The Impact of Warming and Nutrients on Algae Production and Microcystins in Seston from the Iconic Lake Lesser Prespa, Greece

 , ,
, , 
Abstract
:1. Introduction
2. Results
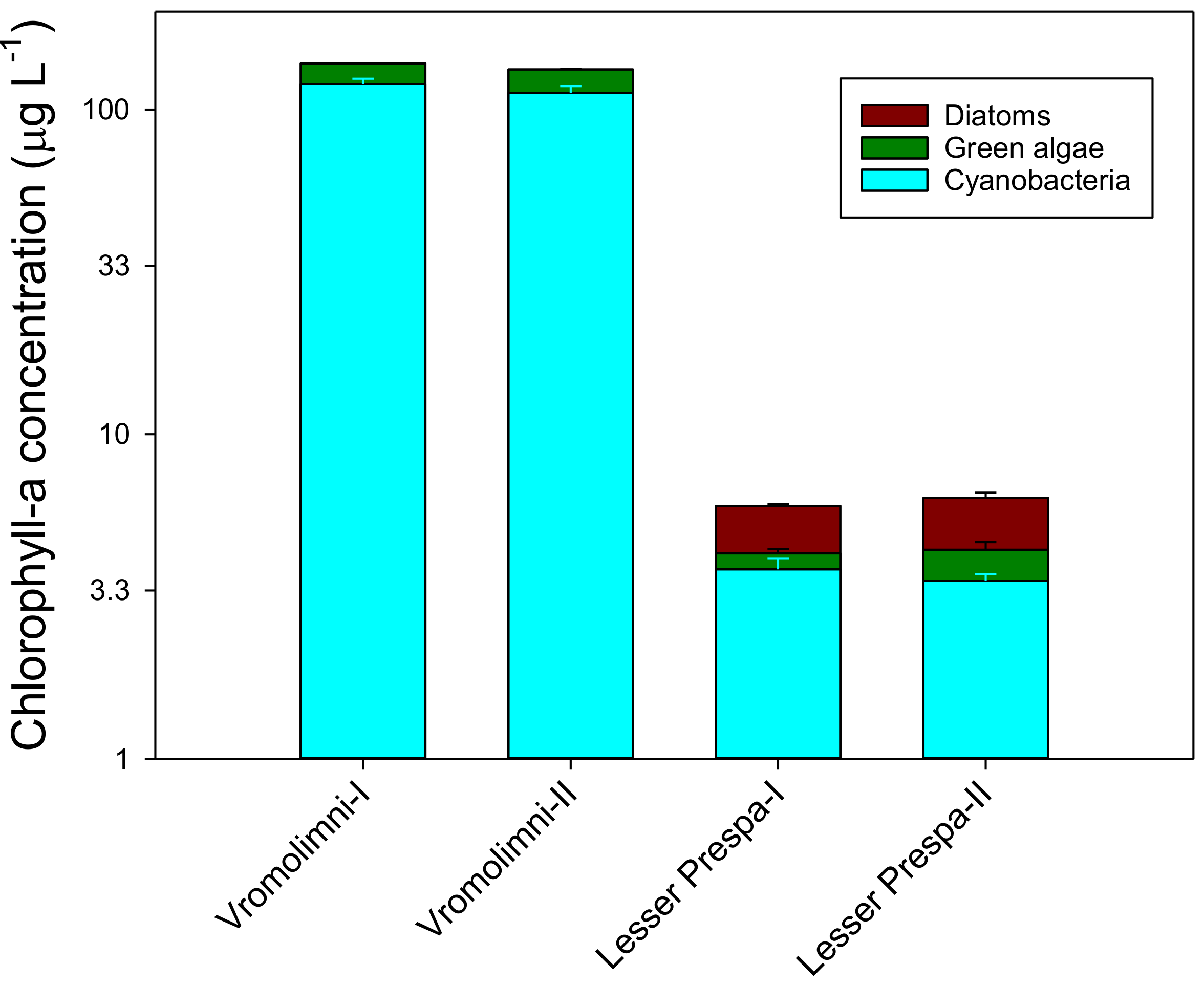
2.1. Initial Chlorophyll-a, Nutrient and MC Concentrations in Water from Lake Lesser Prespa and Vromolimni Pond
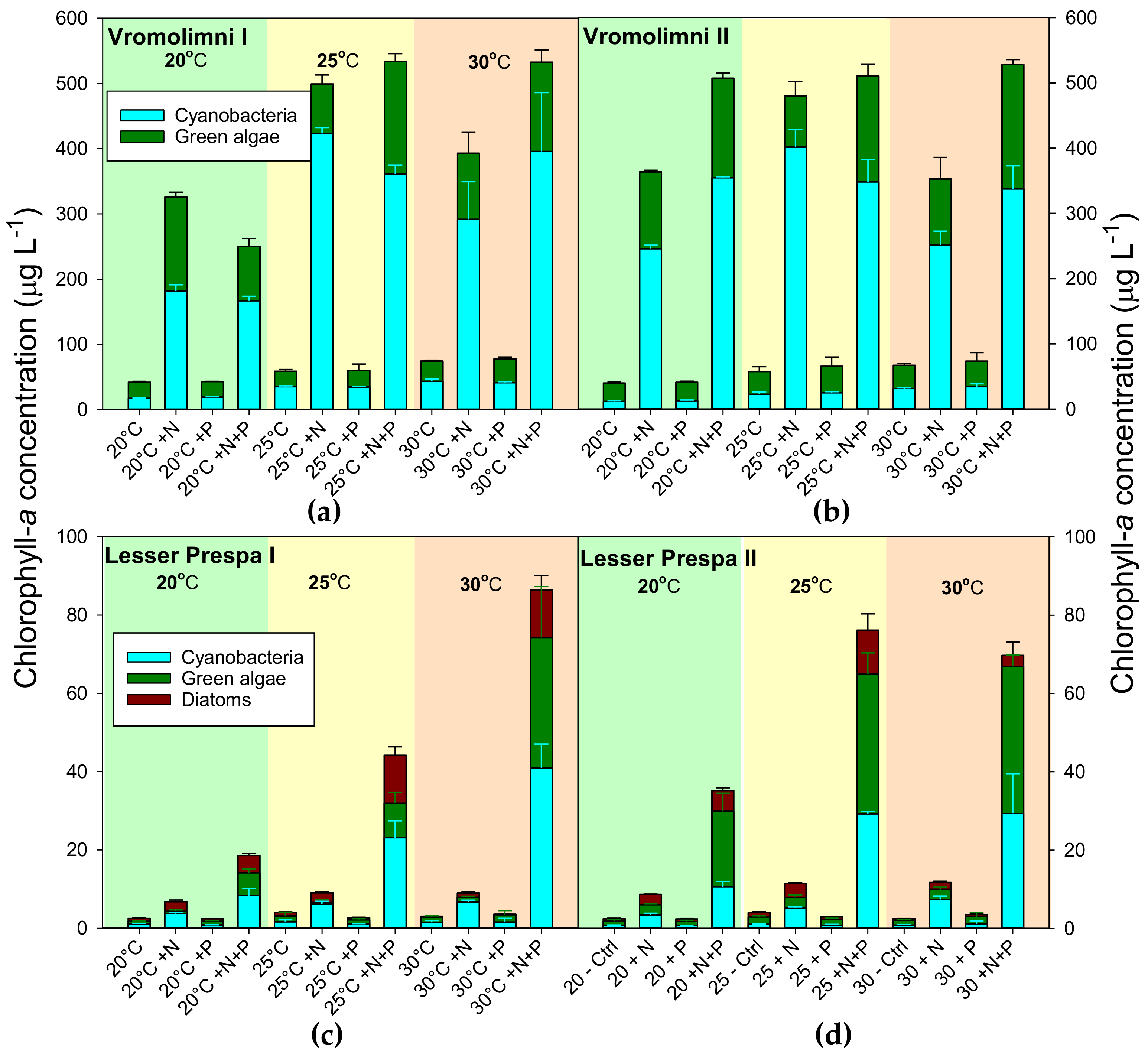
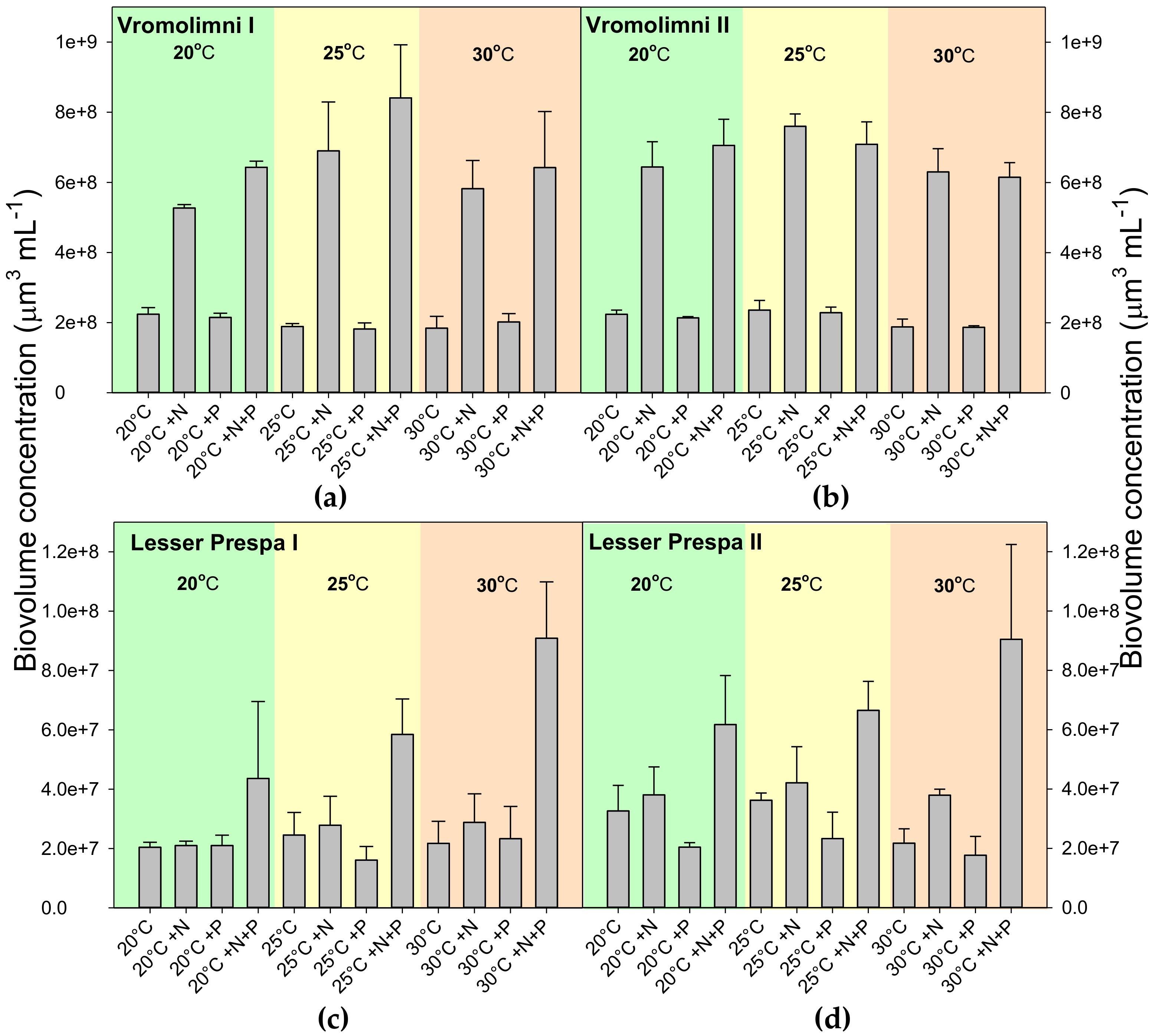
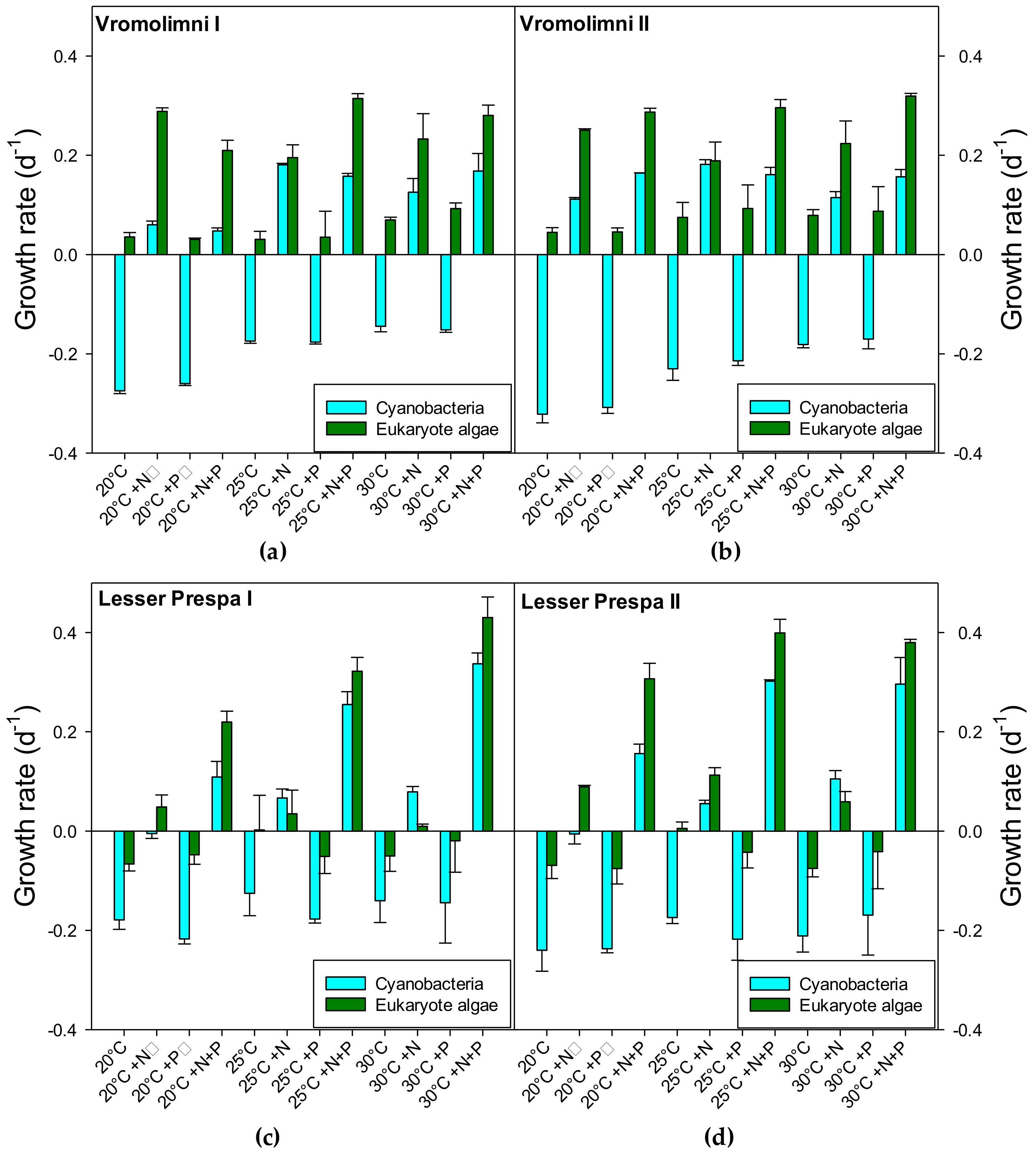
2.2. Final Chlorophyll-a and Biovolume Concentrations
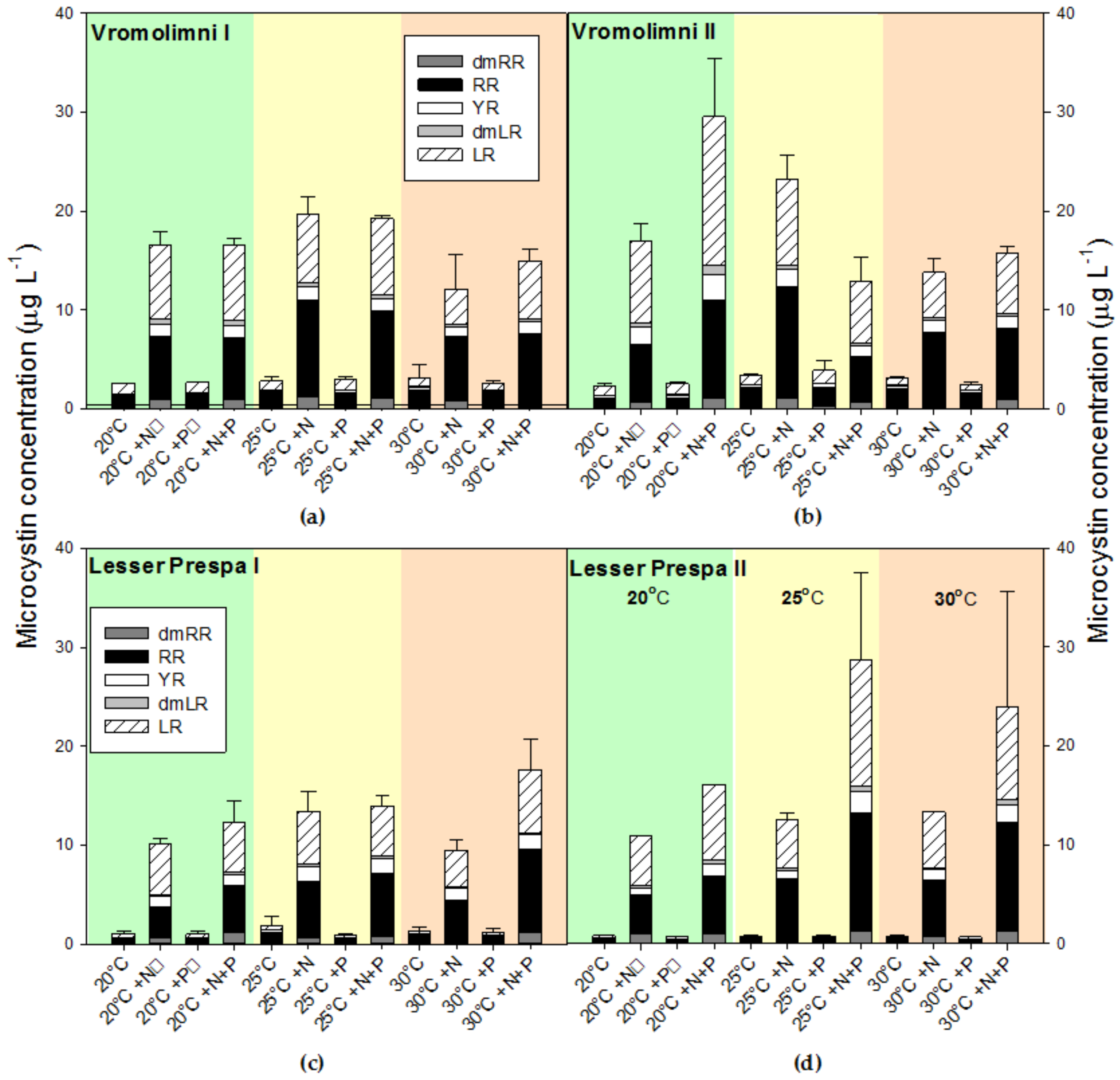
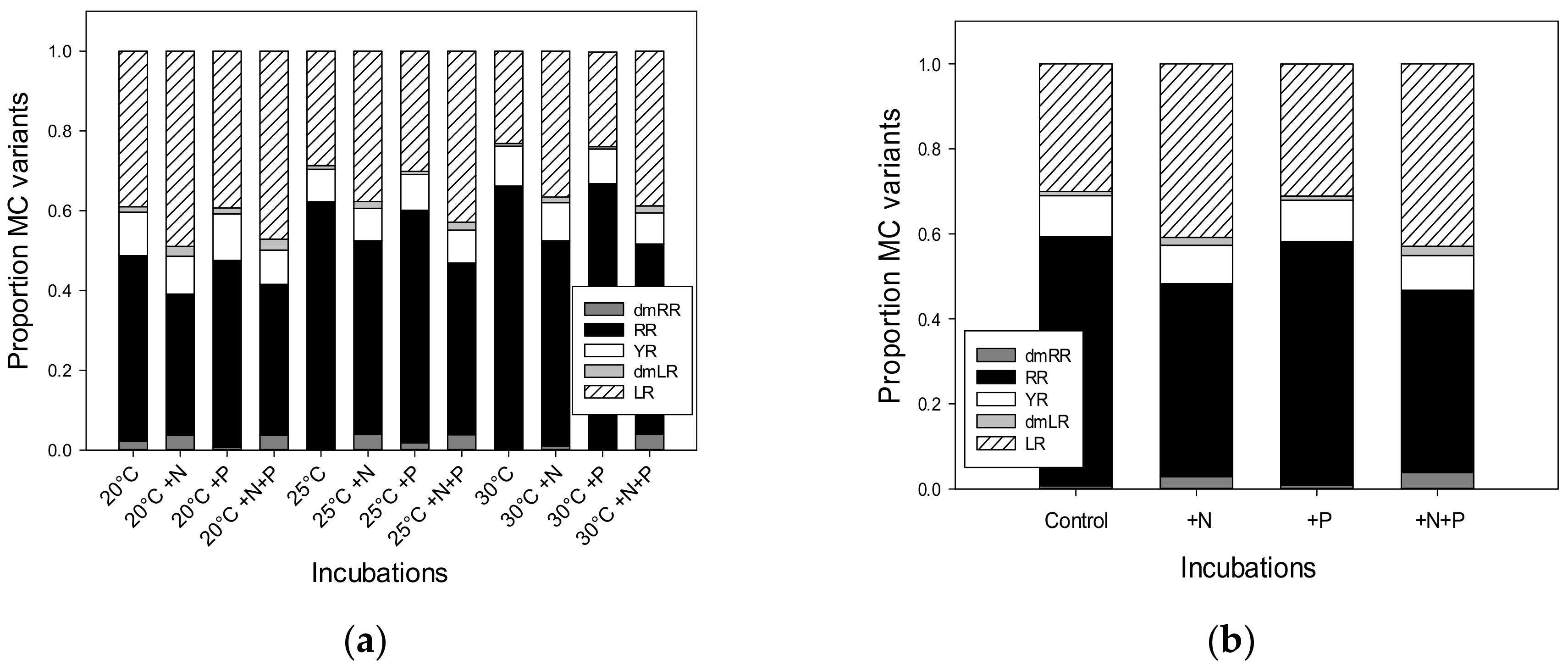
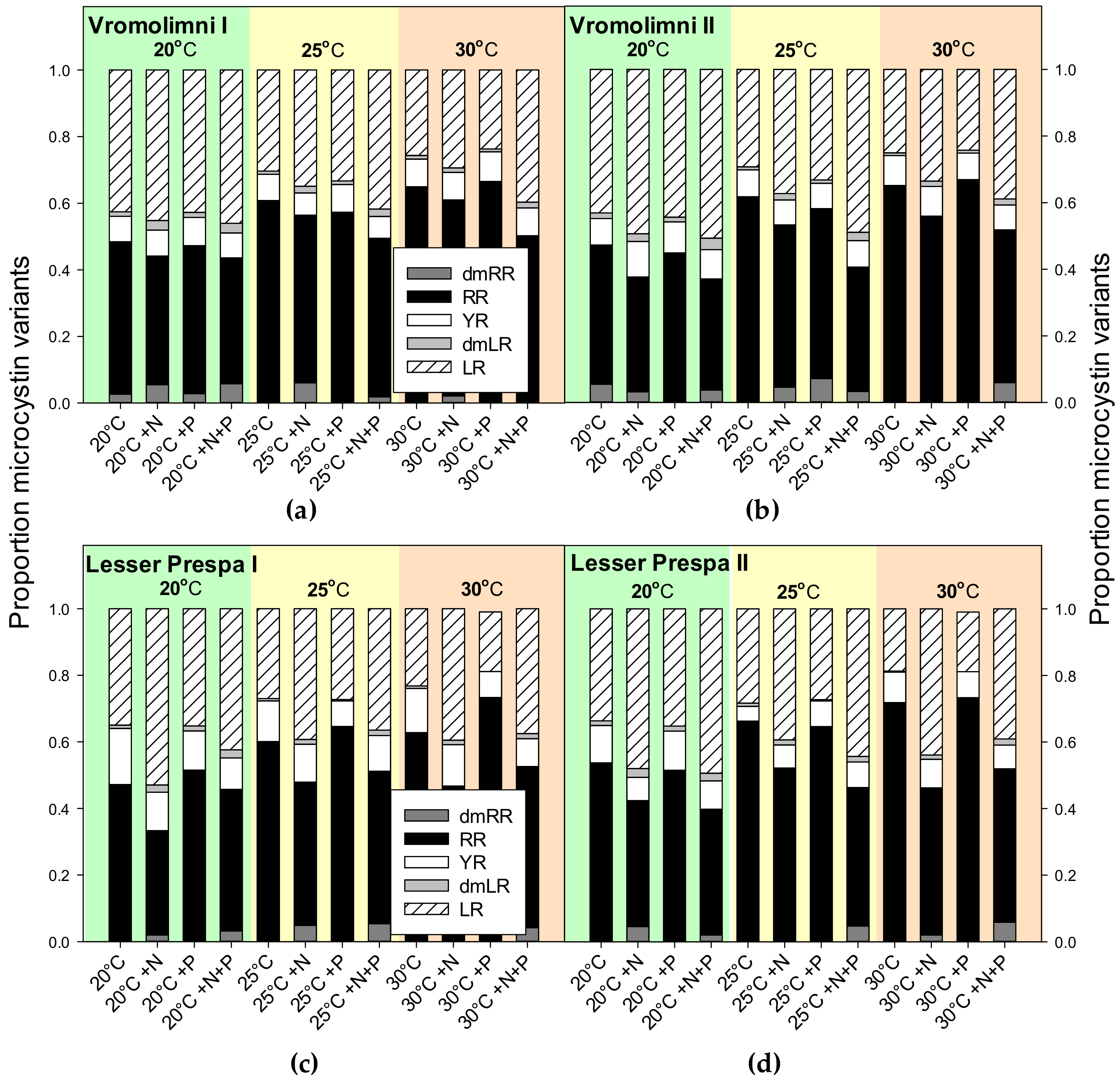
2.3. Final Microcystin Concentrations
3. Discussion
4. Conclusions
5. Materials and Methods
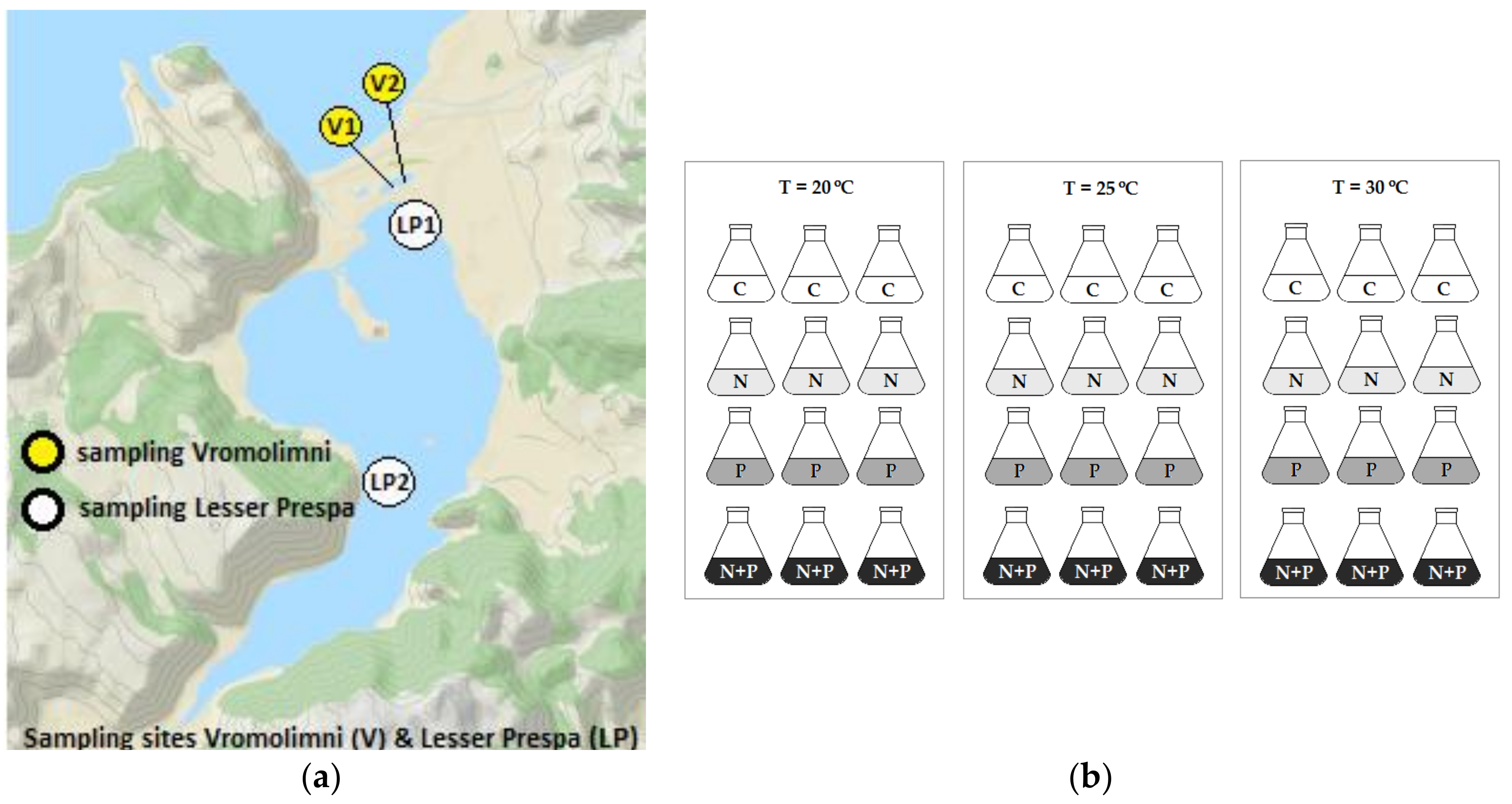
5.1. Field Sampling
5.2. Algal Growth Assay
5.3. Microcystin and Nodularin Analysis Using LC-MS/MS
5.4. Data Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

Appendix B

Appendix C

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | LOD1 (µg L−1) | LOQ2 (µg L−1) |
|---|---|---|
| dm-7-MC-RR | 0.253 | 0.253 |
| MC-RR | 0.187 | 0.187 |
| NOD | <0.017 | 0.017 |
| MC-YR | <0.024 | 0.024 |
| dm-7-MC-LR | <0.023 | 0.023 |
| MC-LR | <0.045 | 0.045 |
| MC-LY | 0.395 | 0.395 |
| MC-LW | 0.269 | 0.538 |
| MC-LF | 0.113 | 0.113 |
References
- Crivelli, A.J.; Catsadorakis, G. Lake Prespa, North-western Greece: A unique Balkan Wetland. Hydrobiologia 1997, 351, 1–196. [Google Scholar] [CrossRef]
- BirdLife International. Species Factsheet: Pelecanus Crispus. 2017. Available online: http://www.birdlife.org (accessed on 9 December 2017).
- De Groot, R.S.; Alkemade, R.; Braat, L.; Hein, L.; Willemen, L. Challenges in integrating the concept of ecosystem services and values in landscape planning, management and decision making. Ecol. Complex. 2010, 7, 260–272. [Google Scholar] [CrossRef]
- Verstijnen, Y.; Maliaka, V.; Catsadorakis, G.; Smolders, A.J.P.; Lürling, M. Nutrient contribution by four colonial-nesting waterbirds to Lake Lesser Prespa (Greece) and their influence on the water quality. In preparation. 2018.
- Maliaka, V.; Lürling, M.; Fritz, C.; Faassen, E.; van Oosterhout, F.; Smolders, A.J.P. Cyanotoxins in the Prespa Lake Area, Greece. Toxins in preparation. 2018.
- van der Schriek, T.; Giannakopoulos, C. Determining the causes for the dramatic recent fall of Lake Prespa (southwest Balkans). Hydrol. Sci. J. 2017, 62, 1131–1148. [Google Scholar] [CrossRef]
- Watson, S.B.; McCauley, E.; Downing, J.A. Patterns in phytoplankton taxonomic composition across temperate lakes of differing nutrient status. Limnol. Oceanogr. 1997, 42, 487–495. [Google Scholar] [CrossRef]
- Smith, V.H.; Tilman, G.D.; Nekola, J.C. Eutrophication: Impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environ. Pollut. 1999, 100, 179–196. [Google Scholar] [CrossRef]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Moss, B.; Kosten, S.; Meerhoff, M.; Battarbee, R.W.; Jeppesen, E.; Mazzeo, N.; Havens, K.; Lacerot, G.; Liu, Z.; De Meester, L.; et al. Allied attack: Climate change and eutrophication. Inland Waters 2011, 1, 101–105. [Google Scholar] [CrossRef]
- Trolle, D.; Nielsen, A.; Rolighed, J.; Thodsen, H.; Andersen, H.E.; Karlsson, I.B.; Refsgaard, J.C.; Olesen, J.E.; Bolding, K.; Kronvang, B.; et al. Projecting the future ecological state of lakes in Denmark in a 6 degree warming scenario. Clim. Res. 2015, 64, 55–72. [Google Scholar] [CrossRef]
- Guedes, I.A.; Moreira da Costa Leite, D.; Alves Manhães, L.; Mascarello Bisch, P.; Azevedo, S.M.F.O.; Pacheco, A.B.F. Fluctuations in microcystin concentrations, potentially toxic Microcystis and genotype diversity in a cyanobacterial community from a tropical reservoir. Harmful Algae 2014, 39, 303–309. [Google Scholar] [CrossRef]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Pacheco, A.B.F.; Guedes, I.A.; Azevedo, S.M.F.O. Is qPCR a reliable indicator of cyanotoxin risk in freshwater? Toxins 2016, 8, 172. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; van Oosterhout, F.; Faassen, E. Eutrophication and warming boost cyanobacterial biomass and microcystins. Toxins 2017, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Kosten, S.; Huszar, V.L.M.; Mazzeo, N.; Scheffer, M.; Sternberg, L.D.S.L.; Jeppesen, E. Lake and watershed characteristics rather than climate influence nutrient limitation in shallow lakes. Ecol. Appl. 2009, 19, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.S. Cyanobacterial Water-Blooms. Adv. Bot. Res. 1987, 13, 67–143. [Google Scholar]
- Ger, K.A.; Urrutia-Cordero, P.; Frost, P.C.; Hansson, L.A.; Sarnelle, O.; Wilson, A.E.; Lürling, M. The interaction between cyanobacteria and zooplankton in a more eutrophic world. Harmful Algae 2016, 54, 128–144. [Google Scholar] [CrossRef] [PubMed]
- Salomon, E.; Bar-Eyal, L.; Sharon, S.; Keren, N. Balancing photosynthetic electron flow is critical for cyanobacterial acclimation to nitrogen limitation. Biochim. Biophys. Acta 2013, 1827, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.M.; Smith, A.J. Nitrogen chlorosis in blue-green algae. Arch. Mikrobiol. 1969, 69, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Harke, M.J.; Gobler, C.J. Daily transcriptome changes reveal the role of nitrogen in controlling microcystin synthesis and nutrient transport in the toxic cyanobacterium, Microcystis aeruginosa. BMC Genom. 2015, 16, 1068. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, J.S.M.; Giani, A. Microcystin production and regulation under nutrient stress conditions in toxic Microcystis strains. Appl. Environ. Microbiol. 2014, 80, 5836–5843. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Falconer, I.R.; Salas, H.J.; Bartram, J. Health risks caused by freshwater cyanobacteria in recreational waters. J. Toxicol. Environ. Health Part B 2000, 3, 323–347. [Google Scholar]
- Rapala, J.; Sivonen, K. Assessment of environmental conditions that favor hepatotoxic and neurotoxic Anabaena spp. strains cultured under light limitation at different temperatures. Microb. Ecol. 1998, 36, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Rapala, J.; Sivonen, K.; Lyra, C.; Niemela, S.I. Variation of microcystin, cyanobacterial hepatotoxins, in Anabaena spp. as a function of growth stimulation. App. Environ. Microbiol. 1997, 63, 2206–2212. [Google Scholar]
- Mowe, M.A.D.; Porojan, C.; Abbas, F.; Mitrovic, S.M.; Lim, R.P.; Furey, A.; Yeo, D.C.J. Rising temperatures may increase growth rates and microcystin production in tropical Microcystis species. Harmful Algae 2015, 50, 88–98. [Google Scholar] [CrossRef]
- Sivonen, K.; Namikoshi, M.; Evans, W.R.; Carmichael, W.W.; Sun, F.; Rouhiainen, L.; Luukkainen, R.; Rinehart, K.L. Isolation and characterization of a variety of microcystins from seven strains of the cyanobacterial genus Anabaena. Appl. Environ. Microbiol. 1992, 58, 2495–2500. [Google Scholar] [PubMed]
- Van de Waal, D.B.; Ferreruela, G.; Tonk, L.; Van Donk, E.; Huisman, J.; Visser, P.M.; Matthijs, H.C.P. Pulsednitrogen supply induces dynamic changes in the amino acid composition and microcystin production of the harmful cyanobacterium Planktothrix agardhii. FEMS Microbiol. Ecol. 2010, 74, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, D. Competition between toxic Microcystis aeruginosa and nontoxic Microcystis wesenbergii with Anabaena PCC7120. J. Appl. Phycol. 2012, 24, 69–78. [Google Scholar] [CrossRef]
- Zurawell, R.W.; Chen, H.; Burke, J.M.; Prepas, E.E. Hepatotoxic cyanobacteria: A review of the biological importance of microcystins in freshwater environments. J. Toxicol. Environ. Health Part B 2005, 8, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Mischler, J.A.; Taylor, P.G.; Townsend, A.R. Nitrogen Limitation of Pond Ecosystems on the Plains of Eastern Colorado. PLoS ONE 2014, 9, e95757. [Google Scholar] [CrossRef] [PubMed]
- Redfield, A.C. The biological control of chemical factors in the environment. Am. Sci. 1958, 46, 205–221. [Google Scholar]
- Gersberg, R.M.; Elkins, B.V.; Lyon, S.R.; Goldman, C.R. Role of aquatic plants in wastewater treatment by artificial wetlands. Water Res. 1986, 20, 363–368. [Google Scholar] [CrossRef]
- Lewis, W.M.; Wurtsbaugh, W.A.; Paerl, H.W. Rationale for control of anthropogenic nitrogen and phosphorus in inland waters. Environ. Sci. Technol. 2011, 45, 10030–10035. [Google Scholar] [CrossRef] [PubMed]
- Catsadorakis, G.; Onmuş, O.; Bugariu, S.; Gül, O.; Hatzilacou, D.; Hatzofe, O.; Malakou, M.; Michev, T.; Naziridis, T.; Nikolaou, H.; et al. Current status of the Dalmatian pelican and the great white pelican populations of the Black Sea/Mediterranean flyway. Endanger. Species Res. 2015, 27, 119–130. [Google Scholar] [CrossRef]
- Lürling, M.; Mackay, E.; Reitzel, K.; Spears, B.M. Editorial—A critical perspective on geo-engineering for eutrophication management in lakes. Water Res. 2016, 97, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NEN. Water: Photometric Determination of the Content of Dissolved Orthophosphate and the Total Content of Phosphorous Compounds by Continuous Flow Analysis; NEN 6663; Netherlands Normalization Institute: Delft, The Netherlands, 1986. [Google Scholar]
- NEN. Water: Photometric Determination of the Content of Ammonium Nitrogen and the Sum of the Contents of Ammoniacal and Organically Bound Nitrogen According to Kjeldahl by Continuous Flow Analysis; NEN 6646; Netherlands Normalization Institute: Delft, The Netherlands, 1990. [Google Scholar]
- NEN. Bepaling van Het Stikstofgehalte in de Vorm van Nitriet en in de Vorm van Nitraat en de Som van Beide Met Doorstroomanalyse (CFA en FIA) en Spectrometrische Detectie; NEN-EN-ISO 13395; Netherlands Normalization Institute: Delft, The Netherlands, 1997. (In Dutch) [Google Scholar]
- Maliaka, V. Data retrieved from the Project ‘Eutrophication Assessment of Prespa Lakes with the Use of Biogeochemical Indicators (2013–2017)’. Society for the Protection of Prespa—Greece, Radboud University Nijmegen, Wageningen University—The Netherlands. Unpublished data.
- Society for the Protection of Prespa. Unpublished data. 2012.
- Lürling, M.; Faassen, E.J. Dog poisonings associated with a Microcystis aeruginosa bloom in the Netherlands. Toxins 2013, 5, 556–567. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maliaka, V.; Faassen, E.J.; Smolders, A.J.P.; Lürling, M. The Impact of Warming and Nutrients on Algae Production and Microcystins in Seston from the Iconic Lake Lesser Prespa, Greece. Toxins 2018, 10, 144. https://doi.org/10.3390/toxins10040144
Maliaka V, Faassen EJ, Smolders AJP, Lürling M. The Impact of Warming and Nutrients on Algae Production and Microcystins in Seston from the Iconic Lake Lesser Prespa, Greece. Toxins. 2018; 10(4):144. https://doi.org/10.3390/toxins10040144
Chicago/Turabian StyleMaliaka, Valentini, Elisabeth J. Faassen, Alfons J.P. Smolders, and Miquel Lürling. 2018. "The Impact of Warming and Nutrients on Algae Production and Microcystins in Seston from the Iconic Lake Lesser Prespa, Greece" Toxins 10, no. 4: 144. https://doi.org/10.3390/toxins10040144
APA StyleMaliaka, V., Faassen, E. J., Smolders, A. J. P., & Lürling, M. (2018). The Impact of Warming and Nutrients on Algae Production and Microcystins in Seston from the Iconic Lake Lesser Prespa, Greece. Toxins, 10(4), 144. https://doi.org/10.3390/toxins10040144