Prevalence of Inherited Hemoglobin Disorders and Relationships with Anemia and Micronutrient Status among Children in Yaoundé and Douala, Cameroon

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Data Collection
2.3. Laboratory Analyses
2.4. Data Analysis
3. Results
3.1. Household and Participant Characteristics
3.2. Prevalence of Inherited Hemoglobin Disorders
3.3. Relationships between Inherited Hemoglobin Disorders and Anemia
3.4. Predictors of Anemia
3.5. Relationships between Inherited Hemoglobin Disorders and Micronutrient Status
3.6. Interpretation of Soluble Transferrin Receptor
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stevens, G.; Finucane, M.; De-Regil, L.; Paciorek, C.; Flaxman, S.; Branca, F.; Peña-Rosas, J.; Bhutta, Z.; Ezzati, M.; Nutrition Impact Model Study Group (Anaemia). Global, regional, and national trends in haemoglobin concentration and prevalence of total and severe anaemia in children and pregnant and non-pregnant women for 1995–2011: A systematic analysis of population-representative data. Lancet Glob Health 2013, 1, e16–e25. [Google Scholar] [PubMed]
- Brabin, B.; Premji, Z.; Verhoeff, F. An analysis of anemia and child mortality. J. Nutr. 2001, 131, 636S–645S. [Google Scholar] [PubMed]
- Northrop-Clewes, C.; Thurnham, D.I. Biomarkers for the differentiation of anemia and their clinical usefulness. J. Blood Med. 2013, 4, 11–22. [Google Scholar] [PubMed]
- Foote, E.; Sullivan, K.; Ruth, L.J.; Oremo, J.; Sadumah, I.; Williams, T.N.; Suchdev, P.S. Determinants of anemia among preschool children in rural, western Kenya. Am. J. Trop. Med. Hyg. 2013, 88, 757–764. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization; UNICEF; United Nations University. Iron Deficiency Anemia: Assessment, Prevention, and Control. A Guide for Programme Managers; World Health Organization: Geneva, Switzerland, 2001. [Google Scholar]
- World Health Organization. Conclusions and recommendations of the who consultation on prevention and control of iron deficiency in infants and young children in malaria-endemic areas. Food Nutr. Bull. 2007, 28, S621–S627. [Google Scholar]
- Sazawal, S.; Black, R.; Ramsan, M.; Chwaya, H.; Stoltzfus, R.J.; Dutta, A.; Dhingra, U.; Kabole, I.; Deb, S.; Othman, M.; et al. Effects of routine prophylactic supplementation with iron and folic acid on admission to hospital and mortality in preschool children in a high malaria transmission setting: Community-based, randomized, placebo-controlled trial. Lancet 2006, 367, 133–143. [Google Scholar] [CrossRef]
- Piel, F.; Weatherall, D. The α-thalassemias. N. Engl. J. Med. 2014, 371, 1908–1916. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Parobek, C.; Fairhurst, R. Haemoglobinopathies and the clinical epidemiology of malaria: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 457–468. [Google Scholar] [CrossRef]
- Elguero, E.; Délicat-Loembet, L.; Rougeron, V.; Arnathau, C.; Roche, B.; Becquart, P.; Gonzalez, J.; Nkoghe, D.; Sica, L.; Leroy, E.; et al. Malaria continues to select for sickle cell trait in central africa. Proc. Natl. Acad. Sci. USA 2015, 112, 7051–7054. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.N.; Mwangi, T.; Wambua, S.; Peto, T.; Weatherall, D.; Gupta, S.; Recker, M.; Penman, B.; Uyoga, S.; Macharia, A.; et al. Negative epistasis between the malaria-protective effects of alpha+-thalassemia and the sickle cell trait. Nat. Genet. 2005, 37, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Piel, F.; Hay, S.; Gupta, S.; Weatherall, D.; Williams, T.N. Global burden of sickle cell anaemia in children under five, 2010–2050: Modelling based on demographics, excess mortality, and interventions. PLoS Med. 2013, 10. [Google Scholar] [CrossRef] [PubMed]
- Modell, B.; Darlison, M. Global epidemiology of haemoglobin disorders and derived service indicators. Bull. World Health Org. 2008, 86, 480–487. [Google Scholar] [CrossRef] [PubMed]
- WHO-TIF. Management of Haemoglobin Disorders: Report of a Joint Who-TIF Meeting, Nicosia, Cyprus, 16–18 November 2007; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Nyakeriga, A.; Troye-Blomberg, M.; Mwacharo, J.; Wambua, S.; Williams, T.N. Nutritional iron status in children with a+thalassemia and the sickle cell trait in a malaria endemic area on the coast of Kenya. Hematologica 2005, 90, 552–554. [Google Scholar]
- Tsang, B.; Sullivan, K.M.; Ruth, L.J.; Williams, T.N.; Suchdev, P.S. Nutritional status of young children with inherited blood disorders in western Kenya. Am. J. Trop. Med. Hyg. 2014, 90, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Skikne, B.S.; Flowers, C.H.; Cook, J.D. Serum transferrin receptor: A quantitative measure of tissue iron deficiency. Blood 1990, 75, 1870–1876. [Google Scholar] [CrossRef] [PubMed]
- Beguin, Y.; Clemons, G.; Pootrakul, P.; Fillet, G. Quantitative assessment of erythropoiesis and functional classification of anemia based on measurements of serum transferrin receptor and erythropoietin. Blood 1993, 81, 1067–1076. [Google Scholar] [PubMed]
- Verhoef, H.; West, C.E.; Ndeto, P.; Burema, J.; Beguin, Y.; Kok, F.J. Serum transferrin receptor concentration indicates increased erythropoiesis in Kenyan children with asymptomatic malaria. Am. J. Clin. Nutr. 2001, 74, 767–775. [Google Scholar] [PubMed]
- Engle-Stone, R.; Nankap, M.; Ndjebayi, A.O.; Gimou, M.-M.; Friedman, A.; Haskell, M.; Tarini, A.; Brown, K.H. Vitamin a status of women and children in yaoundé and douala, cameroon, is unchanged one year after initiation of a national vitamin a oil fortification program. Nutrients 2017, 9, 522. [Google Scholar] [CrossRef] [PubMed]
- Engle-Stone, R.; Nankap, M.; Ndjebayi, A.O.; Allen, L.H.; Shahab-Ferdows, S.; Hampel, D.; Killilea, D.; Gimou, M.-M.; Houghton, L.; Friedman, A.; et al. Iron, zinc, folate, and vitamin b-12 status increased among women and children in yaoundé and douala, cameroon, one year after introducing fortified wheat flour. J. Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Helen Keller International Cameroon. Micronutrient Content of Fortified Foods, and Micronutrient Status of Women and Young Children in Douala and Yaoundé, Cameroon, One Year Following Fortification of Wheat Flour and Refined Vegetable Oil; Helen Keller International: Yaoundé, Cameroon, 2015. [Google Scholar]
- International Zinc Nutrition Consultative Group (IZiNCG). Assessment of the risk of zinc deficiency in populations and options for its control. Hotz c, brown kh, eds. Food Nutr. Bull. 2004, 25 (Suppl. S2), S91–S204. [Google Scholar]
- Erhardt, J.G.; Estes, J.E.; Pfeiffer, C.M.; Biesalski, H.K.; Craft, N.E. Combined measurement of ferritin, soluble transferrin receptor, retinol binding protein, and c-reactive protein by an inexpensive, sensitive, and simple sandwich enzyme-linked immunosorbent assay technique. J. Nutr. 2004, 134, 3127–3132. [Google Scholar] [PubMed]
- Shahab-Ferdows, S.; Engle-Stone, R.; Hampel, D.; Ndjebayi, A.; Nankap, M.; Brown, K.; Allen, L. Regional, socioeconomic, and dietary risk factors for vitamin b-12 deficiency differ from those for folate deficiency in cameroonian women and children. J. Nutr. 2015, 55, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Zyba, S.; Shenvi, S.; Killilea, D.; Holland, T.; Kim, E.; Moy, A.; Sutherland, B.; Gildengorin, V.; Shigenaga, M.; King, J.C. A moderate increase in diet zinc reduced DNA strand breaks in leukocytes and altered plasma proteins without changing plasma zinc concentrations. Am. J. Clin. Nutr. 2017, 105, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Chong, S.; Boehm, C.; Higgs, D.; Cutting, G. Single-tube multiplex-pcr screen for common deletional determinants of alpha-thalassemia. Blood 2000, 95, 360–362. [Google Scholar] [PubMed]
- Engle-Stone, R.; Nankap, M.; Ndjebayi, A.O.; Erhardt, J.G.; Brown, K.H. Plasma ferritin and soluble transferrin receptor concentrations and body iron stores identify similar risk factors for iron deficiency but result in different estimates of the national prevalence of iron deficiency and iron-deficiency anemia among women and children in Cameroon. J. Nutr. 2013, 143, 369–377. [Google Scholar] [PubMed]
- Suchdev, P.S.; Namaste, S.; Aaron, G.; Raiten, D.J.; Brown, K.H.; Flores Ayala, R.; BRINDA Working Group. Overview of the biomarkers reflecting inflammation and nutritional determinants of anemia (brinda) project. Adv. Nutr. 2016, 7, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.D.; Flowers, C.H.; Skikne, B.S. The quantitative assessment of body iron. Blood 2003, 101, 3359–3364. [Google Scholar] [CrossRef] [PubMed]
- Engle-Stone, R.; Haskell, M.J.; Ndjebayi, A.O.; Nankap, M.; Erhardt, J.G.; Gimou, M.-M.; Brown, K.H. Plasma retinol-binding protein predicts plasma retinol concentration in both infected and uninfected cameroonian women and children. J. Nutr. 2011, 141, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Haemoglobin Concentrations for the Diagnosis of Anaemia and Assessment of Severity; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Institut National de la Statistique (INS) et ICF International. Enquête Démographique et de Santé et à Indicateurs Multiples du Cameroun 2011; INS et ICF International: Calverton, MD, USA, 2012. [Google Scholar]
- Weatherall, D.; Clegg, J.; Blankson, J.; McNeil, J. A new sickling disorder resulting from interaction of the genes for haemoglobin s and alpha-thalassemia. Br. J. Haematol. 1969, 17, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Le Hesran, J.-Y.; Personne, I.; Personne, P.; Fievet, N.; Dubois, B.; Beyemé, M.; Boudin, C.; Cot, M.; Deloron, P. Longitudinal study of plasmodium falciparum infection and immune responses in infants with or without the sickle cell trait. Int. J. Epidemiol. 1999, 28, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Suchdev, P.S.; Ruth, L.J.; Earley, M.; Macharia, A.; Williams, T.N. The burden and consequences of inherited blood disorders among young children in western Kenya. Matern. Child Nutr. 2014, 10, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Yip, R.; Johnson, C.; Dallman, P.R. Age-related changes in laboratory values used in the diagnosis of anemia and iron deficiency. Am. J. Clin. Nutr. 1984, 39, 427–436. [Google Scholar] [PubMed]
- Domellöf, M.; Dewey, K.; Lönnerdal, B.; Cohen, R.; Hernell, O. The diagnostic criteria for iron deficiency in infants should be reevaluated. J. Nutr. 2002, 132, 3680–3686. [Google Scholar] [PubMed]
- Grant, F.K.E.; Martorell, R.; Flores-Ayala, R.; Cole, C.R.; Ruth, L.J.; Ramakrishnan, U.; Suchdev, P.S. Comparison of indicators of iron deficiency in Kenyan children. Am. J. Clin. Nutr. 2012, 95, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Wessells, K.; Hess, S.Y.; Ouédraogo, Z.; Rouamba, N.; Ouédraogo, J.; Brown, K.H. Asymptomatic malaria infection affects the interpretation of biomarkers of iron and vitamin a status, even after adjusting for systemic inflammation, but does not affect plasma zinc concentrations among young children in burkina faso. J. Nutr. 2014, 144, 2050–2058. [Google Scholar] [CrossRef] [PubMed]
- Righetti, A.; Wegmüller, R.; Glinz, D.; Ouattara, M.; Adiossan, L.; N’goran, E.; Utzinger, J.; Hurrell, R. Effects of inflammation and plasmodium falciparum infection on soluble transferrin receptor and plasma ferritin concentration in different age groups: A prospective longitudinal study in côte d’ivoire. Am. J. Clin. Nutr. 2013, 97, 1364–1374. [Google Scholar] [CrossRef] [PubMed]
- Mei, Z.; Pfeiffer, C.M.; Looker, A.; Flores-Ayala, R.; Lacher, D.A.; Mirel, L.; Grummer-Strawn, L. Serum soluble transferrin receptor concentrations in us preschool children and non-pregnant women of childbearing age from the national health and nutrition examination survey 2003–2010. Clin. Chim. Acta 2012, 413, 1479–1484. [Google Scholar] [CrossRef] [PubMed]
- Chouliaras, G.; Premetis, E.; Tsiftis, G.; Drosatou, P.; Papassotiriou, I.; Stamoulakatou, A.; Lycopoulou, L. Serum transferrin receptors: Distribution and diagnostic performance in pre-school children. Blood Cells Mol. Dis. 2009, 43, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Glinz, D.; Hurrell, R.; Ouattara, M.; Zimmermann, M.B.; Brittenham, G.; Adiossan, L.; Righetti, A.; Seifert, B.; Diakité, V.; Utzinger, J.; et al. The effect of iron-fortified complementary food and intermitten preventive treatment of malaria on anaemia in 12- to 36-month-old children; a cluster-randomised controlled trial. Malar. J. 2015, 14. [Google Scholar] [CrossRef] [PubMed]


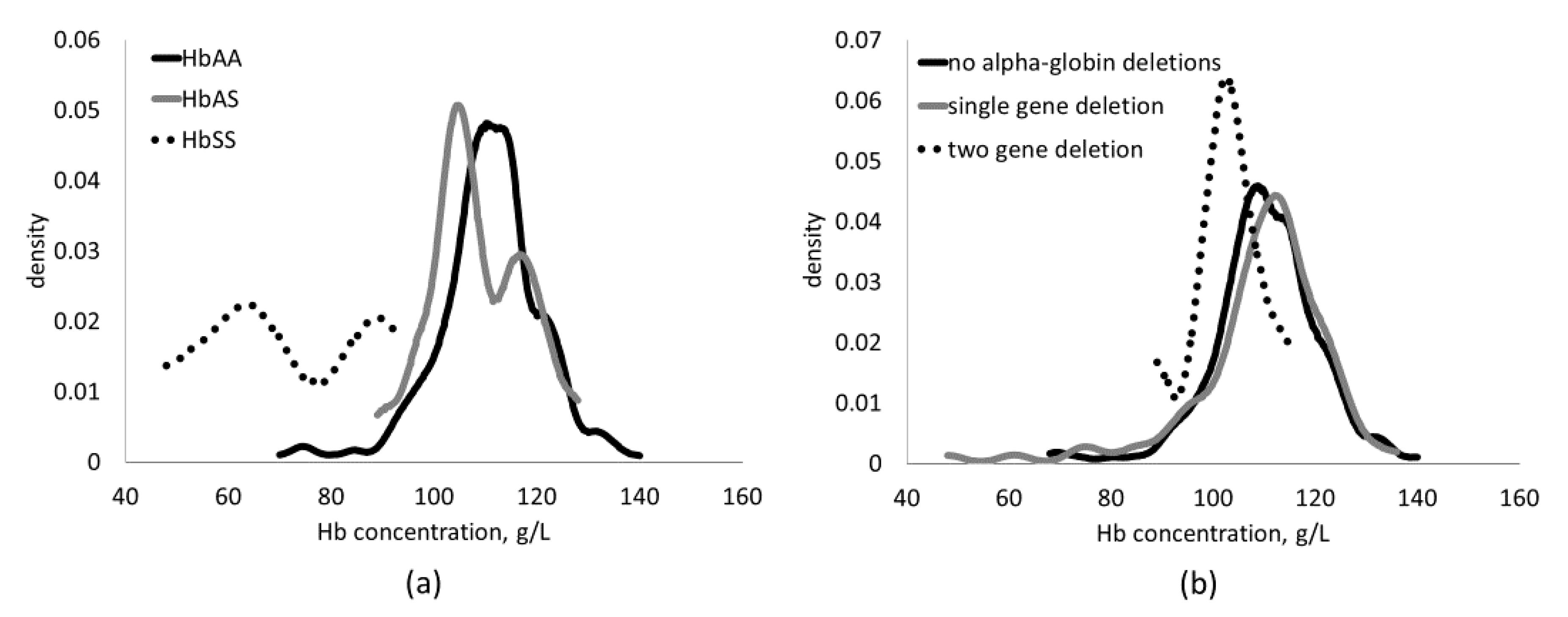{kind=link}
| Variable | Value |
|---|---|
| Age, months | 32.8 ± 0.8 |
| Male, n (%) | 155 (50) |
| Breastfeeding, n (%) | 11 (4) |
| Stunted (HAZ < −2), % | 15.6 ± 2.4 |
| Wasted (WHZ < −2), % | 1.4 ± 0.7 |
| Inflammation (CRP > 5 mg/L and/or AGP > 1 g/L), % | 46.4 ± 3.1 |
| Malaria, % | 8.0 ± 2.1 |
| Hemoglobin, g/L | 110 ± 1 |
| Anemia (Hb < 110 g/L), % | 45.4 ± 2.9 |
| Severe anemia (Hb < 70 g/L), % | 1.0 ± 0.5 |
| Reticulocytes, cells/µL | 47,000 ± 2000 |
| Reticulocytes, cells/µL 2 | 39,000 (29,000, 54,000) 2 |
| Reticulocytes > 150,000 cells/µL, n (%) | 5 (1.7) |
| Unadjusted ferritin, µg/L | 51.1 ± 2.8 |
| Adjusted ferritin 3, µg/L | 32.2 ± 1.2 |
| Adjusted Ferritin 3 < 12 µg/L, % | 13.2 ± 2.0 |
| sTfR, mg/L | 8.22 ± 0.20 |
| sTfR > 8.3 mg/L, % | 25.4 ± 2.3 |
| Unadjusted RBP, µmol/L | 0.88 ± 0.02 |
| Adjusted RBP 3, µmol/L | 1.06 ± 0.01 |
| Adjusted RBP 3 < 0.83 umol/L, % | 13.5 ± 1.9 |
| Plasma zinc, µg/dL | 66.8 ± 1.6 |
| Adjusted plasma zinc 3, µg/dL | 74.0 ± 1.7 |
| Low adjusted plasma zinc 3,4, % | 23.2 ± 3.8 |
| Folate, nmol/L | 56.0 ± 2.1 |
| Plasma B12, pmol/L | 851 ± 51 |
| Urban Total | Yaoundé | Douala | ||
|---|---|---|---|---|
| Any Hb S or alpha-globin 2 deletion | 42.9 (37.4–48.4) | 48.9 (39.7–58.1) | 37.1 (30.1–44.0) | |
| Sickle cell disease or trait 3 | ||||
| HbSS | 1.6 (0.2–3.0) | 2.7 (0.1–5.3) | 0.6 (0–1.9) | |
| HbAS | 13.7 (9.7–17.8) | 12.6 (6.3–18.9) | 14.8 (9.1–20.5) | |
| α+-thalassemia 2 | ||||
| Homozygous (–α/–α) | 3.1 (1.1–5.2) | 4.9 (1.2–8.6) | 1.4 (0–3.5) | |
| Heterozygous (–α/αα) | 30.6 (25.4–35.8) | 37.0 (27.7–46.4) | 24.4 (18.9–29.9) | |
| Other Hb conditions 3 | ||||
| Increased F | 4.7 (2.7–6.6) | 5.5 (2.5–8.4) | 3.9 (1.2–6.7) |
| Independent Variables | Dependent Variable: Hb Concentration (g/L2) | Dependent Variable: Anemia (Hb < 110 g/L) | ||||||
|---|---|---|---|---|---|---|---|---|
| Bivariate Relationships | Adjusted Model | Bivariate Relationships | Adjusted Model | |||||
| β (SE) | p | β (SE) | p 2 | OR (95% CI) | p | OR (95% CI) | p 2 | |
| Age, months | 25.4 (14.4) | 0.088 | 24.7 (11.0) | 0.033 | 0.972 (0.949–0.997 | 0.026 | 0.963 (0.938–0.990) | 0.007 |
| Male | −598 (331) | 0.080 | -- | 0.25 | 0.76 (0.45–1.28) | 0.30 | -- | 0.48 |
| Hb AS | −138 (368) | 0.71 | -- | 0.15 | 2.01 (1.01–4.02) | 0.047 | 2.14 (1.13–4.07) | 0.020 |
| Hb SS | −6960 (1020) | <0.0001 | −6790 (880) | <0.0001 | >999 (>999–>999) 5 | <0.0001 | >999 (>999–>999) | <0.001 |
| α+thalassemia heterozygote | −149 (341) | 0.67 | -- | 0.31 | 0.81 (0.46–1.42) | 0.47 | -- | 0.71 |
| α+thalassemia homozygote | −1470 (490) | 0.006 | −1270 (440) | 0.007 | 4.31 (1.03–17.93) | 0.045 | -- | 0.11 |
| Malaria | −2050 (690) | 0.006 | −1760 (620) | 0.008 | 2.62 (1.10–6.27) | 0.030 | 2.80 (1.03–7.64) | 0.045 |
| Adjusted RBP 3 < 0.83 µmol/L | −1360 (430) | 0.004 | −620 (299) | 0.048 | 2.00 (1.09–3.67) | 0.026 | -- | 0.13 |
| Adjusted ferritin 3 < 12 µg/L | −1270 (310) | 0.0003 | −1170 (330) | 0.001 | 2.46 (1.38–4.41) | 0.002 | 2.08 (1.03–4.17) | 0.040 |
| Low adjusted plasma zinc 3,4 | −425 (399) | 0.30 | -- | 0.99 | 1.26 (0.68–2.32) | 0.46 | -- | 0.90 |
| Plasma CRP, mg/L | −334 (129) | 0.015 | -- | 0.71 | 1.05 (1.01–1.09) | 0.015 | 1.05 (1.01–1.09) | 0.014 |
| Plasma AGP, g/L | −1520 (550) | 0.010 | −1290 (470) | 0.010 | 2.46 (1.16–5.21) | 0.019 | -- | 0.44 |
| Plasma folate, nmol/L | 18.4 (8.1) | 0.03 | -- | 0.27 | 0.975 (0.962–0.989) | 0.0003 | 0.976 (0.961–0.991) | 0.002 |
| Plasma vitamin B12, pmol/L | 1.45 (0.38) | 0.0007 | 0.984 (0.312) | 0.004 | 0.999 (0.998–1.000) | 0.012 | -- | 0.078 |
| sTfR ≤ 8.3 mg/L | sTfR > 8.3 mg/L | p | |
|---|---|---|---|
| n | 211–221 | 72–76 | |
| Age, months | 34.3 ± 1.0 | 29.9 ± 1.5 | 0.014 |
| Male, % | 46.6 ± 3.2 | 64.1 ± 5.9 | 0.012 |
| Hemoglobin, g/L | 112 ± 1 | 102 ± 2 | <0.001 |
| Anemic, % | 37.3 ± 3.1 | 69.4 ± 4.5 | <0.001 |
| Malaria, % | 5.1 ± 1.7 | 18.9 ± 5.5 | 0.0005 |
| Reticulocytes, cells/nL | 38.8 ± 1.6 | 68.4 ± 6.3 | <0.001 |
| Reticulocytes > 150 cells/nL, % | 0 | 6.8 ± 2.9 | <0.001 |
| Inflammation (CRP > 5 mg/L and/or AGP > 1 g/L) | 42.1 ± 3.1 | 59.3 ± 5.6 | 0.002 |
| CRP, mg/L | 4.23 ± 0.49 | 5.31 ± 1.01 | 0.31 |
| AGP, mg/L | 0.94 ± 0.02 | 1.06 ± 0.04 | 0.003 |
| Adjusted ferritin 2 < 12 µg/dL, % | 6.3 ± 1.6 | 33.5 ± 6.8 | <0.001 |
| Adjusted RBP 2 < 0.83 µmol/L, % | 11.7 ± 2.2 | 18.5 ± 4.7 | 0.18 |
| Low adjusted zinc 2,3, % | 21.5 ± 3.8 | 25.8 ± 5.8 | 0.38 |
| Plasma folate, nmol/L | 56.7 ± 2.4 | 54.3 ± 2.7 | 0.44 |
| Plasma vitamin B12, pmol/L | 828 ± 30 | 778 ± 84 | 0.54 |
| HbAS genotype | 13.0 ± 2.1 | 16.2 ± 4.8 | 0.52 |
| HbSS genotype | 0 | 6.3 ± 2.7 | <0.001 |
| α+thalassemia heterozygote | 31.8 ± 2.9 | 27.7 ± 4.5 | 0.44 |
| α+thalassemia homozygote | 2.4 ± 1.0 | 5.4 ± 2.4 | 0.17 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Engle-Stone, R.; Williams, T.N.; Nankap, M.; Ndjebayi, A.; Gimou, M.-M.; Oyono, Y.; Tarini, A.; Brown, K.H.; Green, R. Prevalence of Inherited Hemoglobin Disorders and Relationships with Anemia and Micronutrient Status among Children in Yaoundé and Douala, Cameroon. Nutrients 2017, 9, 693. https://doi.org/10.3390/nu9070693
Engle-Stone R, Williams TN, Nankap M, Ndjebayi A, Gimou M-M, Oyono Y, Tarini A, Brown KH, Green R. Prevalence of Inherited Hemoglobin Disorders and Relationships with Anemia and Micronutrient Status among Children in Yaoundé and Douala, Cameroon. Nutrients. 2017; 9(7):693. https://doi.org/10.3390/nu9070693
Chicago/Turabian StyleEngle-Stone, Reina, Thomas N. Williams, Martin Nankap, Alex Ndjebayi, Marie-Madeleine Gimou, Yannick Oyono, Ann Tarini, Kenneth H. Brown, and Ralph Green. 2017. "Prevalence of Inherited Hemoglobin Disorders and Relationships with Anemia and Micronutrient Status among Children in Yaoundé and Douala, Cameroon" Nutrients 9, no. 7: 693. https://doi.org/10.3390/nu9070693
APA StyleEngle-Stone, R., Williams, T. N., Nankap, M., Ndjebayi, A., Gimou, M. -M., Oyono, Y., Tarini, A., Brown, K. H., & Green, R. (2017). Prevalence of Inherited Hemoglobin Disorders and Relationships with Anemia and Micronutrient Status among Children in Yaoundé and Douala, Cameroon. Nutrients, 9(7), 693. https://doi.org/10.3390/nu9070693