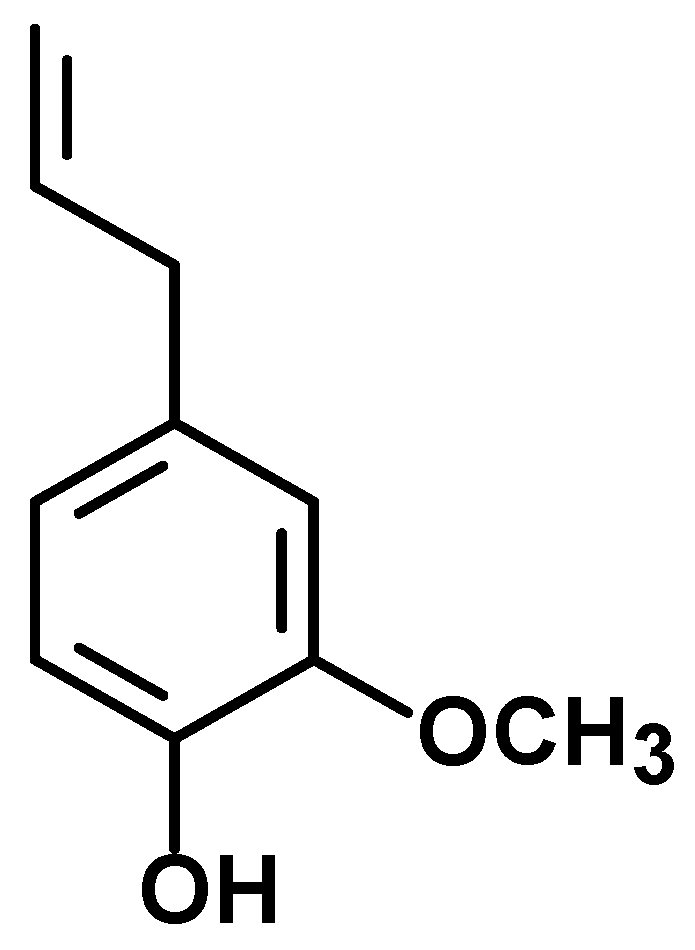
The Dual Antioxidant/Prooxidant Effect of Eugenol and Its Action in Cancer Development and Treatment

 ,
,  and
and 
Abstract
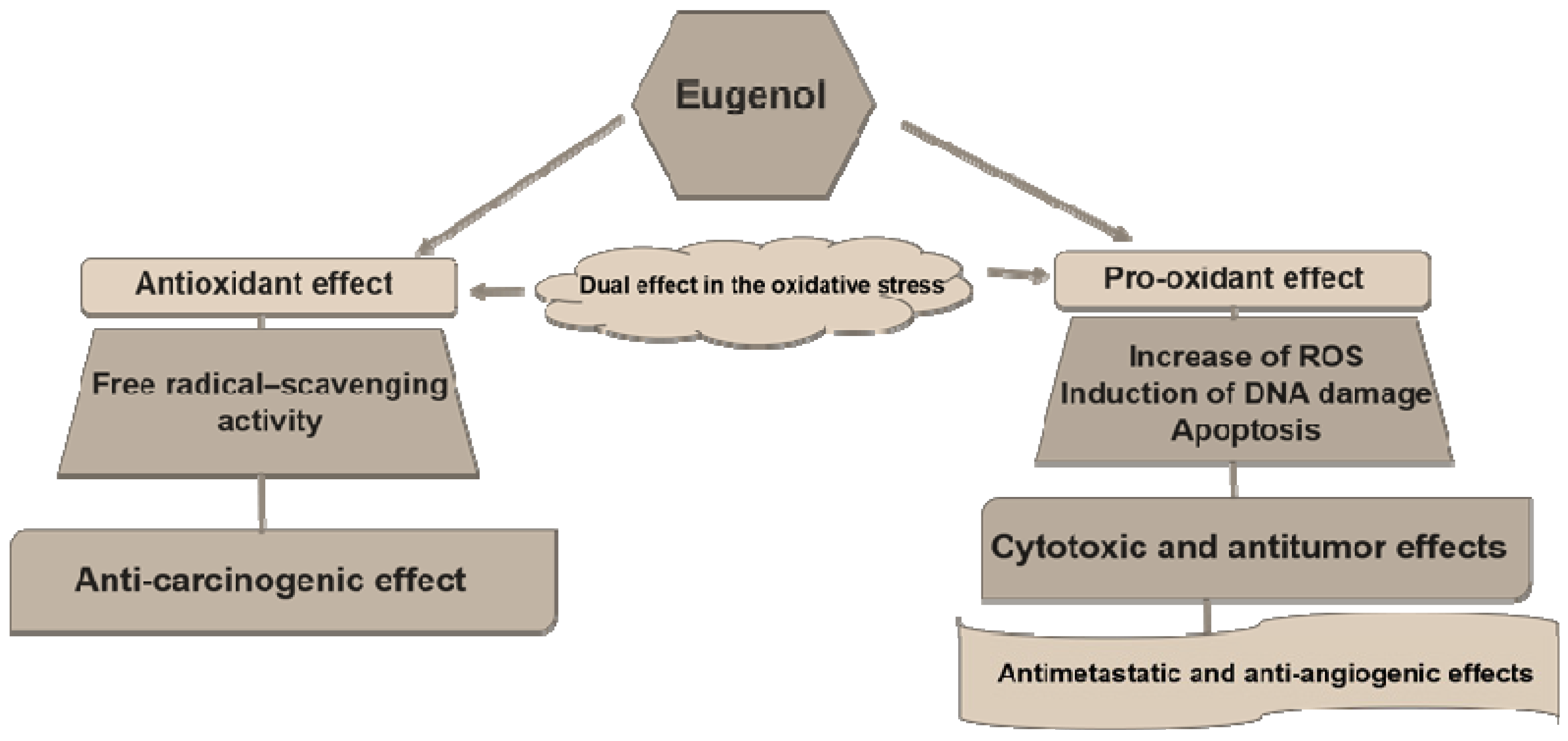
:1. Introduction
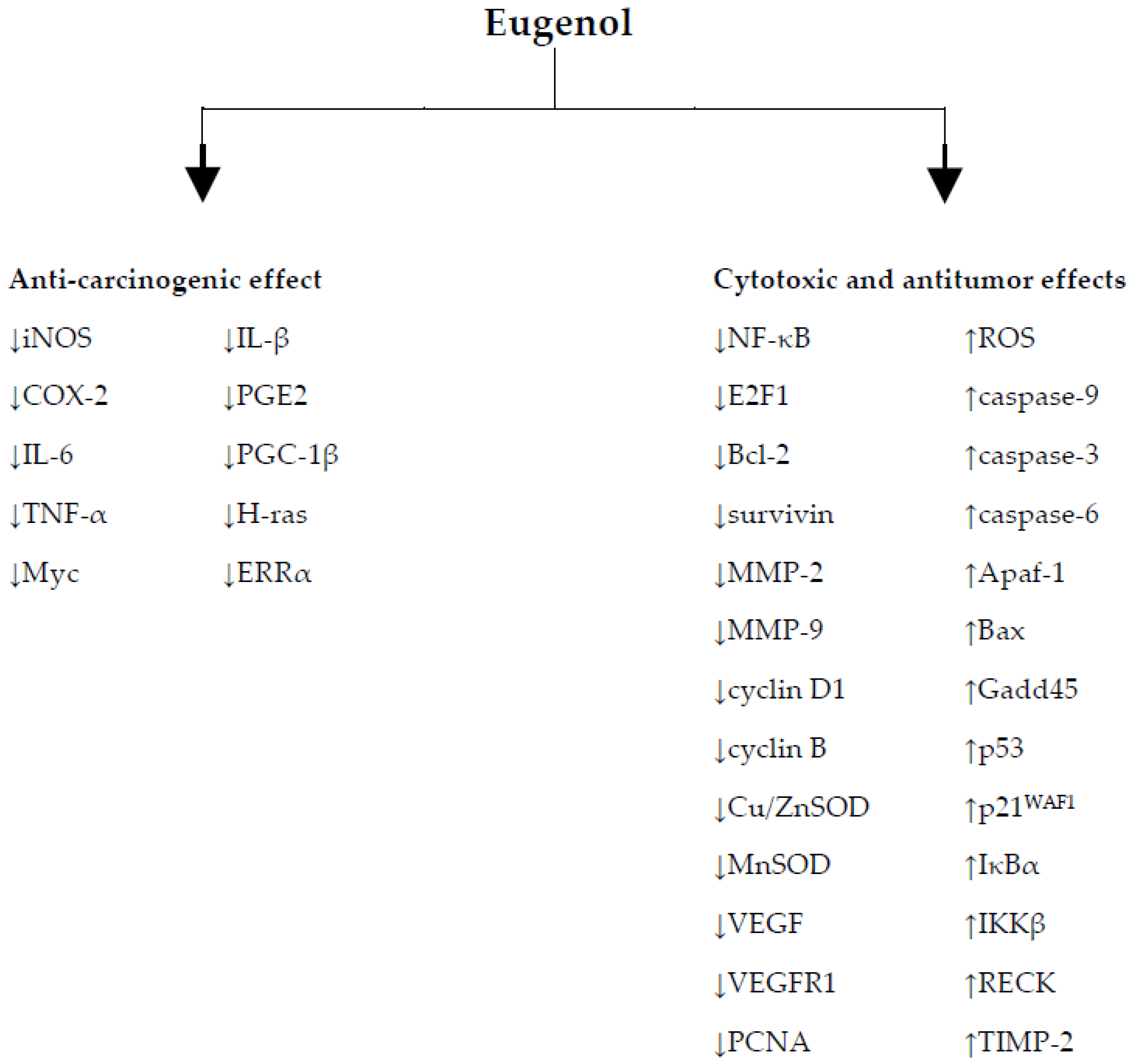
2. Anti-Carcinogenic/Chemopreventive Effect of Eugenol and Its Relation to the Inhibition of Oxidative Stress
3. Cytotoxic and Antitumor Effects of Eugenol and Its Relation to the Induction of Oxidative Stress
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Apaf-1 | Apoptotic protease activating factor 1 |
| Bax | BCL2 associated X, apoptosis regulator |
| Bcl-2 | B-cell lymphoma 2, apopstosis regulator |
| COX-2 | Cyclooxygenase-2 |
| CYP 1A1 | Cytochrome P450 family 1 subfamily A member 1 |
| CYP1B1 | Cytochrome P450 family 1 subfamily B member 1 |
| DFF45 | DNA fragmentation factor 45 |
| DMBA | 7,12-dimethylbenz[a]anthracene |
| DNA | Deoxyribonucleic acid |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| ER | Endoplasmic reticulum |
| Gadd45 | Growth arrest and DNA damage-inducible 45 |
| IC50 | Half maximal inhibitory concentration |
| IKKβ | Inhibitor of kappa B |
| IL-6 | Interleukin-6 |
| iNOS | Inducible nitric oxide synthase |
| IκBα | IκB Kinase α |
| MA | Malonaldehyde |
| MCF 10A-ras | H-ras transfected MCF 10A |
| MMP | Matrix metalloproteinases |
| MNNG | N-Methyl-N′-nitro-N-nitrosoguanidine |
| NF-κB | Nuclear factor kappa B |
| PARP | Poly(ADP-ribose)polymerase |
| PCNA | Proliferating cell nuclear antigen |
| PGE2 | Prostaglandin E2 |
| RECK | Reversion-inducing-cysteine-rich protein with kazal motifs |
| ROS | Reactive Oxygen Species |
| SGOT | Serum glutamic-oxaloacetic transaminase |
| TA | Thioacetamide |
| TBARS | Thiobarbituric acid-reactive substances |
| TIMP-2 | Tissue inhibitor of metalloproteinase-2 |
| TNF-α | Tumor necrosis factor alpha |
| TPA | 12-otetradecanoylphorbol-13-acetate |
| VEGF | Vascular endothelial growth factor |
| VEGFR1 | Vascular endothelial growth factor receptor 1 |
References
- Selim, K.A.; Abdelrasoul, H.; Aboelmagd, M.; Tawila, A.M. The role of the MAPK signaling, topoisomerase and dietary bioactives in controlling cancer incidence. Diseases 2017, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Gào, X.; Schöttker, B. Reduction-oxidation pathways involved in cancer development: A systematic review of literature reviews. Oncotarget 2017, 8, 51888–51906. [Google Scholar] [CrossRef] [PubMed]
- Fiaschi, T.; Chiarugi, P. Oxidative stress, tumor microenvironment, and metabolic reprogramming: A diabolic liaison. Int. J. Cell Biol. 2012, 762825. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, D.P.; Soares, A.K.; de Sousa, D.P. Overview of the role of vanillin on redox status and cancer development. Oxid. Med. Cell Longev. 2016, 9734816. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.A.; Andrade, L.N.; De Sousa, E.B.; De Sousa, D.P. Antitumor phenylpropanoids found in essential oils. Biomed. Res. Int. 2015, 392674. [Google Scholar] [CrossRef] [PubMed]
- Sobral, M.V.; Xavier, A.L.; Lima, T.C.; De Sousa, D.P. Antitumor activity of monoterpenes found in essential oils. Sci. World J. 2014, 953451. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, D.P. Bioactive Essential Oils and Cancer; Springer: New York, NY, USA, 2015. [Google Scholar]
- Ferraz, R.P.; Bomfim, D.S.; Carvalho, N.C.; Soares, M.B.; da Silva, T.B.; Machado, W.J.; Prata, A.P.; Costa, E.V.; Moraes, V.R.; Nogueira, P.C.; et al. Cytotoxic effect of leaf essential oil of Lippia gracilis Schauer (Verbenaceae). Phytomedicine 2013, 20, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.Y.; Zubair, H.; Ullah, M.F.; Ahmad, A.; Hadi, S.M. A prooxidant mechanism for the anticancer and chemopreventive properties of plant polyphenols. Curr. Drug Targets 2012, 13, 1738–1749. [Google Scholar] [CrossRef] [PubMed]
- Forester, S.C.; Lambert, J.D. The role of antioxidant versus pro-oxidant effects of green tea polyphenols in cancer prevention. Mol. Nutr. Food Res. 2011, 55, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Assi, M. The differential role of reactive oxygen species in early and late stages of cancer. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R646–R653. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-G.; Shibamoto, T. Antioxidant properties of aroma compounds isolated from soybeans and mung beans. J. Agric. Food Chem. 2000, 48, 4290–4293. [Google Scholar] [CrossRef] [PubMed]
- Charalambous, G. The Quality of Foods and Beverages V2: Chemistry and Technology, 1st ed.; Academic Press: New York, NY, USA, 1981; p. 408. [Google Scholar]
- Jordán, M.J.; Tandon, K.; Shaw, P.E.; Goodner, K.L. Aromatic profile of aqueous banana essence and banana fruit by gas chromatography-mass spectrometry (GC-MS) and gas chromatography-olfactometry (GC-O). J. Agric. Food Chem. 2001, 49, 4813–4817. [Google Scholar] [CrossRef] [PubMed]
- Kamatou, G.P.; Vermaak, I.; Viljoen, A.M. Eugenol-from the remote maluku islands to the international market place: A review of a remarkable and versatile molecule. Molecules 2012, 17, 6953–6981. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y.M.; Patterson, A.; Piggot, J.R.; Richardson, G.D. Origins of flavour in whiskies and a revised flavour wheel: A review. J. Inst. Brew. 2001, 107, 287–313. [Google Scholar] [CrossRef]
- Bohnert, H.J.; Nguyen, H.R.; Lewis, N.G. Bioengineering and Molecular Biology of Plant Pathways; Elsevier: San Diego, CA, USA, 2008. [Google Scholar]
- Srinivasan, K. Antioxidant potential of spices and their active constituents. Crit. Rev. Food Sci. Nutr. 2014, 54, 352–372. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Athar, M.; Alam, M.S. Eugenol precludes cutaneous chemical carcinogenesis in mouse by preventing oxidative stress and inflammation and by inducing apoptosis. Mol. Carcinog. 2010, 49, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, K.; Unnikrishnan, M.C.; Kuttan, R. Inhibition of tumour promotion in mice by eugenol. Indian J. Physiol. Pharmacol. 1994, 38, 306–308. [Google Scholar] [PubMed]
- Pal, D.; Banerjee, S.; Mukherjee, S.; Roy, A.; Panda, C.K.; Das, S. Eugenol restricts DMBA croton oil induced skin carcinogenesis in mice: Downregulation of c-Myc and H-ras, and activation of p53 dependent apoptotic pathway. J. Dermatol. Sci. 2010, 59, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Van Duuren, B.L.; Goldschmidt, B.M. Cocarcinogenic and tumor-promoting agents in tobacco carcinogenesis. J. Natl. Cancer Inst. 1976, 56, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Azuine, M.A.; Amonkar, A.J.; Bhide, S.V. Chemopreventive efficacy of betel leaf extract and its constituents on 7,12-dimethylbenz(a)anthracene induced carcinogenesis and their effect on drug detoxification system in mouse skin. Indian J. Exp. Biol. 1991, 29, 346–351. [Google Scholar] [PubMed]
- Manikandan, P.; Murugan, R.S.; Priyadarsini, R.V.; Vinothini, G.; Nagini, S. Eugenol induces apoptosis and inhibits invasion and angiogenesis in a rat model of gastric carcinogenesis induced by MNNG. Life Sci. 2010, 86, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, P.; Vinothini, G.; Vidya Priyadarsini, R.; Prathiba, D.; Nagini, S. Eugenol inhibits cell proliferation via NF-κB suppression in a rat model of gastric carcinogenesis induced by MNNG. Invest. New Drugs 2011, 29, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhang, G.; Bie, F.; Lv, Y.; Ma, Y.; Ma, M.; Wang, Y.; Hao, X.; Yuan, N.; Jiang, X. Eugenol inhibits oxidative phosphorylation and fatty acid oxidation via downregulation of c-Myc/PGC-1β/ERRα signaling pathway in MCF10A-ras cells. Sci. Rep. 2017, 7, 12920. [Google Scholar] [CrossRef] [PubMed]
- Koujitani, T.; Yasuhara, K.; Tamura, T.; Onodera, H.; Takagi, H.; Takizawa, T.; Hirose, M.; Hayashi, Y.; Mitsumori, K. Lack of modifying effects of eugenol on development of lung proliferative lesions induced by urethane in transgenic mice carrying the human prototype c-Ha-ras gene. J. Toxicol. Sci. 2001, 26, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Murakami, K.; Yoshino, M. Antioxidant action of eugenol compounds: Role of metal ion in the inhibition of lipid peroxidation. Food Chem. Toxicol. 2005, 43, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Nagababu, E.; Rifkind, J.M.; Boindala, S.; Nakka, L. Assessment of antioxidant activity of eugenol in vitro and in vivo. Methods Mol. Biol. 2010, 610, 165–180. [Google Scholar] [PubMed]
- Gülçin, İ. Antioxidant activity of eugenol: A structure-activity relationship study. J. Med. Food 2011, 14, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.; Kim, M.M. Eugenol with antioxidant activity inhibits MMP-9 related to metastasis in human fibrosarcoma cells. Food Chem. Toxicol. 2013, 55, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Horvathova, E.; Navarova, J.; Galova, E.; Sevcovicova, A.; Chodakova, L.; Snahnicanova, Z.; Melusova, M.; Kozics, K.; Slamenova, D. Assessment of antioxidative, chelating, and DNA-protective effects of selected essential oil components (eugenol, carvacrol, thymol, borneol, eucalyptol) of plants and intact Rosmarinus officinalis oil. J. Agric. Food Chem. 2014, 62, 6632–6639. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, S.K.; Roy, S. Phytopharmacological approach of free radical scavenging and anti-oxidative potential of eugenol and Ocimum gratissimum Linn. Asian Pac. J. Trop. Med. 2014, 7S1, S391–S397. [Google Scholar] [CrossRef]
- Zhang, L.L.; Zhang, L.F.; Xu, J.G.; Hu, Q.P. Comparison study on antioxidant, DNA damage protective and antibacterial activities of eugenol and isoeugenol against several foodborne pathogens. Food Nutr. Res. 2017, 61, 1353356. [Google Scholar] [CrossRef] [PubMed]
- Yogalakshmi, B.; Viswanathan, P.; Anuradha, C.V. Investigation of antioxidant, anti-inflammatory and DNA-protective properties of eugenol in thioacetamide-induced liver injury in rats. Toxicology 2010, 268, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, M.; Hisama, M. Suppression of chemical mutagen-induced SOS response by alkylphenols from clove (Syzygium aromaticum) in the Salmonella typhimurium TA1535/pSK1002 umu test. J. Agric. Food Chem. 2001, 49, 4019–4025. [Google Scholar] [CrossRef] [PubMed]
- Rompelberg, C.J.; Verhagen, H.; van Bladeren, P.J. Effects of the naturally occurring alkenylbenzenes eugenol and trans-anethole on drug-metabolizing enzymes in the rat liver. Food Chem. Toxicol. 1993, 31, 637–645. [Google Scholar] [CrossRef]
- Han, E.H.; Hwang, Y.P.; Jeong, T.C.; Lee, S.S.; Shin, J.G.; Jeong, H.G. Eugenol inhibit 7,12-dimethylbenz[a]anthracene-induced genotoxicity in MCF-7 cells: Bifunctional effects on CYP1 and NAD(P)H: Quinone oxidoreductase. FEBS Lett. 2007, 581, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.K. Anti-genotoxicity of trans-anethole and eugenol in mice. Food Chem. Toxicol. 2001, 39, 493–498. [Google Scholar] [CrossRef]
- Rompelberg, C.J.; Evertz, S.J.; Bruijntjes-Rozier, G.C.; van den Heuvel, P.D.; Verhagen, H. Effect of eugenol on the genotoxicity of established mutagens in the liver. Food Chem. Toxicol. 1996, 34, 33–42. [Google Scholar] [CrossRef]
- Schiestl, R.H.; Chan, W.S.; Gietz, R.D.; Mehta, R.D.; Hastings, P.J. Safrole, eugenol and methyleugenol induce intrachromosomal recombination in yeast. Mutat. Res. 1989, 224, 427–436. [Google Scholar] [CrossRef]
- Stich, H.F.; Stich, W.; Lam, P.P.S. Potentiation of genotoxicity by concurrent application of compounds found in betel quid: Arecoline, eugenol, quercetin, chlorogenic acid and Mn2+. Mutat. Res. 1981, 90, 355–363. [Google Scholar] [CrossRef]
- National Toxicology Program (NTP). Carcinogenesis Studies of Eugenol (CAS No. 97–53–0) in F344/N Rats and B6C3F1 Mice (Feed Studies). Natl. Toxicol. Program Tech. Rep. Ser. 1983, 223, 1–159. [Google Scholar]
- Ghosh, R.; Nadiminty, N.; Fitzpatrick, J.E.; Alworth, W.L.; Slaga, T.J.; Kumar, A.P. Eugenol causes melanoma growth suppression through inhibition of E2F1 transcriptional activity. J. Biol. Chem. 2005, 280, 5812–5819. [Google Scholar] [CrossRef] [PubMed]
- Yoo, C.B.; Han, K.T.; Cho, K.S.; Ha, J.; Park, H.J.; Nam, J.H.; Kil, U.H.; Lee, K.T. Eugenol isolated from the essential oil of Eugenia caryophyllata induces a reactive oxygen species-mediated apoptosis in HL-60 human promyelocytic leukemia cells. Cancer Lett. 2005, 225, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Bodell, W.J.; Ye, Q.; Pathak, D.N.; Pongracz, K. Oxidation of eugenol to form DNA adducts and 8-hydroxy-2’-deoxyguanosine: Role of quinone methide derivative in DNA adduct formation. Carcinogenesis 1998, 19, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, A.; Espinoza, C.; Cardile, V.; Gallardo, C.; Cardona, W.; Lombardo, L.; Catalán, M.; Cuellar, F.; Russo, A. Eugenol and its synthetic analogues inhibit cell growth of human cancer cells. J. Braz. Chem. Soc. 2008, 19, 543–548. [Google Scholar] [CrossRef]
- Ghosh, R.; Ganapathy, M.; Alworth, W.L.; Chan, D.C.; Kumar, A.P. Combination of 2-methoxyestradiol (2-ME2) and eugenol for apoptosis induction synergistically in androgen independent prostate cancer cells. J. Steroid Biochem. Mol. Biol. 2009, 113, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Al-Sharif, I.; Remmal, A.; Aboussekhra, A. Eugenol triggers apoptosis in breast cancer cells through E2F1/survivin down-regulation. BMC Cancer 2013, 13, 600. [Google Scholar] [CrossRef] [PubMed]
- Júnior, P.L.; Câmara, D.A.; Costa, A.S.; Ruiz, J.L.; Levy, D.; Azevedo, R.A.; Pasqualoto, K.F.; de Oliveira, C.F.; de Melo, T.C.; Pessoa, N.D.; et al. Apoptotic effect of eugenol envolves G2/M phase abrogation accompanied by mitochondrial damage and clastogenic effect on cancer cell in vitro. Phytomedicine 2016, 23, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Young, S.C.; Wang, C.J.; Hsu, J.D.; Hsu, J.L.; Chou, F.P. Increased sensitivity of Hep G2 cells toward the cytotoxicity of cisplatin by the treatment of piper betel leaf extract. Arch Toxicol. 2006, 80, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, A.; Sakanashi, Y.; Matsui, H.; Oyama, T.; Nishimura, Y.; Masuda, T.; Oyama, Y. Cytometric analysis of cytotoxicity of polyphenols and related phenolics to rat thymocytes: Potent cytotoxicity of resveratrol to normal cells. Basic Clin. Pharmacol. Toxicol. 2009, 104, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Slamenová, D.; Horváthová, E.; Wsólová, L.; Sramková, M.; Navarová, J. Investigation of anti-oxidative, cytotoxic, DNA-damaging and DNA-protective effects of plant volatiles eugenol and borneol in human-derived HepG2, Caco-2 and VH10 cell lines. Mutat. Res. 2009, 677, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, S.K.; Supriyanto, E. Antiproliferative and molecular mechanism of eugenol-induced apoptosis in cancer cells. Molecules 2012, 17, 6290–6304. [Google Scholar] [CrossRef] [PubMed]
- Koh, T.; Machino, M.; Murakami, Y.; Umemura, N.; Sakagami, H. Cytotoxicity of dental compounds towards human oral squamous cell carcinoma and normal oral cells. In Vivo 2013, 27, 85–95. [Google Scholar] [PubMed]
- Sharma, U.K.; Sharma, A.K.; Gupta, A.; Kumar, R.; Pandey, A.; Pandey, A.K. Pharmacological activities of cinnamaldehyde and eugenol: Antioxidant, cytotoxic and anti-leishmanial studies. Mol. Cell. Biol. 2017, 63, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, S.; Atsumi, T.; Satoh, K.; Kadoma, Y.; Ishihara, M.; Okada, N.; Nagasaki, M.; Yokoe, I.; Sakagami, H. Radical generation, radical-scavenging activity, and cytotoxicity of eugenol-related compounds. In Vitro Mol. Toxicol. 2000, 13, 269–280. [Google Scholar]
- Fujisawa, S.; Atsumi, T.; Ishihara, M.; Kadoma, Y. Cytotoxicity, ROS-generation activity and radical-scavenging activity of curcumin and related compounds. Anticancer Res. 2004, 24, 563–569. [Google Scholar] [PubMed]
- Atsumi, T.; Fujisawa, S.; Tonosaki, K. A comparative study of the antioxidant/prooxidant activities of eugenol and isoeugenol with various concentrations and oxidation conditions. Toxicol. In Vitro 2005, 19, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Okada, N.; Hirata, A.; Murakami, Y.; Shoji, M.; Sakagami, H.; Fujisawa, S. Induction of cytotoxicity and apoptosis and inhibition of cyclooxygenase-2 gene expression by eugenol-related compounds. Anticancer Res. 2005, 25, 3263–3269. [Google Scholar] [PubMed]
- Atsumi, T.; Iwakura, I.; Fujisawa, S.; Ueha, T. Reactive oxygen species generation and photo-cytotoxicity of eugenol in solutions of various pH. Biomaterials 2001, 22, 1459–1466. [Google Scholar] [CrossRef]
- Fujisawa, S.; Atsumi, T.; Satoh, K.; Sakagami, H. Interaction between 2-ethoxybenzoic acid (EBA) and eugenol, and related changes in cytotoxicity. J. Dent. Res. 2003, 82, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Pisano, M.; Pagnan, G.; Loi, M.; Mura, M.E.; Tilocca, M.G.; Palmieri, G.; Fabbri, D.; Dettori, M.A.; Delogu, G.; Ponzoni, M.; et al. Antiproliferative and pro-apoptotic ctivity of eugenol-related biphenyls on malignant melanoma cells. Mol. Cancer 2007, 6, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.C.; Choi, D.S.; Lim, J.S.; Jeong, H.C.; Kim, I.R.; Lee, M.H.; Park, B.S. Caspases-dependent apoptosis in human melanoma cell by eugenol. Korean J. Anat. 2006, 39, 245–253. [Google Scholar]
- Shin, S.H.; Park, J.H.; Kim, G.C. The mechanism of apoptosis induced by eugenol in human osteosarcoma cells. J. Korean Assoc. Oral Maxillofac. Surg. 2007, 33, 20–27. [Google Scholar]
- Hussain, A.; Brahmbhatt, K.; Priyani, A.; Ahmed, M.; Rizvi, T.A.; Sharma, C. Eugenol enhances the chemotherapeutic potential of gemcitabine and induces anticarcinogenic and anti-inflammatory activity in human cervical cancer cells. Cancer Biother. Radiopharm. 2011, 26, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Priyani, A.; Sadrieh, L.; Brahmbhatt, K.; Ahmed, M.; Sharma, C. Concurrent sulforaphane and eugenol induces differential effects on human cervical cancer cells. Integr. Cancer Ther. 2012, 11, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, S.K.; Mazumdar, A.; Mondhe, D.; Mandal, M. Apoptotic effect of eugenol in human colon cancer cell lines. Cell Biol. Int. 2011, 35, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Vidhya, N.; Devaraj, S.N. Induction of apoptosis by eugenol in human breast cancer cells. Indian J. Exp. Biol. 2011, 49, 871–878. [Google Scholar] [PubMed]
- Al Wafai, R.; El-Rabih, W.; Katerji, M.; Safi, R.; El Sabban, M.; El-Rifai, O.; Usta, J. Chemosensitivity of MCF-7 cells to eugenol: Release of cytochrome-c and lactate dehydrogenase. Sci. Rep. 2017, 7, 43730. [Google Scholar] [CrossRef] [PubMed]
- Koh, T.; Murakami, Y.; Tanaka, S.; Machino, M.; Onuma, H.; Kaneko, M.; Sugimoto, M.; Soga, T.; Tomita, M.; Sakagami, H. Changes of metabolic profiles in an oral squamous cell carcinoma cell line induced by eugenol. In Vivo 2013, 27, 233–243. [Google Scholar] [PubMed]
- Majeed, H.; Antoniou, J.; Fang, Z. Apoptotic effects of eugenol-loaded nanoemulsions in human colon and liver cancer cell lines. Asian Pac. J. Cancer Prev. 2014, 15, 9159–9164. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, S.K.; Mondhe, D.; Wani, Z.A.; Pal, H.C.; Mandal, M. Effect of honey and eugenol on ehrlich ascites and solid carcinoma. J. Biomed. Biotechnol 2010, 989163:1–989163:5. [Google Scholar] [CrossRef] [PubMed]
- Osseni, R.A.; Rat, P.; Bogdan, A.; Warnet, J.M.; Touitou, Y. Evidence of prooxidant and antioxidant action of melatonin on human liver cell line HepG2. Life Sci. 2000, 68, 387–399. [Google Scholar] [CrossRef]
- Schwartz, J.L. The dual roles of nutrients as antioxidants and prooxidants: Their effects on tumor cell growth. J. Nutr. 1996, 126, 1221S–1227S. [Google Scholar] [PubMed]
- Shi, M.; Xu, B.; Azakami, K.; Morikawa, T.; Watanabe, K.; Morimoto, K.; Komatsu, M.; Aoyama, K.; Takeuchi, T. Dual role of vitamin C in an oxygen-sensitive system: Discrepancy between DNA damage and cell death. Free Radic Res. 2005, 39, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Chakraborthy, A.; Ramani, P.; Sherlin, H.J.; Premkumar, P.; Natesan, A. Antioxidant and pro-oxidant activity of Vitamin C in oral environment. Indian J. Dent. Res. 2014, 25, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, S.; Atsumi, T.; Kadoma, Y.; Sakagami, H. Antioxidant and prooxidant action of eugenol-related compounds and their cytotoxicity. Toxicology 2002, 177, 39–54. [Google Scholar] [CrossRef]
- Kim, D.O.; Lee, C.Y. Comprehensive study on vitamin C equivalent antioxidant capacity (VCEAC) of various polyphenolics in scavenging a free radical and its structural relationship. Crit. Rev. Food Sci. Nutr. 2004, 44, 253–273. [Google Scholar] [CrossRef] [PubMed]
- Mahboub, R.; Memmou, F. Antioxidant activity and kinetics studies of eugenol and 6-bromoeugenol. Nat. Prod. Res. 2015, 29, 966–971. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Carcinogenesis Model | Carcinogen | Eugenol Administration | Effect | References |
|---|---|---|---|---|
| Skin carcinogenesis | DMBA + TPA | Topical | Reduction in tumor incidence and size; and/or development of papillomatous keratoacanthoma with minimal cell proliferation but without squamous cell carcinoma | [20] |
| Skin carcinogenesis | DMBA + croton oil | Topical | Inhibition of tumor formation ~60% | [21,22] |
| Skin carcinogenesis | benzo[a]pyrene | Topical | Inhibition of tumor formation ~50% | [23] |
| Skin Carcinogenesis | DMBA | Topical | Minimal protection | [24] |
| Gastric carcinogenesis | MNNG | Intragastric | Inhibition of tumor formation ~75% | [25,26] |
| Lung carcinogenesis | Urethane | Oral | No protection | [28] |
| Cell Lines | Histological Type | Origin | IC50 (µM) | References |
|---|---|---|---|---|
| Cancer cells | ||||
| Sbcl2 | Primary melanoma | Human | ~0.5 | [45] |
| WM3211 | Primary melanoma | Human | ~0.5 | [45] |
| WM98-1 | Primary melanoma | Human | ~0.5 | [45] |
| WM1205Lu | Metastatic melanoma | Human | ~0.5 | [45] |
| SK-Mel-28 | Melanoma | Human | 7.2 | [51] |
| A2058 | Melanoma | Human | 12.2 | [51] |
| WM266-4 | Melanoma | Human | >100 | [64] |
| SK-Mel-28 | Melanoma | Human | >100 | [64] |
| LCP-Mel | Melanoma | Human | >100 | [64] |
| LCM-Mel | Melanoma | Human | >100 | [64] |
| PNP-Mel | Melanoma | Human | >100 | [64] |
| CN-MelA | Melanoma | Human | >100 | [64] |
| 13443 | Melanoma | Human | >100 | [64] |
| GR-Mel | Melanoma | Human | >100 | [64] |
| HSG | Submandibular gland adenocarcinoma | Human | ~100 | [59] |
| 396 | [60] | |||
| T47-D | Breast carcinoma | Human | 0.9 | [50] |
| MDA-MB-231 | Breast adenocarcinoma | Human | 1.7 | [50] |
| 15.1 | [51] | |||
| ~1600 | [71] | |||
| MCF-7 | Breast adenocarcinoma | Human | 1.5 | [50] |
| 22.8 | [51] | |||
| ~400 | [70] | |||
| 900 | [71] | |||
| HCT-15 | Colon adenocarcinoma | Human | 300 | [69] |
| HT-29 | Colon adenocarcinoma | Human | 500 | [69] |
| Caco-2 | Colon carcinoma | Human | ~750 | [73] |
| SNU-C5 | Colon carcinoma | Human | 129.4 | [46] |
| LNCaP | Prostate adenocarcinoma | Human | ~550 | [49] |
| PC-3 | Prostate carcinoma | Human | ~180 | [49] |
| DU-145 | Prostate carcinoma | Human | 30.4 | [48] |
| SIHA | Cervical carcinoma | Human | 18.3 | [51] |
| HeLa | Cervical carcinoma | Human | 500 | [72] |
| HepG2 | Hepatocellular carcinoma | Human | 118.6 | [46] |
| ~500 | [73] | |||
| 3LL | Lewis lung carcinoma | Mouse | 89.6 | [46] |
| KB | Oral squamous cell carcinoma | Human | 28.5 | [48] |
| HSC-2 | Oral squamous cell carcinoma | Human | ~700 | [72] |
| HOS | Osteosarcoma | Human | 1500 | [66] |
| HL-60 | Promyelocytic leukemia | Human | 23.7 | [46] |
| 380 | [61] | |||
| U-937 | Histocytic lymphoma | Human | 39.4 | [46] |
| Non-cancer cells | ||||
| MCF 10A | Breast epithelial | Human | 2.2 | [50] |
| Tumor | Histological Type | Origin | Dose (mg/kg) | Treatment | Route | Inhibition Rate (%) | References |
|---|---|---|---|---|---|---|---|
| B16 | Melanoma | Mouse | 125 | Twice a week | i.p. | 62 | [45] |
| Ehrlich (ascites model) | Carcinoma | Mouse | 100 | Every two days for four weeks | i.p. | 28.9 | [74] |
| Ehrlich (solid model) | Carcinoma | Mouse | 100 | Every two days for four weeks | i.p. | 24.4 | [74] |
| MDA-MB-231 | Breast adenocarcinoma | Human | 100 | Every two days for four weeks | i.p. | ~66 | [50] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezerra, D.P.; Militão, G.C.G.; De Morais, M.C.; De Sousa, D.P. The Dual Antioxidant/Prooxidant Effect of Eugenol and Its Action in Cancer Development and Treatment. Nutrients 2017, 9, 1367. https://doi.org/10.3390/nu9121367
Bezerra DP, Militão GCG, De Morais MC, De Sousa DP. The Dual Antioxidant/Prooxidant Effect of Eugenol and Its Action in Cancer Development and Treatment. Nutrients. 2017; 9(12):1367. https://doi.org/10.3390/nu9121367
Chicago/Turabian StyleBezerra, Daniel Pereira, Gardenia Carmen Gadelha Militão, Mayara Castro De Morais, and Damião Pergentino De Sousa. 2017. "The Dual Antioxidant/Prooxidant Effect of Eugenol and Its Action in Cancer Development and Treatment" Nutrients 9, no. 12: 1367. https://doi.org/10.3390/nu9121367
APA StyleBezerra, D. P., Militão, G. C. G., De Morais, M. C., & De Sousa, D. P. (2017). The Dual Antioxidant/Prooxidant Effect of Eugenol and Its Action in Cancer Development and Treatment. Nutrients, 9(12), 1367. https://doi.org/10.3390/nu9121367