, P.A.; Zhang, G.; Qiao, S. Lactobacillus reuteri I5007 modulates intestinal host defense peptide expression in the model of IPEC-J2 cells and neonatal piglets. Nutrients 2017, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Resta-Lenert, S.; Barrett, K.E. Live probiotics protect intestinal epithelial cells from the effects of infection with enteroinvasive Escherichia coli (EIEC). Gut 2003, 52, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kokkotou, E.G.; Mustafa, N.; Bhaskar, K.R.; Sougioultzis, S.; O’Brien, M.; Pothoulakis, C.; Kelly, C.P. Saccharomyces boulardii inhibits ERK1/2 mitogen-activated protein kinase activation both in vitro and in vivo and protects against Clostridium difficile toxin A-induced enteritis. J. Biol. Chem. 2006, 281, 24449–24454. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef] [PubMed]
- Segatto, O.; Anastasi, S.; Alemà, S. Regulation of epidermal growth factor receptor signalling by inducible feedback inhibitors. J. Cell Sci. 2011, 128, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Fanning, A.S.; Jameson, B.J.; Jesaitis, L.A.; Anderson, J.M. The tight junction protein ZO-1 establishes a link between the transmembrane protein occludin and the actin cytoskeleton. J. Biol. Chem. 1998, 273, 29745–29753. [Google Scholar] [CrossRef] [PubMed]
- Schulzke, J.D.; Ploeger, S.; Amasheh, M.; Fromm, A.; Zeissig, S.; Troeger, H.; Richter, J.; Bojarski, C.; Schumann, M.; Fromm, M. Epithelial Tight Junctions in Intestinal Inflammation. Ann. N. Y. Acad. Sci. 2009, 1165, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Orlando, A.; Linsalata, M.; Notarnicola, M.; Tutino, V.; Russo, F. Lactobacillus GG restoration of the gliadin induced epithelial barrier disruption: The role of cellular polyamines. BMC Microbiol. 2014, 14, 19. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.-J.M.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhu, Y.H.; Yang, J.C.; Yang, G.Y.; Zhou, D.; Wang, J.F. A Selected Lactobacillus rhamnosus Strain Promotes EGFR-Independent Akt Activation in an Enterotoxigenic Escherichia coli K88- Infected IPEC-J2 Cell Model. PLoS ONE 2015, 10, e0125717. [Google Scholar] [CrossRef] [PubMed]
- Dent, P.; Yacoub, A.; Fisher, P.B.; Hagan, M.P.; Gran, S. MAPK pathways in radiation responses. Oncogene 2003, 22, 5885–5896. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Zhao, D.-H.; Jiang, M. VSL#3 probiotics regulate the intestinal epithelial barrier in vivo and in vitro via the p38 and ERK signaling pathways. Int. J. Mol. Med. 2012, 29, 202–208. [Google Scholar] [PubMed]
- Tao, Y.; Drabik, K.A.; Waypa, T.S.; Musch, M.W.; Alverdy, J.C.; Schneewind, O.; Chang, E.B.; Petrof, E.O. Soluble factors from Lactobacillus GG activate MAPKs and induce cytoprotective heat shock proteins in intestinal epithelial cells. Am. J. Physiol. Cell Physiol. 2006, 290, C1018–C1030. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Liu, L.; Dempsey, P.J.; Tsai, Y.H.; Raines, E.W.; Wilson, C.L.; Cao, H.; Cao, Z.; Lui, L.; Polk, D.B. A Lactobacillus rhamnosus GG-derived soluble protein, p40, stimulates ligand release from intestinal epithelial cells to transactivate epidermal growth factor receptor. J. Biol. Chem. 2013, 288, 30742–30751. [Google Scholar] [CrossRef] [PubMed]
- Yoda, K.; Miyazawa, K.; Hosoda, M.K.; Hiramatsu, M.; Yan, F.; He, F. Lactobacillus GG-fermented milk prevents DSS-induced colitis and regulates intestinal epithelial homeostasis through activation of epidermal growth factor receptor. Eur. J. Nutr. 2014, 53, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Yoda, K.; He, F.; Miyazawa, K.; Hiramatsu, M.; Yan, F. Fermented milk containing Lactobacillus GG alleviated DSS-induced colitis in mice and activated epidermal growth factor receptor and Akt signaling in intestinal epithelial cells. Microb. Ecol. Health Dis. 2012, 23, 1651–2235. [Google Scholar]
- Tomosada, Y.; Villena, J.; Murata, K.; Chiba, E.; Shimazu, T.; Aso, H.; Iwabuchi, N.; Xiao, J.Z.; Saito, T.; Kitazawa, H. Immunoregulatory effect of bifidobacteria strains in porcine intestinal epithelial cells through modulation of ubiquitin-editing enzyme A20 expression. PLoS ONE 2013, 8, e59259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.W.; Kim, H.M.; Yang, K.M.; Kim, S.A.; Kim, S.K.; An, M.J.; Park, J.J.; Lee, S.K.; Kim, T.I.; Kim, W.H.; et al. Bifidobacterium lactis inhibits NF-kappaB in intestinal epithelial cells and prevents acute colitis and colitis-associated colon cancer in mice. Inflamm. Bowel Dis. 2010, 16, 1514–1525. [Google Scholar] [CrossRef] [PubMed]
- Finamore, A.; Roselli, M.; Imbinto, A.; Seeboth, J.; Oswald, I.P.; Mengheri, E. Lactobacillus amylovorus inhibits the TLR4 inflammatory signaling triggered by enterotoxigenic Escherichia coli via modulation of the negative regulators and involvement of TLR2 in intestinal Caco-2 Cells and pig explants. PLoS ONE 2014, 9, e94891. [Google Scholar] [CrossRef] [PubMed]
- Takanashi, N.; Tomosada, Y.; Villena, J.; Murata, K.; Takahashi, T.; Chiba, E.; Tohno, M.; Shimazu, T.; Aso, H.; Suda, Y.; et al. Advanced application of bovine intestinal epithelial cell line for evaluating regulatory effect of lactobacilli against heat-killed enterotoxigenic Escherichia coli-mediated inflammation. BMC Microbiol. 2013, 13, e54. [Google Scholar] [CrossRef] [PubMed]
- Mirpuri, J.; Sotnikov, I.; Myers, L.; Denning, T.L.; Yarovinsky, F.; Parkos, C.A. Lactobacillus rhamnosus (LGG) Regulates IL-10 signaling in the developing murine colon through upregulation of the IL-10R2 receptor subunit. PLoS ONE 2012, 7, e51955. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.W.; Myers, L.E.S.; Ray, L.; Song, S.C.; Nasr, T.R.; Berardinelli, A.J.; Kundu, K.; Murthy, N.; Hansen, J.M.; Neish, A.S. Lactobacillus rhamnosus blocks inflammatory signaling in vivo via reactive oxygen species generation. Free Radic. Biol. Med. 2009, 47, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Cao, H.; Cover, T.L.; Washington, M.K.; Shi, Y.; Liu, L.; Chaturvedi, R.; Peek, R.M., Jr.; Wilson, K.T.; Polk, D.B. Colon-specific delivery of a probiotic-derived soluble protein ameliorates intestinal inflammation in mice through an EGFR-dependent mechanism. J. Clin. Investig. 2011, 121, 2242–2253. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, T.; Villena, J.; Tohno, M.; Fujie, H.; Hosoya, S.; Shimosato, T.; Aso, H.; Suda, Y.; Kawai, Y.; Saito, T.; et al. Immunobiotic Lactobacillus jensenii elicits anti-inflammatory activity in porcine intestinal epithelial cells by modulating negative regulators of the Toll-like receptor signaling pathway. Infect. Immun. 2012, 80, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fatheree, N.Y.; Mangalat, N.; Rhoads, J.M. Lactobacillus reuteri strains reduce incidence and severity of experimental necrotizing enterocolitis via modulation of TLR4 and NF-κB signaling in the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 302, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Zanello, G.; Berri, M.; Dupont, J.; Sizaret, P.-Y.; D’Inca, R.; Salmon, H.; Meurens, F. Saccharomyces cerevisiae modulates immune gene expressions and inhibits ETEC-mediated ERK1/2 and p38 signaling pathways in intestinal epithelial cells. PLoS ONE 2011, 6, e18573. [Google Scholar] [CrossRef] [PubMed]
- Kaci, G.; Lakhdari, O.; Doré, J.; Dusko-Ehrlich, S.; Renault, P.; Blottiere, H.M.; Delorme, C. Inhibition of the NF-κB pathway in human intestinal epithelial cells by commensal streptococcus salivarius. Appl. Environ. Microbiol. 2011, 77, 4681–4684. [Google Scholar] [CrossRef] [PubMed]
- Petrof, E.O.; Kojima, K.; Ropeleski, M.J.; Musch, M.W.; Tao, Y.; De Simone, C.; Chang, E.B. Probiotics inhibit nuclear factor-kappa B and induce heat shock proteins in colonic epithelial cells through proteasome inhibition. Gastroenterology 2004, 127, 1474–1487. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Signaling to NF-κB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Toll-like receptors. Curr. Protoc. Immunol. 2015, 12–14. [Google Scholar]
- Manicassamy, S.; Pulendran, B. Modulation of adaptative immunity with Toll-like receptors. Semin. Immunol. 2009, 21, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Ben-Neriah, Y. Phosphorylation meets ubiquitination: The control of NF-κB activity. Ann. Rev. Immunol. 2000, 18, 621–663. [Google Scholar] [CrossRef] [PubMed]
- Moue, M.; Tohno, M.; Shimazu, T.; Kido, T.; Aso, H.; Saito, T.; Kitazawa, H. Toll-like receptor 4 and cytokine expression involved in functional immune response in an originally established porcine intestinal epitheliocyte cell line. BBA 2008, 1780, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Shibolet, O.; Podolsky, D.K. TLRs in the Gut. IV. Negative regulation of Toll-like receptors and intestinal homeostasis: Addition by subtraction. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, 1469–1473. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Gulen, M.F.; Qin, J.; Yao, J.; Bulek, K.; Kish, D.; Altuntas, C.Z.; Wakd, D.; Ma, C.; Zhou, H.; et al. The Toll-interleukin-1 receptor member SIGIRR regulates colonic epithelial homeostasis, inflammation, and tumorigenesis. Immunity 2007, 26, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Boone, D.L.; Turer, E.E.; Lee, E.G.; Ahmad, R.C.; Wheeler, M.T.; Tsui, C.; Hurley, P.; Chien, M.; Chai, S.; Hitotsumatsu, O.; et al. The ubiquitin-modifying enzyme A20 is required for termination of Toll-like receptor responses. Nat. Immunol. 2004, 5, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Lamothe, B.; Besse, A.; Campos, A.D.; Webster, W.K.; Wu, H.; Darnay, B.G. Site-specific Lys-63-linked tumor necrosis factor receptor-associated factor 6 auto-ubiquitination is a critical determinant of I kappa B kinase activation. J. Biol. Chem. 2007, 282, 4102–4112. [Google Scholar] [CrossRef] [PubMed]
- Donato, K.A.; Gareau, M.G.; Wang, Y.J.; Sherman, P.M. Lactobacillus rhamnosus GG attenuates interferon-c and tumour necrosis factor-a-induced barrier dysfunction and pro-inflammatory signaling. Microbiology 2010, 156, 3288–3297. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Wu, H.; Collier-Hyams, L.S.; Hansen, J.M.; Li, T.; Yamoah, K.; Pan, Z.Q.; Jones, D.P.; Neish, A.S. Commensal bacteria modulate cullin-dependent signaling via generation of reactive oxygen species. EMBO J. 2007, 26, 4457–4466. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, A.P. The IL-10/STAT3-mediated anti-inflammatory response: Recent developments and future challenges. Brief. Funct. Genom. 2013, 12, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Sato, A. Intestinal epithelial barrier and mucosal immunity. Cell. Mol. Life Sci. 2005, 62, 1333–1338. [Google Scholar]
- Mabbott, N.A.; Donaldson, D.S.; Ohno, H.; Williams, I.R.; Mahajan, A. Microfold (M) cells: Important immunosurveillance posts in the intestinal epithelium. Mucosal Immunol. 2013, 6, 666–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacPherson, G.; Milling, S.; Yrlid, U.; Cousins, L.; Turnbull, E.; Huang, A.P. Uptake of antigens from the intestine by Dendritic Cells. Ann. N. Y. Acad. Sci. 2004, 1029, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Melmed, G.; Thomas, L.S.; Lee, N.; Tesfay, S.Y.; Lukasek, K.; Michelsen, K.S.; Zhou, Y.; Hu, B.; Arditi, M.; Abreu, M.T. Human intestinal epithelial cells are broadly unresponsive to Toll-like receptor 2-dependent bacterial ligands: Implications for host-microbial interactions in the gut. J. Immunol. 2003, 170, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Mo, J.H.; Katakura, K.; Alkalay, I.; Rucker, A.N.; Liu, Y.T.; Lee, H.K.; Shen, C.; Cojocaru, G.; Shenouda, S.; et al. Maintenance of colonic homeostasis by distinctive apical TLR9 signalling in intestinal epithelial cells. Nat. Cell Biol. 2006, 8, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Huang, J.; Tang, W.; Chen, B.; Cai, W. Effects of probiotics, probiotic DNA and the CpG oligodeoxynucleotides on ovalbumin-sensitized Brown- Norway rats via TLR9/NF-κB pathway. FEMS Immunol. Med. Microbiol. 2012, 66, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Ghadimi, D.; Vrese, M.; Heller, K.J.; Schrezenmeir, J. Effect of natural commensal- origin DNA on toll-like receptor 9 (TLR9) signaling cascade, chemokine IL-8 expression, and barrier integritiy of polarized intestinal epithelial cells. Inflamm. Bowel Dis. 2010, 16, 410–427. [Google Scholar] [CrossRef] [PubMed]
- Bouladoux, N.; Hall, J.A.; Grainger, J.R.; dos Santos, L.M.; Kann, M.G.; Nagarajan, V.; Verthelyi, D.; Belkaid, Y. Regulatory role of suppressive motifs from commensal DNA. Mucosal Immunol. 2012, 5, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Iliev, I.D.; Spadoni, I.; Mileti, E.; Matteoli, G.; Sonzogni, A.; Sampietro, G.M.; Foschi, D.; Caprioli, F.; Viale, G.; Rescigno, M. Human intestinal epithelial cells promote the differentiation of tolerogenic dendritic cells. Gut 2009, 58, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Coombes, J.L.; Siddiqui, K.R.; Arancibia-Cárcamo, C.V.; Hall, J.; Sun, C.M.; Belkaid, Y.; Powrie, F. A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-beta and retinoic acid-dependent mechanism. J. Exp. Med. 2007, 204, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.G.; Kayama, H.; Ueda, Y.; Takahashi, T.; Asahara, T.; Tsuji, H.; Tsuji, N.M.; Kiyono, H.; Ma, J.S.; Kusu, T.; et al. Probiotic Bifidobacterium breve induces IL-10-producing Tr1 cells in the colon. PLoS Pathog. 2012, 8, e1002714. [Google Scholar] [CrossRef] [PubMed]
- Macho Fernandez, E.; Valenti, V.; Rockel, C.; Hermann, C.; Pot, B.; Boneca, I.G.; Grangette, C. Anti-inflammatory capacity of selected lactobacilli in experimental colitis is driven by NOD2-mediated recognition of a specific peptidoglycan-derived muropeptide. Gut 2011, 60, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.-K.; Lee, C.-G.; So, J.-S.; Chae, C.S.; Hwang, J.S.; Sahoo, A.; Nam, J.H.; Rhee, J.H.; Hwang, K.C.; Im, S.H. Generation of regulatory dendritic cells and CD4+Foxp3+ T cells by probiotics administration suppresses immune disorders. Proc. Natl. Acad. Sci. USA 2010, 107, 2159–2164. [Google Scholar] [CrossRef] [PubMed]
- Zeuthen, L.H.; Fink, L.N.; Frøkiær, H. Toll-like receptor 2 and nucleotide-binding oligomerization domain-2 play divergent roles in the recognition of gut-derived lactobacilli and bifidobacteria in dendritic cells. Immunology 2008, 24, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Muñoz-Quezada, S.; Gomez-Llorente, C.; Matencio, E.; Bernal, M.J.; Romero, F.; Gil, A. Cell-free culture supernatant of Bifidobacterium breve CNCM I-4035 decreases pro-inflammatory cytokines in human dendritic cells challenged with Salmonella typhi through TLR activation. PLoS ONE 2013, 8, e59370. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ghosh, S. Negative regulation of toll-like receptor-mediated signalling by Tollip. J. Biol. Chem. 2002, 277, 7059–7065. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Muñoz-Quezada, S.; Gomez-Llorente, C.; Matencio, E.; Bernal, M.J.; Romero, F.; Gil, A. Human Intestinal Dendritic Cells Decrease Cytokine Release against Salmonella Infection in the Presence of Lactobacillus paracasei upon TLR Activation. PLoS ONE 2012, 7, e43197. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Muñoz-Quezada, S.; Gomez-Llorente, C.; Romero, F.; Gil, A. Lactobacillus rhamnosus and its cell-free culture supernatant differentially modulate inflammatory biomarkers in Escherichia coli-challenged human dendritic cells. Br. J. Nutr. 2014, 111, 1727–1737. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H.R.; Frokiaer, H.; Pestka, J.J. Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells. J. Immunol. 2002, 168, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Di Santo, J.P.; Vosshenrich, C.A.; Satoh-Takayama, N. A ‘natural’ way to provide innate mucosal immunity. Curr. Opin. Immunol. 2010, 22, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Rowland, I.; Tuohy, K.M.; Thomas, L.V.; Yaqoob, P. Selective effects of Lactobacillus casei Shirota on T-cell activation, natural killer cell activity and cytokine production. Clin. Exp. Immunol. 2010, 161, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Fink, L.N.; Zeuthen, L.H.; Christensen, H.R.; Morandi, B.; Frokiaer, H.; Ferlazzo, G. Distinct gut-derived lactic acid bacteria elicit divergent dendritic cell-mediated NK cell responses. Int. Immunol. 2007, 19, 1319–1327. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, S.; Wakita, D.; Sato, T.; Mitamura, R.; Izumo, T.; Shibata, H.; Kiso, Y.; Chamoto, K.; Togashi, Y.; Kitamura, H.; Nishimura, T. Essential role of Toll-like receptors for dendritic cell and NK1.1(+) cell-dependent activation of type 1 immunity by Lactobacillus pentosus strain S-PT84. Immunol. Lett. 2008, 120, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Suzuki, T.; Shimada, S.-I.; Shida, K.; Nanno, M.; Okumura, K. Interleukin-12 is involved in the enhancement of human natural killer cell activity by Lactobacillus casei Shirota. Clin. Exp. Immunol. 2006, 146, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Okumura, K. Effects of a fermented milk drink containing Lactobacillus casei strain Shirota on the human NK-cell activity. J. Nutr. 2007, 137, 791S–793S. [Google Scholar] [PubMed]
- Carroll, I.M.; Andrus, J.M.; Bruno-Barcena, J.M.; Klaenhammer, T.R.; Hassan, H.M.; Threadgill, D.S. Anti-inflammatory properties of Lactobacillus gasseri expressing manganedse superoxide dismutase using the interleukin 10-deficient mouse model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G729–G738. [Google Scholar] [CrossRef] [PubMed]
- Roselli, M.; Finamore, A.; Britti, M.S.; Mengheri, E. Probiotic bacteria Bifidobacterium animalis MB5 and Lactobacillus rhamnosus GG protect intestinal Caco-2 cells from the inflammation-associated response induced by enterotoxigenic Escherichia coli K88. Br. J. Nutr. 2006, 95, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, K.; Gill, H.S.; Chandra, R.K. Enhancement of natural immune function by dietary consumption of Bifidobacterium lactis (HN019). Eur. J. Clin. Nutr. 2000, 54, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Souza, E.L.; Elian, S.D.; Paula, L.M.; Garcia, C.C.; Viera, A.T.; Teixeira, M.M.; Arantes, R.M.; Nicoli, J.R.; Martins, F.S. Escherichia coli strain Nissle 1917 ameliorates experimental colitis by modulating intestinal permeability, the inflammatory response and clinical signs in a faecal transplantation model. J. Med. Microbiol. 2016, 65, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; van Bergenhenegouwen, J.; van de Kant, H.J.G.; Folkerts, G.; Garssen, J.; Vos, A.P.; Morgan, M.E.; Kraneveld, A.D. Specific probiotic dietary supplementation leads to different effects during remission and relapse in murine chronic colitis. Benef. Microbes 2016, 7, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Elian, S.D.; Souza, E.L.; Vieira, A.T.; Teixeira, M.M.; Arantes, R.M.; Nicoli, J.R.; Martins, F.S. Bifidobacterium longum subsp. Infantis BB-02 attenuates acute murine experimental model of inflammatory bowel disease. Benef. Microbes 2015, 6, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Boonma, P.; Spinler, J.K.; Venable, S.F.; Versalovic, J.; Tumwasorn, S. Lactobacillus rhamnosus L34 and Lactobacillus casei L39 suppress Clostridium difficile-induced IL-8 production by colonic epithelial cells. BMC Microbiol. 2014, 14, 177. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Hicks, C.; Li, Y.; Su, J.; Shiloach, J.; Kaufman, J.B.; Fitz, Y.; Eichacker, P.Q.; Cui, X. Purified cell wall from the probiotic bacterium Lactobacillus gasseri activates systemic inflammation and, at higher doses, produces lethality in a rat model. Crit. Care 2014, 18, R140. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, Y.J.; Lee, S.H.; Yu, J.; Jeong, S.K.; Hong, S.J. Effects of Lactobacillus rhamnosus on allergic march model by suppressing Th2, Th17, and TSLP responses via CD4(+)CD25(+)Foxp3(+) Tregs. Clin. Immunol. 2014, 153, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Vong, L.; Lorentz, R.J.; Assa, A.; Glogauer, M.; Sherman, P.M. Probiotic Lactobacillus rhamnosus inhibits the formation of neutrophil extracellular traps. J. Immunol. 2014, 192, 1870–1877. [Google Scholar] [CrossRef] [PubMed]
- Hidemura, A.; Saito, H.; Fukatsu, K.; Matsuda, T.; Kitayama, J.; Ikeda, S.; Kang, W.; Nagawa, H. Oral administration of Bifidobacterium longum culture condensate in a diet-restricted murine peritonitis model enhances polymorphonuclear neutrophil recruitment into the local inflammatory site. Nutrition 2003, 19, 270–274. [Google Scholar] [CrossRef]
- Lammers, K.M.; Vergopoulos, A.; Babel, N.; Gionchetti, P.; Rizzello, F.; Morselli, C.; Caramelli, E.; Fiorentino, M.; d’Errico, A.; et al. Probiotic therapy in the prevention of pouchitis onset: Decreased interleukin-1beta, interleukin-8, and interferon-gamma gene expression. Inflamm. Bowel Dis. 2005, 11, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Peral, M.C.; Rachid, M.M.; Gobbato, N.M.; Huaman Martinez, M.A.; Valdez, J.C. Interleukin-8 production by polymorphonuclear leukocytes from patients with chronic infected leg ulcers treated with Lactobacillus plantarum. Clin. Microbiol. Infect. 2010, 16, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Shinnoh, M.; Horinaka, M.; Yasuda, T.; Yoshikawa, S.; Morita, M.; Yamada, T.; Miki, T.; Sakai, T. Clostridium butyricum MIYAIRI 588 shows antitumour effects by enhancing the release of TRAIL from neutrophils through MMP-8. Int. J. Oncol. 2013, 42, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.O.; Sheikh, H.I.; Ha, S.D.; Martins, A.; Reid, G. G-CSF-mediated inhibition of JNK is a key mechanism for Lactobacillus rhamnosus-induced suppression of TNF production in macrophages. Cell. Microbiol. 2006, 8, 1958–1971. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Paek, N.S.; Kwon, O.S.; Hahm, K.B. Anti-inflammatory actions of probiotics through activating suppressor of cytokine signaling (SOCS) expression and signaling in Helicobacter pylori infection: A novel mechanism. J. Gastroenterol. Hepatol. 2010, 25, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Latvala, S.; Miettinen, M.; Kekkonen, R.A.; Korpela, R.; Julkunen, I. Lactobacillus rhamnosus GG and Streptococcus thermophiles induce suppressor of cytokine signalling 3 (SOCS3) gene expression directly and indirectly via interleukin-10 in human primary macrophages. Clin. Exp. Immunol. 2011, 165, 94–103. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, J.J.; Murray, P.J. Cytokine signaling modules in inflammatory responses. Immunity 2008, 28, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Nishio, H.; Tanigawa, T.; Yamagami, H.; Okazaki, H.; Watanabe, K.; Tominaga, K.; Fujiwara, Y.; Oshitani, N.; Asahara, T.; et al. Probiotic Lactobacillus casei strain Shirota prevents indomethacin-induced small intestinal injury: Involvement of lactic acid. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Sougioultzis, S.; Simeonidis, S.; Bhaskar, K.R.; Chen, X.; Anton, P.M.; Keates, S.; Pothoulakis, C.; Kelly, C.P. Saccharomyces boulardii produces a soluble anti-inflammatory factor that inhibits NF-κB-mediated IL-8 gene expression. Biochem. Biophys. Res. Commun. 2006, 343, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Ménard, S.; Candalh, C.; Bambou, J.C.; Terpend, K.; Cerf-Bensussan, N.; Heyman, M. Lactic acid bacteria secrete metabolites retaining anti-inflammatory properties after intestinal transport. Gut 2004, 53, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Kim, N.R.; Gim, M.G.; Lee, J.M.; Lee, S.Y.; Ko, M.Y. Lipoteichoic acid isolated from Lactobacillus plantarum inhibits lipopolysaccharide-induced TNF-a production in THP-1 cells and endotoxin shock in mice. J. Immunol. 2008, 180, 2553–2561. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Lee, S.Y.; Kim, N.R.; Ko, M.Y.; Lee, J.M.; Yi, T.H.; Chung, S.K.; Chung, D.K. Inhibitory effects of Lactobacillus plantarum lipoteichoic acid (LTA) on Staphylococcus aureus LTA-induced tumor necrosis factor-α production. J. Microbiol. Biotechnol. 2008, 18, 1191–1196. [Google Scholar] [PubMed]
- Kim, H.G.; Lee, S.Y.; Kim, N.R.; Lee, H.Y.; Ko, M.Y.; Jung, B.J.; Kim, C.M.; Lee, J.M.; Park, J.H.; Han, S.H.; et al. Lactobacillus plantarum lipoteichoic acid down-regulated Shigella flexneri peptidoglycan-induced inflammation. Mol. Immunol. 2011, 48, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.P.; Thibodeaux, C.H.; Peña, J.A.; Ferry, G.D.; Versalovic, J. Probiotic Lactobacillus reuteri suppress proinflammatory cytokines via c-Jun. Inflamm. Bowel Dis. 2008, 14, 1068–1083. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Tsuzuki, Y.; Hokari, R.; Komoto, S.; Kurihara, C.; Kawaguchi, A.; Nagao, S.; Miura, S. Anti-inflammatory effects of the genus Bifidobacterium on macrophages by modification of phospho-IkB and SOCS gene expression. Int. J. Exp. Pathol. 2009, 90, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.J.; Colquhoun, P.; Reid, G.; Kim, S.O. Reduced expression of basal and probiotic-inducible G-CSF in intestinal mononuclear cells is associated with inflammatory bowel disease. Inflamm. Bowel Dis. 2009, 15, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.J.; Spanton, S.; Sheikh, H.I.; Kim, S.O. The anti-inflammatory role of granulocyte colony-stimulating factor in macrophage– dendritic cell crosstalk after Lactobacillus rhamnosus GR-1 exposure. J. Leukoc. Biol. 2011, 89, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Habil, N.; Al-Murrani, W.; Beal, J.; Foey, A.D. Probiotic bacterial strains differentially modulate macrophage cytokine production in a strain-dependent and cell subset-specific manner. Benef. Microbes 2011, 2, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Habil, N.; Beal, J.; Foey, A.D. Lactobacillus casei strain Shirota selectively modulates macrophage subset cytokine production. Int. J. Probiot. Prebiot. 2012, 7, 1–12. [Google Scholar]
- Saemann, M.D.; Bohmig, G.A.; Osterreicher, C.H.; Burtscher, H.; Parolini, O.; Diakos, C.; Stockl, J.; Horl, W.H.; Zlabinger, G.J. Anti-inflammatory effects of sodium butyrate on human monocytes: Potent inhibition of IL-12 and up-regulation of IL-10 production. FASEB J. 2000, 14, 2380–2382. [Google Scholar] [CrossRef] [PubMed]
- Foey, A.D. Butyrate regulation of distinct macrophage subsets: Opposing effects on M1 and M2 macrophages. Int. J. Probiot. Prebiot. 2011, 6, 147–158. [Google Scholar]
- Kaji, R.; Kiyoshima-Shibata, J.; Nagaoka, M.; Nanno, M.; Shida, K. Bacterial teichoic acids reverse predominant IL-12 production induced by certain Lactobacillus Strains into predominant IL-10 production via TLR2-dependent ERK activation in macrophages. J. Immunol. 2010, 184, 3505–3513. [Google Scholar] [CrossRef] [PubMed]
- Lammers, K.M.; Brigidi, P.; Vitali, B.; Gionchetti, P.; Rizzello, F.; Caramelli, E.; Matteuzzi, D.; Campieri, M. Immunomodulatory effects of probiotic bacteria DNA: IL-1 and IL-10 response in human peripheral blood mononuclear cells. FEMS Immunol. Med. Microbiol. 2003, 38, 165–172. [Google Scholar] [CrossRef]
- Sun, K.-Y.; Xu, D.-H.; Xie, C.; Plummer, S.; Tang, J.; Yang, X.F.; Ji, X.H. Lactobacillus paracasei modulates LPS-induced inflammatory cytokine release by monocyte-macrophages via the up-regulation of negative regulators of NF-kappaB signaling in a TLR2-dependent manner. Cytokine 2017, 92, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Foey, A.D. Macrophage polarization: A collaboration of differentiation, activation and pre-programming? J. Clin. Cell Immunol. 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Merry, R.; Belfield, L.; McArdle, P.; McLennan, A.; Crean, S.J.; Foey, A. Oral health and pathology: A macrophage account. Br. J. Oral Maxillofac. Surg. 2012, 50, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Elinav, E.; Strowig, T.; Henao-Mejia, J.; Flavell, R.A. Regulation of the antimicrobial response by NLR proteins. Immunity 2011, 34, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-1beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Faustin, B.; Lartigue, L.; Bruey, J.M.; Luciano, F.; Sergienko, E.; Bailly-Maitre, B.; Volkmann, N.; Hanein, D.; Rouiller, I.; Reed, J.C. Reconstituted NALP1 inflammasome reveals two-step mechanism of caspase-1 activation. Mol. Cell 2007, 25, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.C.; Ali, S.R.; McGillivray, S.; Tseng, P.H.; Mariathasan, S.; Humke, E.W.; Eckmann, L.; Powell, J.J.; Nizet, V.; Dixit, V.M.; et al. A NOD2-NALP1 complex mediates caspase-1-dependent IL-1beta secretion in response to Bacillus anthracis infection and muramyl dipeptide. Proc. Natl. Acad. Sci. USA 2008, 107, 21635–21640. [Google Scholar]
- Guarda, G.; Braun, M.; Staehil, F.; Tardivel, A.; Mattmann, C.; Forster, I.; Farlik, M.; Decker, T.; Du Pasquier, R.A.; Romero, P.; et al. Type I interferon inhibits interleukin-1 production and inflammasome activation. Immunity 2011, 34, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Iliev, I.D.; Kitazawa, H.; Shimosato, T.; Katoa, S.; Morita, H.; He, F.; Hosoda, M.; Saito, T. Strong immunostimulation in murine immune cells by Lactobacillus rhamnosus GG DNA containing novel oligodeoxynucleotide pattern. Cell. Microbiol. 2005, 7, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Girardin, S.E.; Boneca, I.G.; Viala, J.; Chamaillard, M.; Labigne, A.; Thomas, G.; Philpott, D.J.; Sansonetti, P.J. Nod2 is a general sensor of peptidoglycan through muramyl dipeptide (MDP) detection. J. Biol. Chem. 2003, 278, 8869–8872. [Google Scholar] [CrossRef] [PubMed]
- Triantafilou, M.; Miyake, K.; Golenbock, D.T.; Triantafilou, K. Mediators of innate immune recognition of bacteria concentrate in lipid rafts and facilitate lipopolysaccharide-induced cell activation. J. Cell Sci. 2002, 115, 2603–2611. [Google Scholar] [PubMed]
- Jackson, S.K.; Abate, W.; Tonks, A.J. Lysophospholipid acyltransferases: Novel potential regulators of the inflammatory response and target for drug discovery. Pharmacol. Ther. 2008, 119, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Schmid, B.; Finnen, M.J.; Harwood, J.L.; Jackson, S.K. Acylation of lysophosphatidylcholine plays a key role in the response of monocytes to lipopolysaccharide. Eur. J. Biochem. 2003, 270, 2728–2788. [Google Scholar] [CrossRef]
- Neville, N.T.; Parton, J.; Harwood, J.L.; Jackson, S.K. The activity of monocyte lysophosphatidylcholine acyltransferase and coenzyme A-independent transacylase is changed by the inflammatory cytokines tumour necrosis factor alpha and interferon gamma. Biochim. Biophys. Acta 2005, 1733, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.K.; Parton, J. Lysophospholipid acyltransferases in monocyte inflammatory responses and sepsis. Immunobiol. 2004, 209, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.K.; Abate, W.; Oarton, J.; Jones, S.; Harwood, J.L. Lysophospholipid metabolism facilitates Toll-like receptor 4 membrane translocation to regulate the inflammatory response. J. Leukoc. Biol. 2008, 84, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Neilsen, P.M.; Noll, J.E.; Mattiske, S.; Bracken, C.P.; Gregory, P.A.; Schulz, R.B.; Lim, S.P.; Kumar, R.; Suetani, R.J.; Goodall, G.J.; et al. Mutant p53 drives invasion in breast tumours through up-regulation of miR-155. Oncogene 2013, 32, 2992–3000. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Zhang, X.; Sun, W.; Hou, X.; Yao, Z.; Zhu, Y. CRIPR/CAS9-mediated genome editing of miRNA-155 inhibits proinflammatory cytokine production by RAW264.7 cells. BioMed. Res. Int. 2015, 2015, e326042. [Google Scholar]
- Bala, S.; Marcos, M.; Kodys, K.; Csak, T.; Catalano, D.; Mandrekar, P.; Szabo, G. Up-regulation of microRNA-155 in macrophages contributes to increased tumour necrosis factor a (TNFα) production via increased Mrna half-life in alcoholic liver disease. J. Biol. Chem. 2011, 286, 1436–1444. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, K.A.; Gaudet, A.D.; Amici, S.A.; Popovich, P.G.; Guerau-de-Arellano, M. Control of the inflammatory macrophage transcriptional signature by miR-155. PLoS ONE 2016, 11, e0159724. [Google Scholar] [CrossRef] [PubMed]
- Quinn, E.M.; Wang, J.H.; O’Callaghan, G.; Redmond, H.P. MicroRNA-146a is upregulated by and negatively regulates TLR2 signaling. PLoS ONE 2013, 8, e62232. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer-Redmer, S.; Bekurtz, J.C.; Arends, D.; Bortfeldt, R.; Kutz-Lohroff, B.; Sharbati, S.; Einspanier, R.; Brockmann, G.A. Feeding of Enterococcus faecium NCIMB 10415 leads to intestinal miRNA-423–5p-induced regulation of immune-relevent genes. Appl. Environ. Microbiol. 2016, 82, 2263–2269. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K.; Lopez-Collazo, E. Endotoxin tolerance: New mechanisms, molecules and clinical significance. Trends Immunol. 2009, 30, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; O’Connell, R.M.; Mazmanian, S.K. Coordination of tolerogenic immune responses by commensal microbiota. J. Autoimmun. 2010, 34, J220–J225. [Google Scholar] [CrossRef] [PubMed]
- Kitazawa, H.; Ishii, Y.; Uemura, J.; Kawai, Y.; Saito, T.; Kaneko, T.; Noda, K.; Itoh, T. Augmentation of macrophage functions by an extracellular phosphopolysaccharide from Lactobacillus delbrueckii ssp. Bulgaricus. Food Microbiol. 2000, 17, 109–118. [Google Scholar] [CrossRef]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef] [PubMed]
- Petrof, E.O.; Claud, E.C.; Sun, J.; Abramova, T.; Guo, Y.; Waypa, T.S.; He, S.-M.; Nakagawa, Y.; Chang, E.B. Bacteria-free solution derived from Lactobacillus plantarum inhibits multiple NF-kappaB pathways and inhibits proteasome function. Inflamm. Bowel Dis. 2009, 15, 1537–1547. [Google Scholar] [CrossRef] [PubMed]
- Frick, J.-S.; Schenk, K.; Quitadamo, M.; Kahl, F.; Koberle, M.; Bohn, E.; Aepfelbacher, M.; Autenrieth, I.B. Lactobacillus fermentum attenuates the proinflammatory effect of Yersinia enterocolitica on human epithelial cells. Inflamm. Bowel Dis. 2007, 13, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, B.; Schmitter, J.-M.; Urdaci, M.C. Identification of novel proteins secreted by Lactobacillus rhamnosus GG grown in de Mann-Rogosa-Sharpe broth. Lett. Appl. Microbiol. 2009, 48, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Seth, A.; Yan, F.; Polk, D.B.; Rao, R.K. Probiotics ameliorate the hydrogen-peroxide-induced epithelial barrier disruption by a PKC- and MAP kinase-dependent mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1060–G1069. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qu, X.; Yang, H.; Kang, H.; Xu, Y.; Bai, B.; Song, W. Bifidobacteria DNA induces murine macrophage activation in vitro. Cell Mol. Immunol. 2005, 2, 473–478. [Google Scholar] [PubMed]
- Lurhs, H.; Gerke, T.; Muller, J.G.; Melcher, R.; Schauber, F.; Boxberger, F.; Scheppach, W.; Menzel, T. Butyrate inhibits NF-Kb activation in lamina propria macrophages of patients with ulcerative colitis. Scand. J. Gastroenterol. 2002, 27, 458–466. [Google Scholar]
- Leung, C.H.; Lam, W.; Ma, D.L.; Gullen, E.A.; Cheng, Y.C. Butyrate mediates nucleotide-binding and oligomerisation domain (NOD) 2-dependent mucosal immune responses against peptidoglycan. Eur. J. Immunol. 2009, 39, 3529–3537. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S.; Kalliomäki, M.; Isolauri, E. Probiotics during pregnancy and breast-feeding might confer immunomodulatory protection against atopic disease in the infant. J. Allergy Clin. Immunol. 2002, 109, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, G.; Rao, S.; Patole, S.; Bulsara, M. Updated meta-analysis of probiotics for preventing necrotizing enterocolitis in preterm neonates. Pediatrics 2010, 125, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Tursi, A.; Brandimarte, G.; Papa, A.; Giglio, A.; Elisei, W.; Giorgetti, G.M.; Forti, G.; Morini, S.; Hassan, C.; Pistoia, M.A.; et al. Treatment of relapsing mild-to-moderate ulcerative colitis with the probiotic VSL#3 as adjunctive to a standard pharmaceutical treatment: A double-blind, randomized, placebo-controlled study. Am. J. Gastroenterol. 2010, 105, 2218–2227. [Google Scholar] [PubMed]




{kind=link}
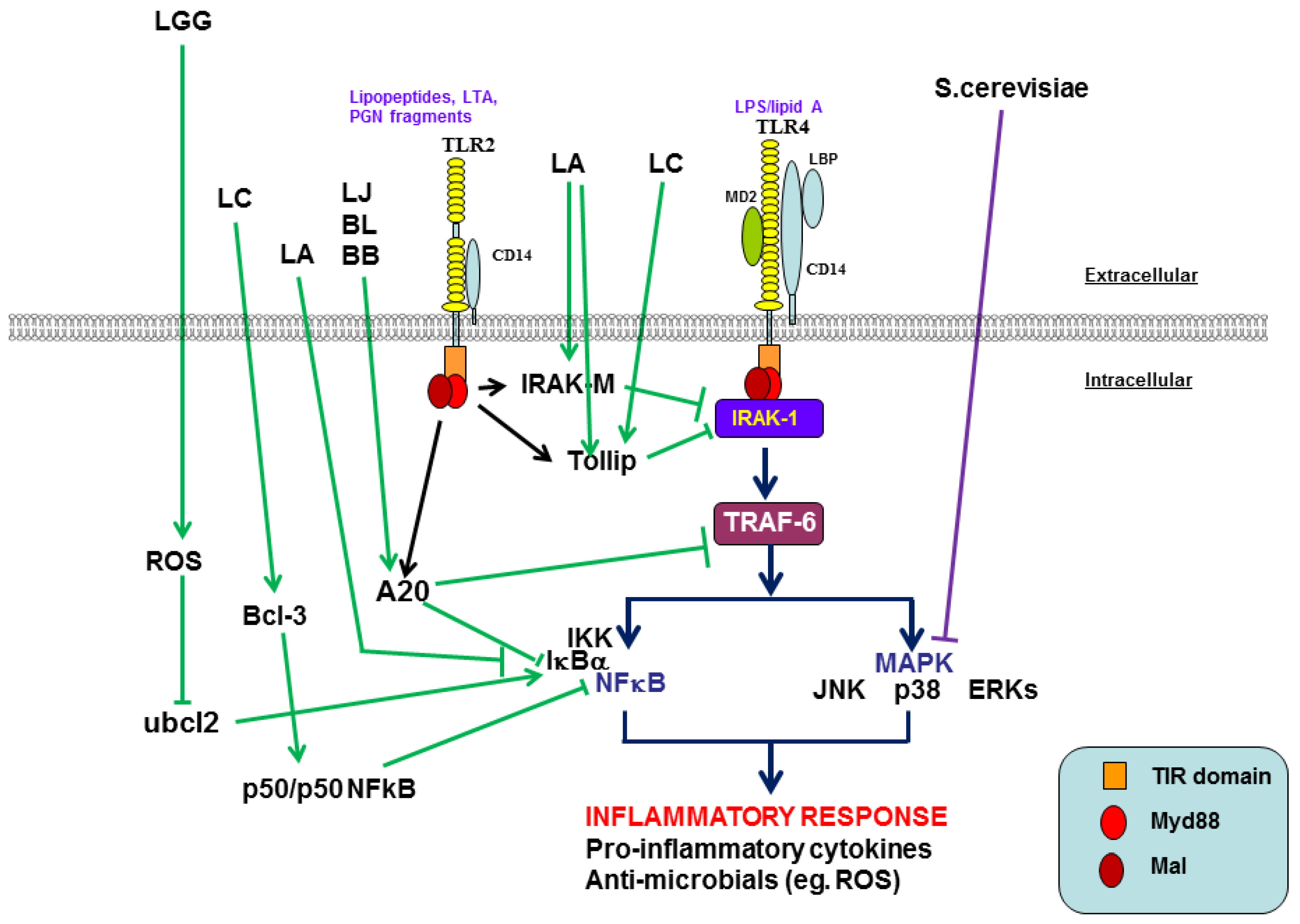{kind=link}
{kind=link}
| Probiotic Species (Soluble Product) | Probiotic–Induced Effector Response | Cells/Cell Line | Signaling Pathway | Reference |
|---|---|---|---|---|
| Bifidobacteria breve M-16V Bifidobacteria longum BB536 | Increase IL-8, MCP-1 and IL-6 levels | PIE cells | NFκB via activation of A20 | [25] |
| Bifidobacteria lactis KCTC 5727 | Suppress NFκB-binding activity & IκB degradation | HT-29 cells | NFκB | [26] |
| Lactobacillus amylovorus DSM 16698T | Decrease ETEC-induced IL-8 & IL-1β production | Caco-2/TC7 cells (higher transport activity). | TLR4 signaling via activation of Tollip and IRAK-M | [27] |
| Lactobacillus casei OLL2768 | Decrease IL-6, IL-8, IL-1α and MCP-1 | BIE cells | NFκB and p38 MAPK | [28] |
| Lactobacillus rhamnosus GG | Increased expression of IL-10R2 | Ex vivo C57 BL/6J mice Immature colon samples | Phospho-STAT3, increased expression of SOCS-3 | [29] |
| Lactobacillus rhamnosus GG | TLR2 up-regulation | IPEC-J2 | TLR signaling | [18] |
| Lactobacillus rhamnosus GG (LGG) | Reduce NFκB activation via ROS | FHs74Int cells (human foetal) | NFκB | [30] |
| Lactobacillus rhamnosus GG p40 and p75 proteins | Enhance tight junction | YAMC | EGFR/Akt | [23,24,31] |
| Lactobacillus rhamnosus GG p40 protein (LGGp40) | Decrease cytokine-induced apoptosis | YAMC | EGFR/Akt via Src | [22] |
| Lactobacillus jensenii TL2937 | Activate negative regulators A20, Bcl-3 and MKP-1 | PIE cells | TLR4-dependent NFκB and MAPK | [32] |
| Lactobacillus plantarum (LP) WCFS1 | Increased tight junction proteins | Caco-2 cells | TLR2 signaling | [17] |
| Lactobacillus reuteri DSM 17938 | Inhibit IκB phosphorylation | Ex vivo rat model | NFκB | [33] |
| Saccharomyces cerevisiae (Sc) CNCM I-3856 | Decrease IL-6 and IL-8 expression | IPEC-1 | Decrease of ERK1/2 and p38 phosphorylation | [34] |
| Streptococcus salivarius K12 | Inhibit NFκB activity | HT-29 | NFκB | [35] |
| VSL#3 | Increased tight junction proteins | HT-29 cells | Phosphorylation of ERK and p38 MAPKs | [20] |
| VSL#3 | Induction of heat shock proteins (hsp) | Colonic IECs | Suppression of NFκB via inhibition of proteosome | [36] |
| Probiotic | T Cell Activation | PRR and Signaling Pathway | Reference |
|---|---|---|---|
| Bifidobacterium breve Yakult strain | Tr1 cells | TLR2/MyD88 | [60] |
| B. longum BB536 DNA | Increased CD4+CD25+ Treg cells & Th1 activation | TLR9 and IκB-α phosphorylation | [55] |
| IRT5 | CD4+Foxp3+ regulatory T cells | Not described. | [62] |
| LGG DNA | Increased CD4+CD25+ Treg cells and Th1 activation | TLR9 and IκB-α phosphorylation | [55] |
| LGG DNA | ND | TLR9 and reduced IκBα degradation and p38 phosphorylation | [56] |
| Lactobacillus paracasei ATCC 25302 DNA | CD4+ Foxp3+ Treg cells | TLR9 | [57] |
| Lactobacillus salivarius Ls33 | CD4+ Foxp3+ Treg cells | NOD2 | [61] |
| Probiotic | Effector Response | Cells/Cell Line/Model | Reference |
|---|---|---|---|
| L. gasseri NC1500 L. gasseri + MnSOD NC1501 | Reduction in severity of inflammation —decreased Nφ & Mφ infiltration | IL-10 deficient mouse model of colitis | [75] |
| B. animalis MB5L. rhamnosus GG Probiotics & Secreted factors | Inhibition of Nφ transmigration by suppression of IL-8, Gro-α, ENAP-78 and suppression of chemokine regulators, IL-1β & TNFα. | Enterotoxigenic E.coli K88 – induced inflammation in Caco-2 epithelial cells | [76] |
| B. lactis HN019 | Increased Nφ phagocytic capacity & enhanced phagocyte-mediated bacteriocidal activity. | Clinical trial: Healthy human PBMCs | [77] |
| E. coli Nissle 1917 | Amelioration of inflammation: Reduced levels of Nφs & chemokines. | DSS-induced colitis, murine model. Prophylaxis – faecal transplantation model. | [78] |
| L. rhamnosus NutRes1 B. breve NutRes204 | Worsening of faecal condition accompanied by reduced Nφ numbers. L. rhamnosus improved this & increased TLR2/6, CCL2, TNFα, IL-1β, IL-6. | DSS-induced chronic colitis, murine model. Relapse–Remision cycling. | [79] |
| B. longum subsp. Infantis BB-02 | Reduction in inflammation: decreased Nφ infiltration & CXCL-1. | DSS-induced acute colitis, murine model. | [80] |
| L. rhamnosus L34 L. casei L39 | Suppression of IL-8 expression. Conditioned media: L34 suppressed NFκB phosphorylation, L39 suppressed both NFκB & c-Jun. | C. difficile-associated disease colitis model. Infant faecal isolates on HT-29 epithelial cells. | [81] |
| L. gasseri ATC33323 cell wall extract | Cell wall extract—lethality. Increased TNFα, IL-1β, MIP-1α, MCP-1, NO & decreased Nφ count. | Sepsis model in Sprague-Dawley rats. | [82] |
| L. rhamnosus Lcr35 | AM development prevented by suppressing IL-4 (Th2), IL-17 (Th17), TSLP via a Foxp3+ Treg-dependent mechanism. | Murine OVA challenge allergic march (AM) model. | [83] |
| L. rhamnosus GG | Inhibition of PMA- and S.aureus-induced neutrophil extracellular traps (NETs), ROS production & phagocytic capacity. | HL60-derived Nφ cell model. | [84] |
| B. longum OLL6001 Culture condensate (BCC) | Augmentation of Nφ recruitment. Upregulation of adhesion molecule & cytokine expression. Immuno-enhancing. | Diet-restricted murine peritonitis model: ip glycogen injection. | [85] |
| VSL#3 Probiotic mixture | Reduction in mucosal levels of Nφ chemoattractant, IL-8 & tissue influx of Nφs. | Human retrospective study of proB-treated UC pouchitis. | [86] |
| L. plantarum ATCC10241 | Reduction in bacterial load, Nφ numbers, apoptotic/necrotic cells and IL-8. Wound healing. | Chronic infected leg ulcers (diabetic and non-diabetic). | [87] |
| Clostridium butyricum MIYAIRI 588 | Anti-tumour effects by MMP-8 driven release of TRAIL (TLR2/4–dependent) | Nφs in BCG-responsive bladder cancer patients | [88] |
| Probiotic | Effector Response | Cells/Cell Line | Reference |
|---|---|---|---|
| L. casei Shirota L. fermentum MS15 L. plantarumNCIMB41605 L. salivarius NCIMB41606 B. breve NCIMB 8807 | TNFα: Increased—CD14hi M1/M2 Decreased—CD14lo M1/M2 IL-6: Decreased—M2 NF-κB: Decreased—M1 Increased—M2 | M1 & M2 Mφ subsets (THP-1 cell line) | [103] [104] |
| Secreted Protein | +/− IL-6 M1 Mφs +/− IL-8/TNFα | M1 & M2 Mφ subsets (THP-1) | [103] [104] |
| SCFA-butyrate | Suppression of IL-12 & augmentation of IL-10 | Human monocytes | [105] |
| SCFA-butyrate | Decreased TNFα Increased IL-10 | M1 & M2 Mφ subsets (THP-1) | [106] |
| L. helveticus DSM13137 L. rhamnosus GG S. thermophilus THS B. breve DSM13692 | Increased IL-10:IL-12 ratios Induced SOCS3 (IL-10 & p38 MAPK-dependent) | Human Mφs GM-CSF-PB monocytes | [91] |
| L. plantarum K8 KCTC10887BP LTA (pLTA) | Suppression of LPS Induced TNFα (pLTA suppn of ERK, JNK, p38 MAPK phosphorylation, IκB degradation & TLR4. Induction of IRAK-M. | THP-1 & U937 pro-monocytes Murine sepsis model L929-BMM Balb/c | [96] |
| L. plantarum KCTC10887BP LTA (pLTA) | Suppression of S. aureus LTA (aLTA) induced TNFα pLTA suppresses Myd88, NFκB & MAPKs. Antagonises aLTA. | THP-1 pro-monocytes | [97] |
| L. plantarum KCTC10887BP LTA (pLTA) | Suppression of S. flexneri PGN Induced inflammation (TNFα, IL-1β). Toln decreases NOD2; pLTA cross Toln Flex PGN | THP-1 pro-monocytes | [98] |
| B. breve ATCC15700 B. longum ATCC15697 Enterococcus faecalis ATCC19433 | Differential modulation of TNFα, IL-1β, IL-12p40 mRNA. Decreased IκB phosphorylation and induction of SOCS-1, -3. | RAW264.7 murine Mφs LPS-stimulated | [100] |
| LAB LTA L. plantarum ATCC14917T L.casei YIT9029 | Regulate TLR2-dependent ERK mediated IL-12/IL-10 axis: LP high IL-10/IL-12, LC low IL-10/IL-12. | Murine peritoneal Mφs Balb/c | [107] |
| L. rhamnosus GR1 & GG | G-CSF-mediated inhibition of JNK: Suppression of TNFα | Murine immort peritoneal & BMM C57Bl/6 Human THP-1s | [89] |
| ProB DNA L. casei & B.breve | Induction of IL-1, IL-10 & IL-6 | Human PBMCs | [108] |
| L. paracasei Cultech | TLR2-dependent up-regulation of negative regulators of NFκB | Human PBMCs & PMA-THP-1 cells. | [109] |
| L. reuteri ATCC PTA 6475 CF48-3A & ATCC55730 conditioned medium | Suppression of LPS-induced TNFα & MCP-1 via inhibition of MAPK-driven cJun/AP-1actn | THP-1 cells, MonoMac-6 cells, CD Mφs | [99] |
| Probiotic Format | Effector Response | Cells/Cell Line | Reference |
|---|---|---|---|
| L. fermentum MS15 L. plantarum NCIMB41605 L.Salivarius NCIMB41606 B.Breve NCIMB8807 Secreted Protein | TNFα: Increased—CD14hi M1 Selective suppression—CD14loM1/M2 IL-6: Decreased—M2 NFκB: Selective suppression—CD14hiM1. Increased—M2 | M1 & M2 Mφ subsets (THP-1 cell line) | [103] |
| L. casei Shirota Secreted Protein | Suppressed LPS-induced TNFα, NFκB activation & augmented IL-1β (M1 Mφs). Suppressed LPS-induced IL-6 & augmented IL-1β (M2 Mφs), independent of NFκB activation. | M1 & M2 Mφ subsets (THP-1 cell line) | [104] |
| SCFA-butyrate | Suppression of LPS/PGN-induced TNFα (M1 & M2). Suppression of LPS/PGN-induced IL-1β (M2). Augmentation of LPS/PGN-induced IL-10 (M2). Suppression of monocyte LPS/PGN—induced TNFα & IL-1β. | M1 & M2 Mφ subsets (THP-1 cell line) | [106] |
| L. plantarum (Non-designated strain) Conditioned medium (Lp-CM) | Inhibition of NFκB binding activity andproteosome-dependent degredation of IκBα. (Stimuli: TNFα, LPS, Flagellin, Poly I:C). Suppression of MCP-1 secretion. | Murine YAMC intestinal epithelial cells, RAW264.7 Mφs, Primary DCs | [135] |
| L. plantarum K8 KCTC10887BP LTA (pLTA) | Suppression of LPS-induced TNFα pLTA suppn of ERK, JNK, p38 MAPK phosphorylation, IκB degradation & TLR4. Induction of IRAK-M expression. | THP-1 & U937 pro-monocytes. Murine sepsis model L929-BMM Balb/c | [96] |
| L. plantarum KCTC10887BP LTA (pLTA) | Suppression of S. aureus LTA (aLTA) induced TNFα pLTA suppresses Myd88, NFκB & MAPKs. Antagonises aLTA. | THP-1 pro-monocytes | [97] |
| L. plantarum KCTC10887BP LTA (pLTA) | Suppression of S.flexneri PGN induced inflammation (TNFα, IL-1β). Toln decreases NOD2; pLTA cross Toln Flex PGN | THP-1 pro-monocytes | [98] |
| L. rhamnosus GG p75 & p40 secreted proteins | Protection from H2O2-mediated damage to IEC barrier TJs (PKC & MAPK-dependent). | IECs | [134] [138] |
| LAB LTA L.plantarum ATCC14917T L.casei YIT9029 | Regulate TLR2-dependent ERK mediated IL-12/IL-10 axis: LP high IL-10/IL-12, LC low IL-10/IL-12. | Murine peritoneal Mφs Balb/c | [107] |
| L. rhamnosus GG secreted protein Serpin B1, protease inhibitor of neutrophil elastase. Band G2 & G4. | Prevention of epithelial cell barrier damage. | Mass Spec. (MS/MS) analysis of LGG S/N grown in MRS broth. | [137] |
| ProB DNA L.casei & B.breve | Induction of IL-1, IL-10 & IL-6 | Human PBMCs | [108] |
| L. rhamnosus GG DNA | Immunostimulation | Murine immune cells B cells | [117] |
| Bifidobacteria (Non-designated strain) unmethylated CpG DNA | Augmentation of Mφ phagocytosis, NO release and secretion of IL-1β, IL-6, IL-12p40 and TNFα (CpG DNA recognised by TLR9). | Murine Mφs : J774A.1 cells | [139] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Llewellyn, A.; Foey, A. Probiotic Modulation of Innate Cell Pathogen Sensing and Signaling Events. Nutrients 2017, 9, 1156. https://doi.org/10.3390/nu9101156
Llewellyn A, Foey A. Probiotic Modulation of Innate Cell Pathogen Sensing and Signaling Events. Nutrients. 2017; 9(10):1156. https://doi.org/10.3390/nu9101156
Chicago/Turabian StyleLlewellyn, Amy, and Andrew Foey. 2017. "Probiotic Modulation of Innate Cell Pathogen Sensing and Signaling Events" Nutrients 9, no. 10: 1156. https://doi.org/10.3390/nu9101156
APA StyleLlewellyn, A., & Foey, A. (2017). Probiotic Modulation of Innate Cell Pathogen Sensing and Signaling Events. Nutrients, 9(10), 1156. https://doi.org/10.3390/nu9101156