Palmitoleic Acid Inhibits RANKL-Induced Osteoclastogenesis and Bone Resorption by Suppressing NF-κB and MAPK Signalling Pathways



Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Fatty Acid Preparation
2.3. Cell Culture
2.4. Alamar Blue Assay for Cell Viability
2.5. Osteoclast Formation and TRAP Activity Assay
2.6. Actin Ring Formation
2.7. Bone Resorption Assay
2.8. Polymerase Chain Reaction (PCR)
2.9. Western Blotting
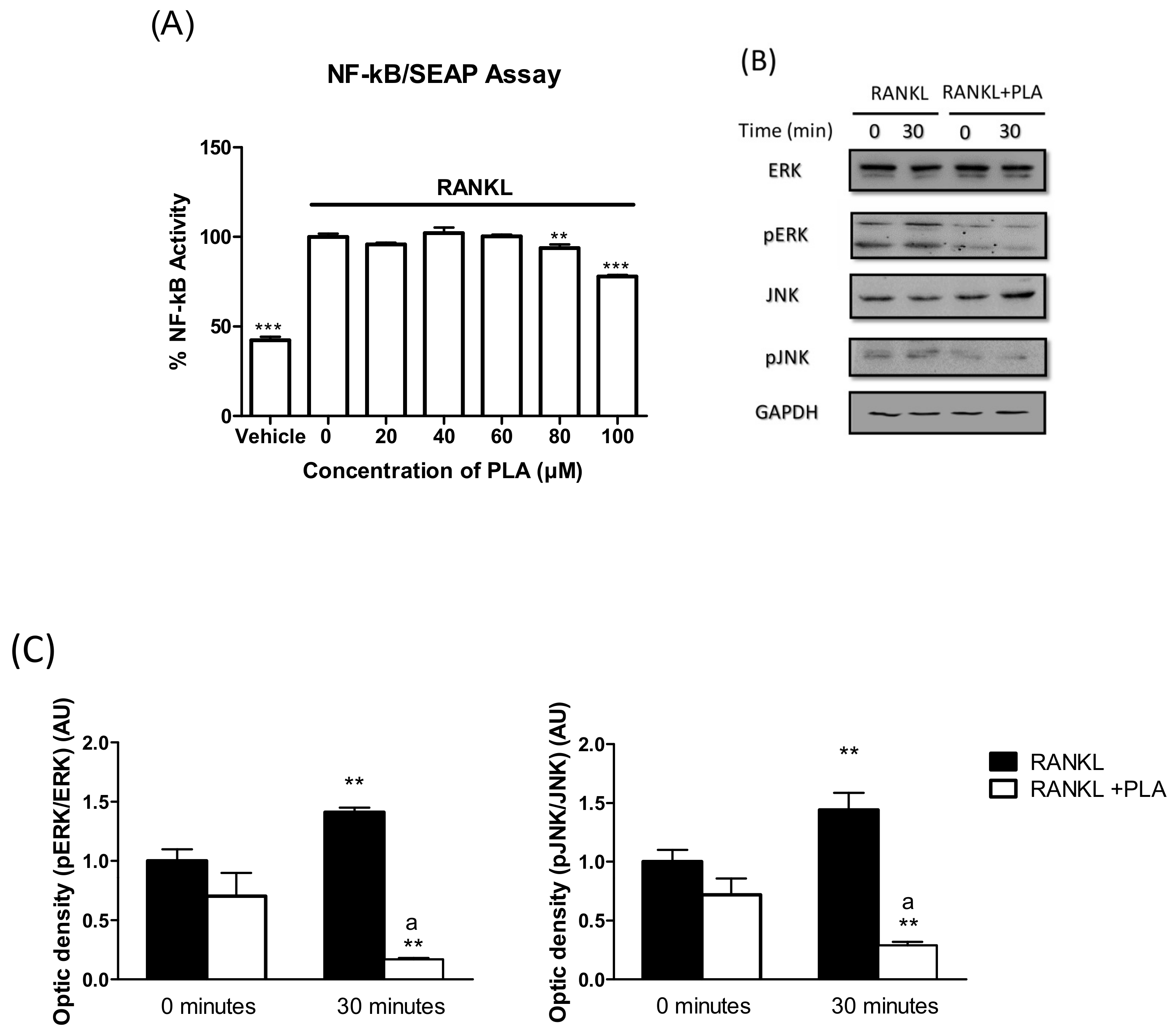
2.10. NF-κB/SEAP Assay
2.11. Mature Osteoclasts
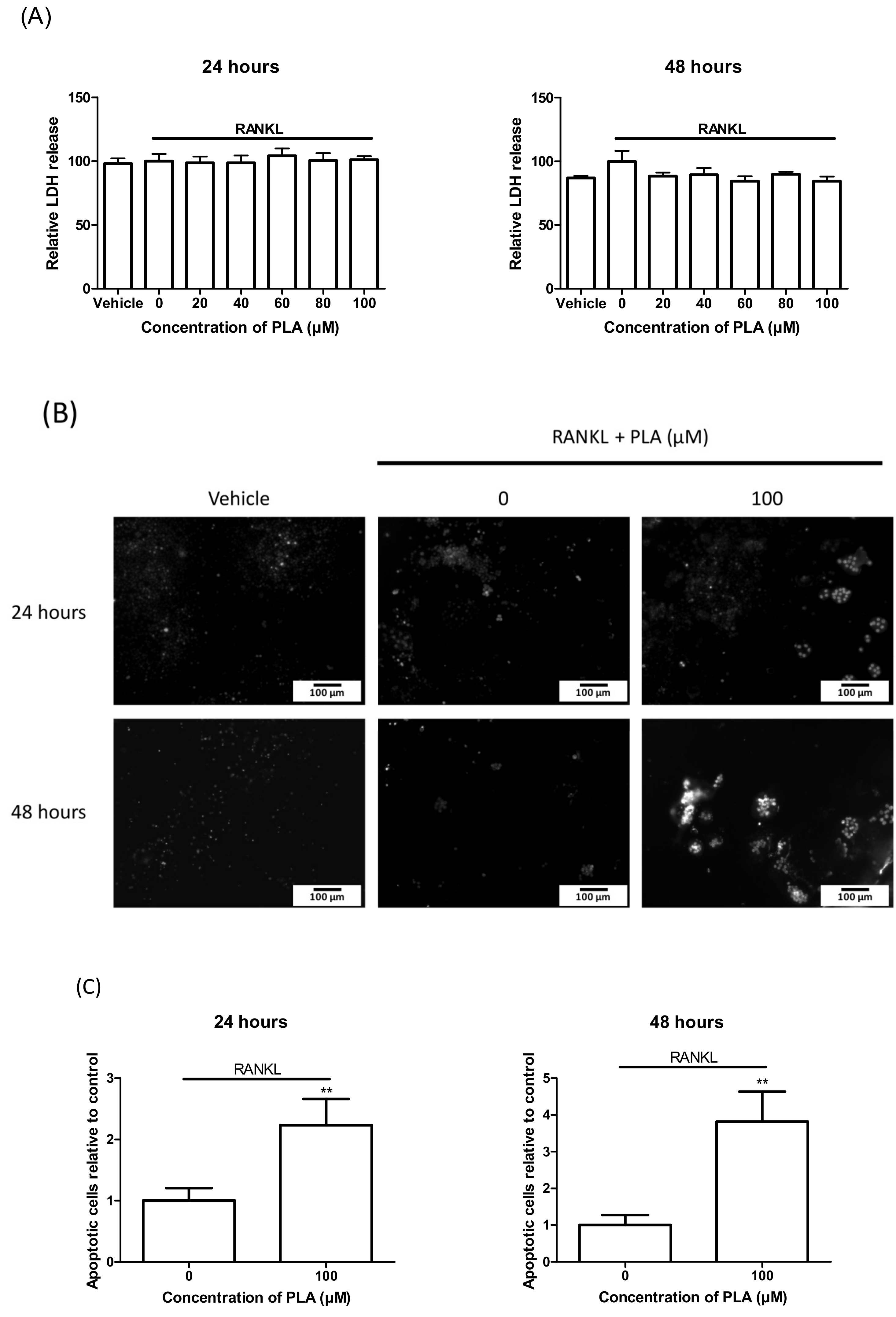
2.11.1. LDH Assay
2.11.2. Hoechst Staining
2.11.3. Statistical Analysis
3. Results
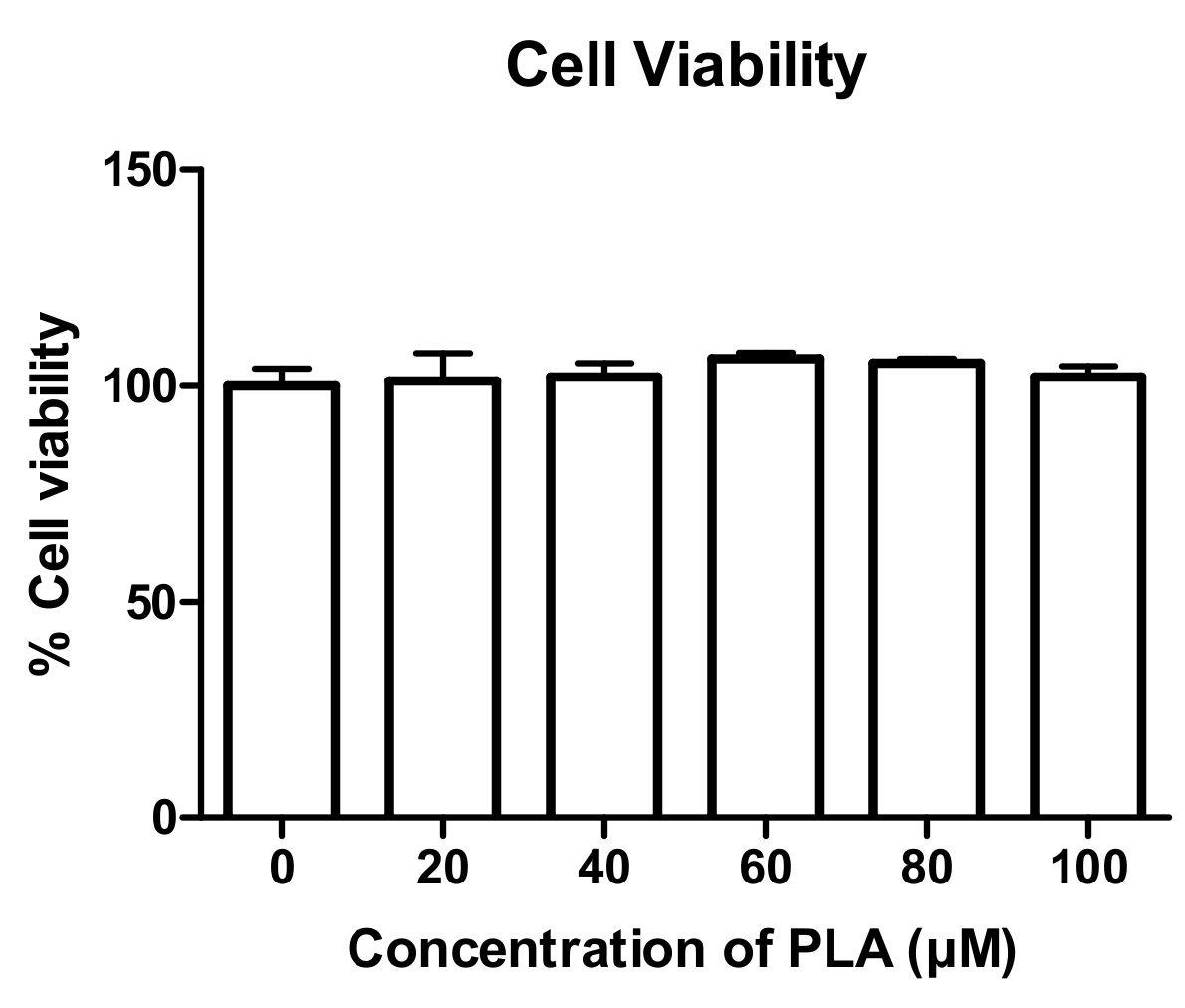
3.1. The Effect of PLA on Cell Viability
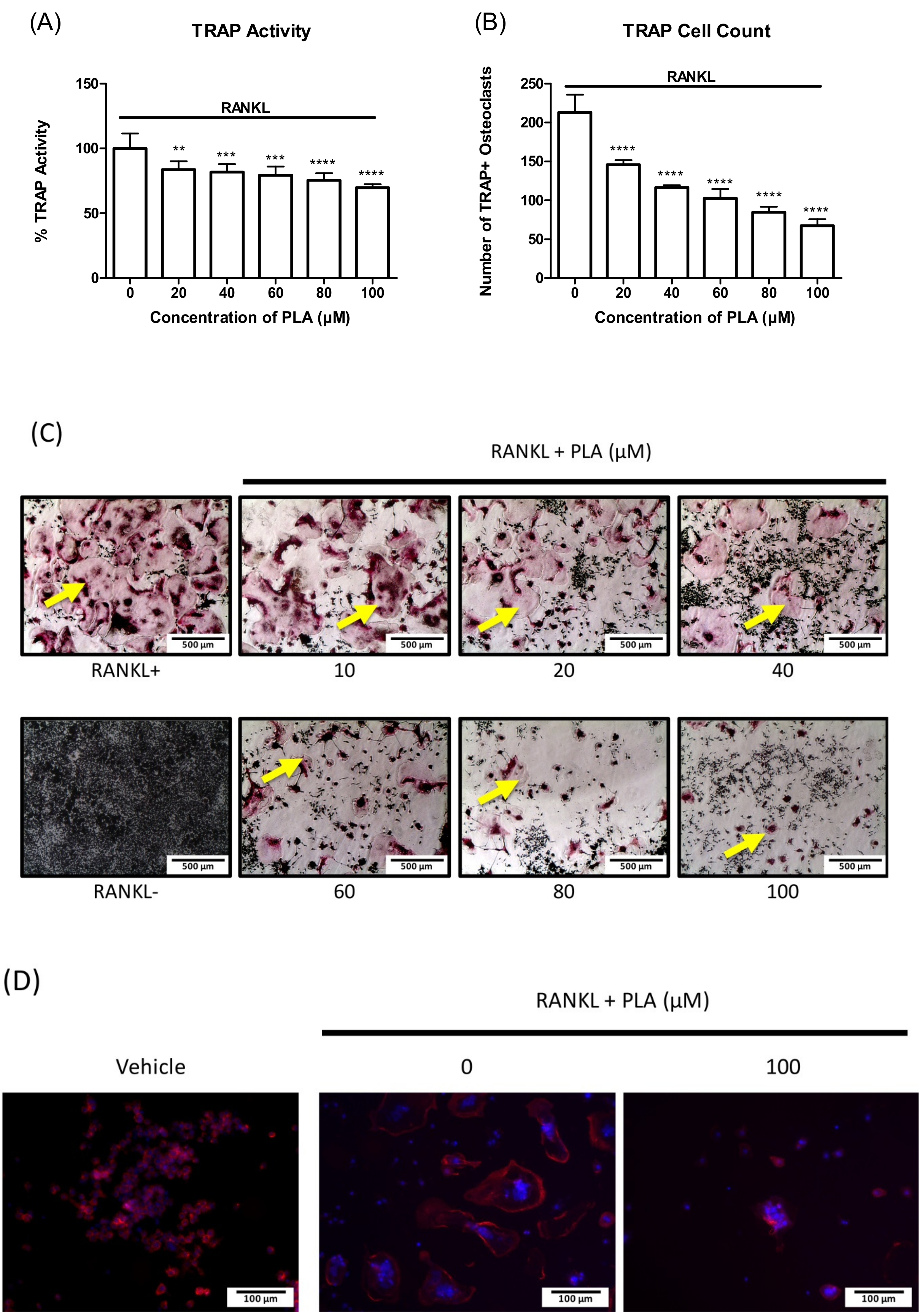
3.2. PLA Inhibits Osteoclast Formation
3.3. The Effect of PLA on RAW264.7 Murine Macrophage Morphology
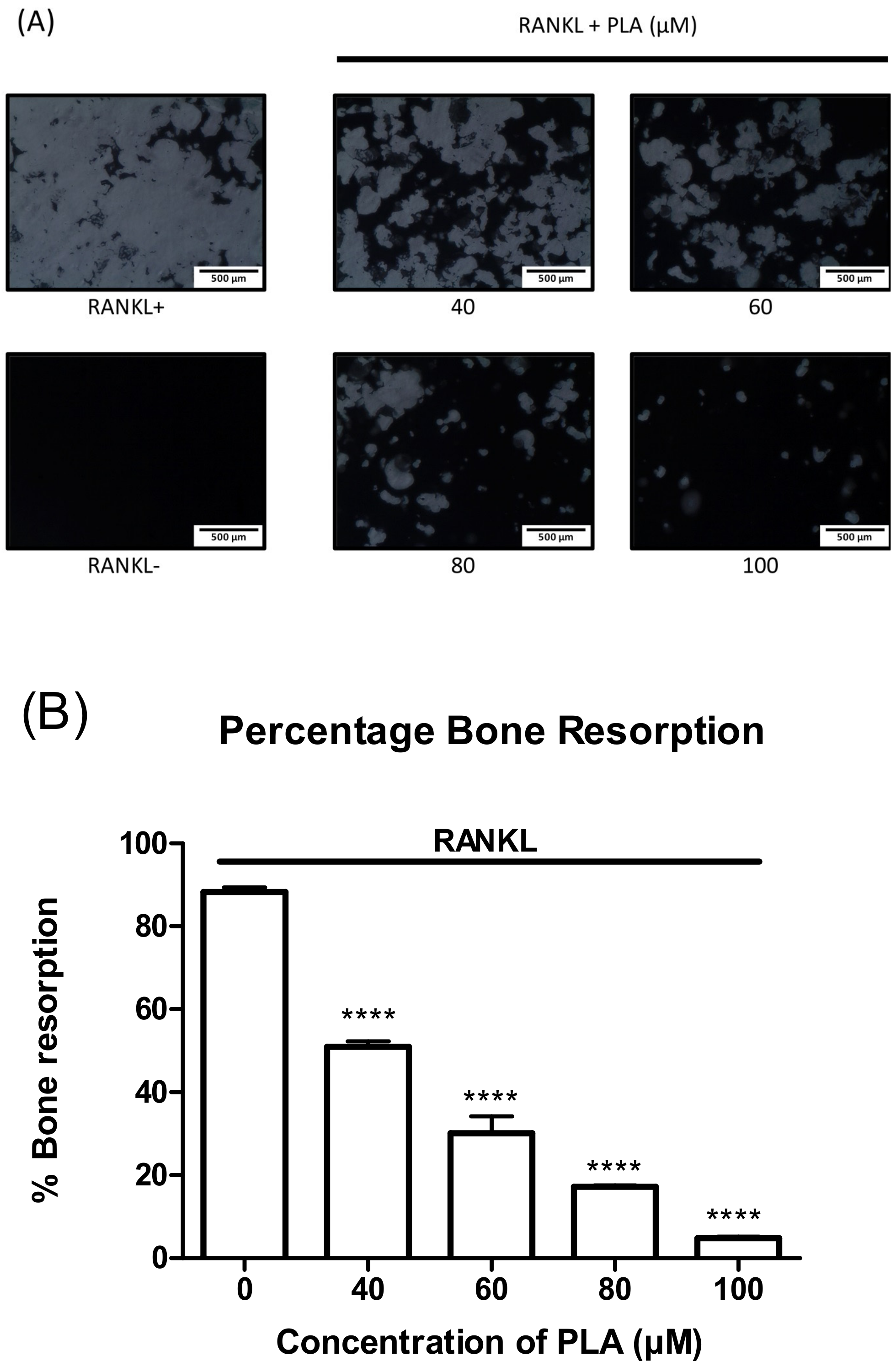
3.4. PLA Suppresses Bone Resorption
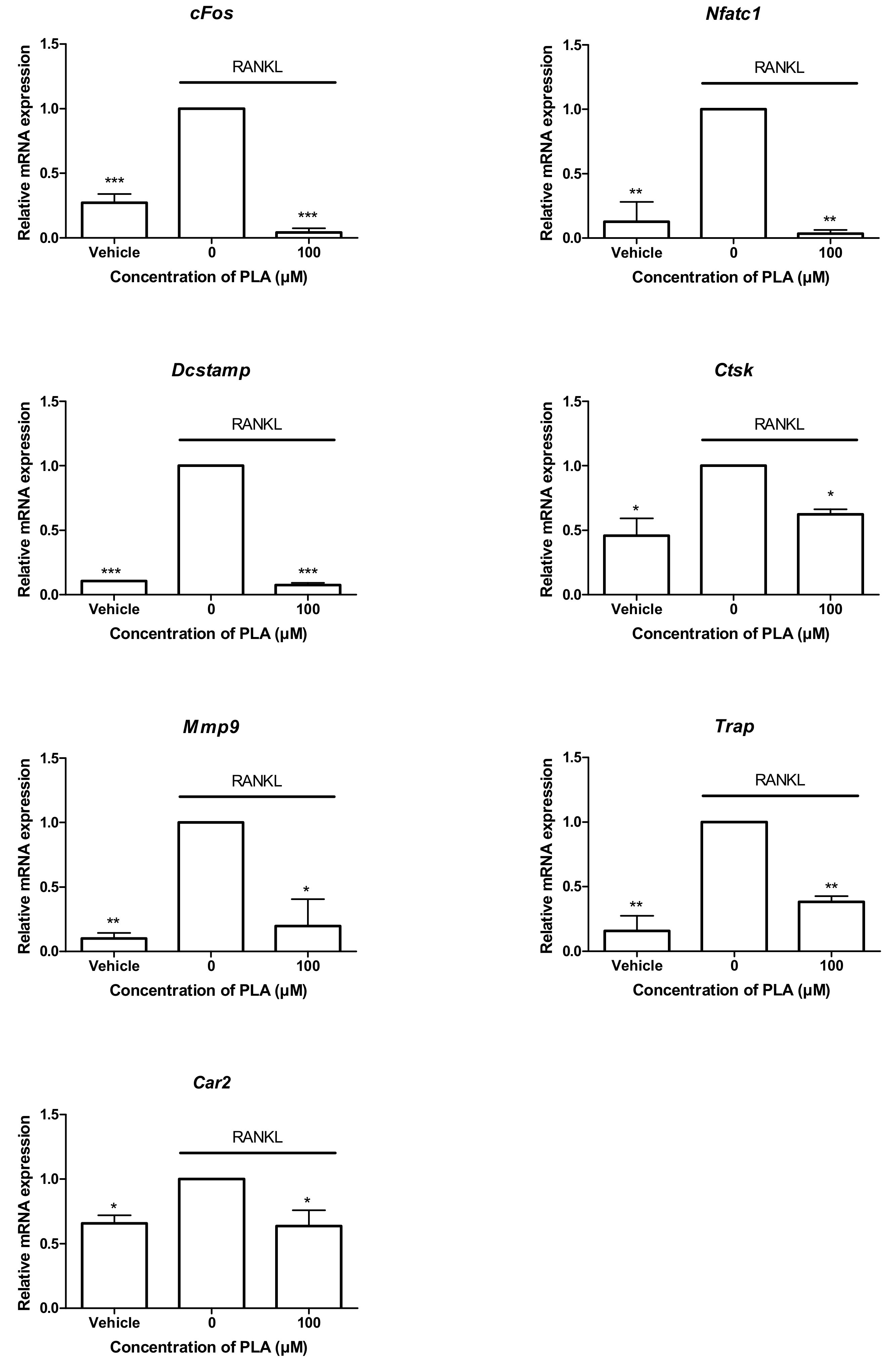
3.5. PLA Suppresses the Expression of Osteoclast-Specific Gene Expression
3.6. PLA Inhibits RANKL-Induced Activation of NF-κB and MAPK Pathways
3.7. Induction of Apoptosis by PLA in Mature Osteoclasts
4. Discussion
5. Conclusions
Acknowledgments
Author contributions
Conflicts of interest
References
- Raggatt, L.J.; Partridge, N.C. Cellular and molecular mechanisms of bone remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [PubMed]
- Blair, H.C.; Athanasou, N.A. Recent advances in osteoclast biology and pathological bone resorption. Histol. Histopathol. 2004, 19, 189–199. [Google Scholar] [PubMed]
- Bonewald, L.F.; Johnson, M.L. Osteocytes, mechanosensing and wnt signaling. Bone 2008, 42, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Baum, R.; Gravallese, E.M. Impact of inflammation on the osteoblast in rheumatic diseases. Curr. Osteoporos. Rep. 2014, 12, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B. Advances in the regulation of osteoclasts and osteoclast functions. J. Dent. Res. 2013, 92, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shavit, Z. The osteoclast: A multinucleated, hematopoietic-Origin, bone-Resorbing osteoimmune cell. J. Cell. Biochem. 2007, 102, 1130–1139. [Google Scholar] [CrossRef] [PubMed]
- Kasonga, A.E.; Deepak, V.; Kruger, M.C.; Coetzee, M. Arachidonic acid and docosahexaenoic acid suppress osteoclast formation and activity in human cd14+monocytes, in vitro. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Kajarabille, N.; Díaz-Castro, J.; Hijano, S.; López-Frías, M.; López-Aliaga, I.; Ochoa, J.J. A new insight to bone turnover: Role of-3 polyunsaturated fatty acids. Sci. World J. 2013. [Google Scholar] [CrossRef] [PubMed]
- Boeyens, J.C.; Deepak, V.; Chua, W.H.; Kruger, M.C.; Joubert, A.M.; Coetzee, M. Effects of omega3- and omega6- Polyunsaturated fatty acids on rankl-induced osteoclast differentiation of RAW264.7 cells: A comparative in vitro study. Nutrients 2014, 6, 2584–2601. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Ohk, B.; Yoon, H.J.; Kang, W.Y.; Seong, S.J.; Kim, S.-Y.; Yoon, Y.-R. Docosahexaenoic acid signaling attenuates the proliferation and differentiation of bone marrow-Derived osteoclast precursors and promotes apoptosis in mature osteoclasts. Cell Signal. 2017, 29, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Bhattacharya, A.; Fernandes, G. Docosahexaenoic acid is more potent inhibitor of osteoclast differentiation in RAW 264.7 cells than eicosapentaenoic acid. J. Cell. Physiol. 2008, 214, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Drosatos-Tampakaki, Z.; Drosatos, K.; Siegelin, Y.; Gong, S.; Khan, S.; Van Dyke, T.; Goldberg, I.J.; Schulze, P.C.; Schulze-Spate, U. Palmitic acid and DGAT1 deficiency enhance osteoclastogenesis, while oleic acid-induced triglyceride formation prevents it. J. Bone Miner. Res. 2014, 29, 1183–1195. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Li, H.; Xu, H.; Halim, V.; Zhang, W.; Wang, H.; Ong, K.T.; Woo, S.-L.; Walzem, R.L.; Mashek, D.G. Palmitoleate induces hepatic steatosis but suppresses liver inflammatory response in mice. PLoS ONE 2012, 7, e39286. [Google Scholar] [CrossRef] [PubMed]
- Nestel, P.; Clifton, P.; Noakes, M. Effects of increasing dietary palmitoleic acid compared with palmitic and oleic acids on plasma lipids of hypercholesterolemic men. J. Lipid Res. 1994, 35, 656–662. [Google Scholar] [PubMed]
- Burns, T.; Duckett, S.; Pratt, S.; Jenkins, T. Supplemental palmitoleic (C16:1 cis-9) acid reduces lipogenesis and desaturation in bovine adipocyte cultures. J. Anim. Sci. 2012, 90, 3433–3441. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, N.; Watson, M.; Sakamoto, K.; Hundal, H.S. Differential effects of palmitate and palmitoleate on insulin action and glucose utilization in rat l6 skeletal muscle cells. Biochem. J. 2006, 399, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-H.; Miyahara, H.; Hatanaka, A. Chronic administration of palmitoleic acid reduces insulin resistance and hepatic lipid accumulation in kk-A y mice with genetic type 2 diabetes. Lipids Health Dis. 2011, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Kantartzis, K.; Celebi, N.; Staiger, H.; Machann, J.; Schick, F.; Cegan, A.; Elcnerova, M.; Schleicher, E.; Fritsche, A. Circulating palmitoleate strongly and independently predicts insulin sensitivity in humans. Diabetes Care 2010, 33, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Fabbrini, E.; Magkos, F.; Su, X.O.; Abumrad, N.A.; Nejedly, N.; Coughlin, C.C.; Okunade, A.L.; Patterson, B.W.; Klein, S. Insulin sensitivity is not associated with palmitoleate availability in obese humans. J. Lipid Res. 2011, 52, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Cao, H.M.; King, I.B.; Lemaitre, R.N.; Song, X.L.; Siscovick, D.S.; Hotamisligil, G.S. Circulating palmitoleic acid and risk of metabolic abnormalities and new-Onset diabetes. Am. J. Clin. Nutr. 2010, 92, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Deepak, V.; Kasonga, A.; Kruger, M.C.; Coetzee, M. Inhibitory effects of eugenol on rankl-induced osteoclast formation via attenuation of NF-κB and mapk pathways. Connect Tissue Res. 2015, 56, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Visagie, A.; Kasonga, A.; Deepak, V.; Moosa, S.; Marais, S.; Kruger, M.C.; Coetzee, M. Commercial honeybush (cyclopia spp.) tea extract inhibits osteoclast formation and bone resorption in RAW264. 7 murine macrophages—an in vitro study. Int. J. Environ. Res. Public Health 2015, 12, 13779–13793. [Google Scholar] [CrossRef] [PubMed]
- Rasband, W. Imagej Software; National Institutes of Health: Bethesda, MD, USA, 1997; Volume 2012. [Google Scholar]
- Deepak, V.; Kasonga, A.; Kruger, M.C.; Coetzee, M. Carvacrol inhibits osteoclastogenesis and negatively regulates the survival of mature osteoclasts. Biol. Pharm. Bull. 2016. [Google Scholar] [CrossRef] [PubMed]
- Burd, J.; Usategui-Gomez, M. A colorimetric assay for serum lactate dehydrogenase. Clin. Chim. Acta 1973, 46, 223–227. [Google Scholar] [CrossRef]
- Chan, F.K.-M.; Moriwaki, K.; De Rosa, M.J. Detection of necrosis by release of lactate dehydrogenase activity. Methods Mol. Biol. 2013, 979, 65–70. [Google Scholar] [PubMed]
- Shomali, T.; Rezaian, M.; Rassouli, A.; Asadi, F. Effect of eicosapentaenoic acid on bone changes due to ethylprednisolone in rats. Basic. Clin. Pharm. Toxicol. 2009, 105, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Chan, D.; Felix, E.; Cartwright, C.; Menter, D.G.; Madden, T.; Klein, R.D.; Fischer, S.M.; Newman, R.A. Formation and antiproliferative effect of prostaglandin E3 from eicosapentanoic acid in human lung cancer cells. J. Lipid Res. 2004, 45, 1030–1039. [Google Scholar] [CrossRef] [PubMed]
- Hӧgstrom, M.; Nӧrdstrom, P.; Nӧrdstrom, A. N-3 fatty acids are positively associated with peak bone mineral density and bone accrual in healthy men: The NO2 study. Am. J. Clin. Nutr. 2007, 85, 803–807. [Google Scholar]
- Shimizu, T.; Yokotani, K. Effects of centrally administered prostaglandin E3 and thromboxane A3 on plasma noradrenaline and adrenaline in rats: Comparison with prostaglandin E2 and thromboxane A2. Eur. J. Pharm. 2009, 611, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Redlich, K.; Smolen, J.S. Inflammatory bone loss: Pathogenesis and therapeutic intervention. Nat. Rev. Drug Discov. 2012, 11, 234–250. [Google Scholar] [CrossRef] [PubMed]
- Collin-Osdoby, P.; Osdoby, P. Rankl-Mediated osteoclast formation from murine RAW 264.7 cells. Bone Res. Protoc. Second Ed. 2012, 816, 187–202. [Google Scholar]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lee, S.H.; Kim, J.H.; Choi, Y.; Kim, N. NFATc1 induces osteoclast fusion via up-Regulation of Atp6v0d2 and the dendritic cell-Specific transmembrane protein (DC-STAMP). Mol. Endocrinol. 2008, 22, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Nakashima, T.; Hiroshi, N.; Penninger, J.M. Rankl-Rank signaling in osteoclastogenesis and bone disease. Trends Mol. Med. 2006, 12, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.P.; Chen, D.; Boyce, B.F. Mice deficient in NF-κB p50 and p52 or rank have defective growth plate formation and post-Natal dwarfism. Bone Res. 2013, 1, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Sudo, T.; Saito, T.; Osada, A.; Tsujimoto, M. Involvement of p38 mitogen-Activated protein kinase signaling pathway in osteoclastogenesis mediated by receptor activator of NF-κB ligand (RANKL). J. Biol. Chem. 2000, 275, 31155–31161. [Google Scholar] [CrossRef] [PubMed]
- David, J.P.; Sabapathy, K.; Hoffmann, O.; Idarraga, M.H.; Wagner, E.F. Jnk1 modulates osteoclastogenesis through both c-Jun phosphorylation-Dependent and independent mechanisms. J. Cell Sci. 2002, 115, 4317–4325. [Google Scholar] [CrossRef] [PubMed]
- Hutchins-Wiese, H.L.; Picho, K.; Watkins, B.A.; Li, Y.; Tannenbaum, S.; Claffey, K.; Kenny, A.M. High-Dose eicosapentaenoic acid and docosahexaenoic acid supplementation reduces bone resorption in postmenopausal breast cancer survivors on aromatase inhibitors: A pilot study. Nut. Cancer Int. J. 2014, 66, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Saraste, A.; Pulkki, K. Morphologic and biochemical hallmarks of apoptosis. Cardiovasc. Res. 2000, 45, 528–537. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′–3′) | Reverse Primer Sequence (5′–3′) |
|---|---|---|
| Gapdh | GATGACATCAAGAAGGTGGTGAAGC | ATACCAGGAAATGAGCTTGACAAAG |
| Mmp9 | GTCATCCAGTTTGGTGTCGCG | AGGGGAAGACGCACAGCTC |
| Ctsk | CTGGAGGGCCAACTCAAGA | CCTCTGCATTTAGCTGCCTT |
| Trap | CAGCTGTCCTGGCTCAA | GTAGGCAGTGACCCCGT |
| cFos | CCCATCGCAGACCAGAGC | ATCTTGCAGGCAGGTCGGT |
| Dcstamp | ATGACTTGCAACCTAAGGGCAAAG | GTCTGGTTCCAAGAAACAAGGTCAT |
| Nfatc1 | GTGGAGAAGCAGAGCAC | ACGCTGGTACTGGCTTC |
| Car2 | GAGTTTGATGACTCTCAGGACAA | CATATTTGGTGTTCCAGTGAACCA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Heerden, B.; Kasonga, A.; Kruger, M.C.; Coetzee, M. Palmitoleic Acid Inhibits RANKL-Induced Osteoclastogenesis and Bone Resorption by Suppressing NF-κB and MAPK Signalling Pathways. Nutrients 2017, 9, 441. https://doi.org/10.3390/nu9050441
Van Heerden B, Kasonga A, Kruger MC, Coetzee M. Palmitoleic Acid Inhibits RANKL-Induced Osteoclastogenesis and Bone Resorption by Suppressing NF-κB and MAPK Signalling Pathways. Nutrients. 2017; 9(5):441. https://doi.org/10.3390/nu9050441
Chicago/Turabian StyleVan Heerden, Bernadette, Abe Kasonga, Marlena C. Kruger, and Magdalena Coetzee. 2017. "Palmitoleic Acid Inhibits RANKL-Induced Osteoclastogenesis and Bone Resorption by Suppressing NF-κB and MAPK Signalling Pathways" Nutrients 9, no. 5: 441. https://doi.org/10.3390/nu9050441
APA StyleVan Heerden, B., Kasonga, A., Kruger, M. C., & Coetzee, M. (2017). Palmitoleic Acid Inhibits RANKL-Induced Osteoclastogenesis and Bone Resorption by Suppressing NF-κB and MAPK Signalling Pathways. Nutrients, 9(5), 441. https://doi.org/10.3390/nu9050441