Effects of Dietary Astaxanthin Supplementation on Energy Budget and Bioaccumulation in Procambarus clarkii (Girard, 1852) Crayfish under Microcystin-LR Stress

Abstract
:1. Introduction
2. Results
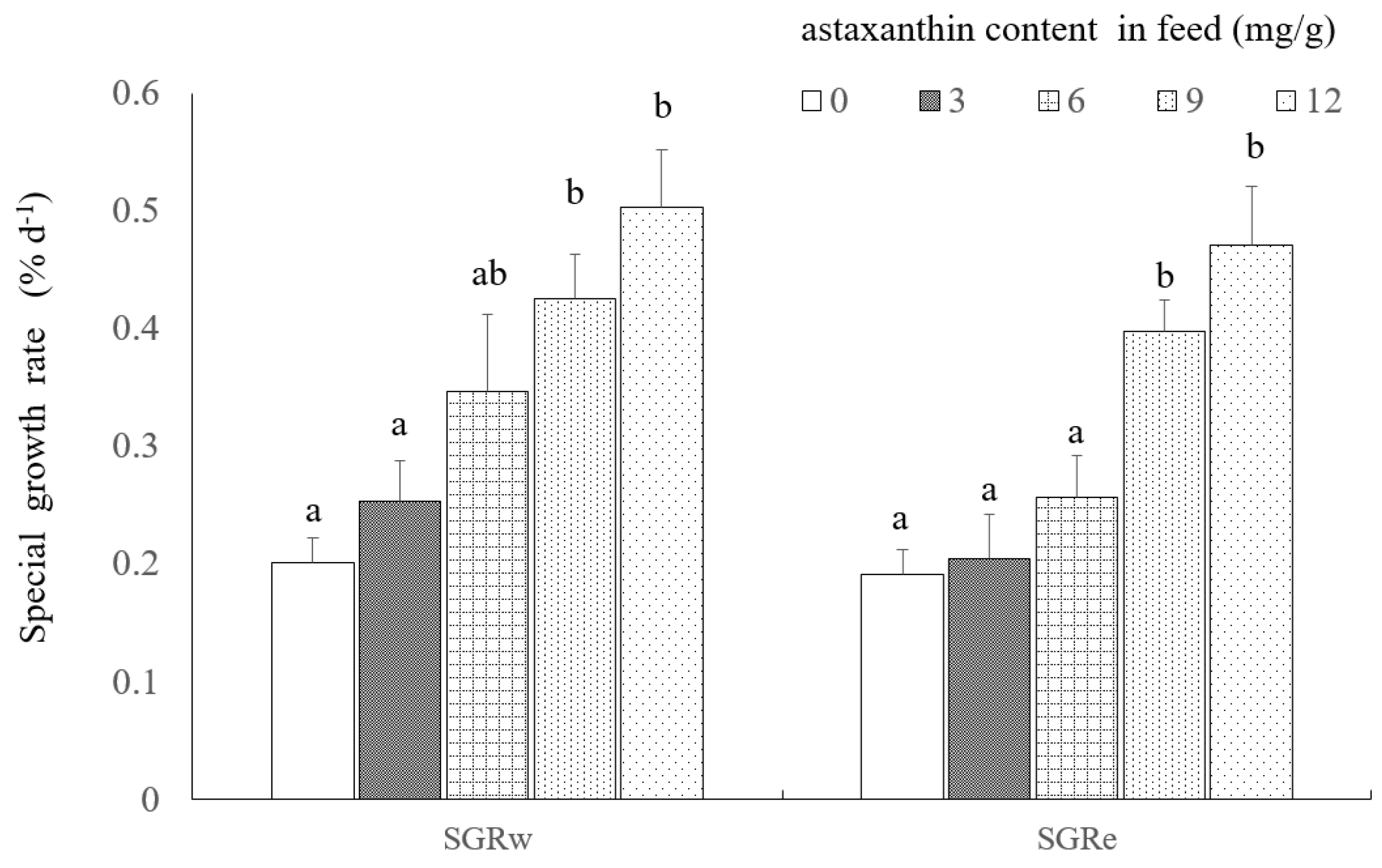
2.1. Growth
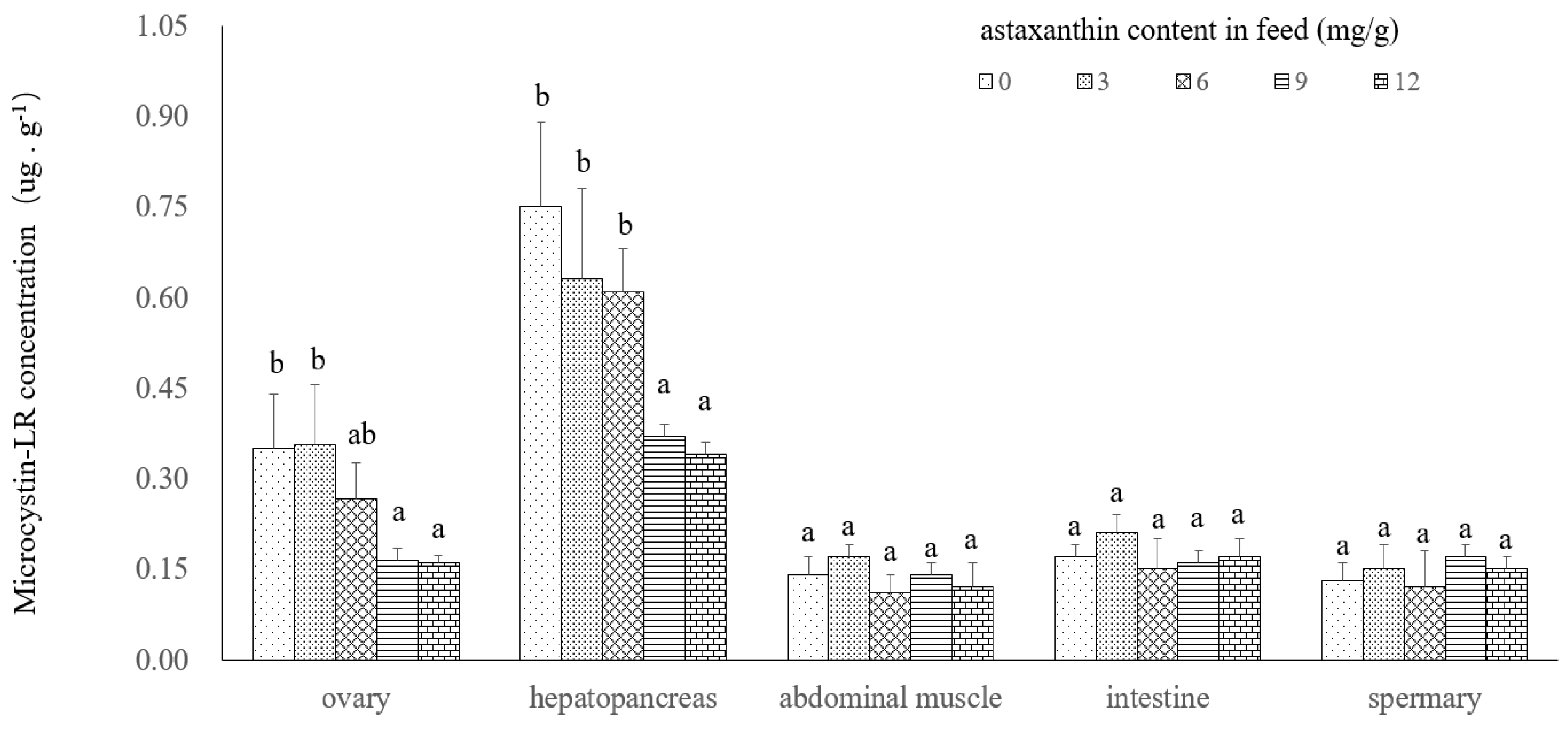
2.2. Microcystin Depuration
2.3. Energy Allocation
3. Discussion
4. Material and Methods
4.1. Source of Animals and Acclimation
4.2. Experimental Design and Procedure
4.4. Energy Determination and Estimation of Energy Budget
4.3. Calculation and Data Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Gherardi, F.; Lazzara, L. Effects of the density of an invasive crayfish (P. clarkii) on a pelagic and surface microalgae in a Mediterranean wetland. Arch. Hydrobiol. 2006, 165, 401–414. [Google Scholar] [CrossRef]
- Carmichael, W.W. Toxic Microcystis and the Environment; Watanabe, M.F., Harada, K., Carmichael, W.W., Fujiki, H., Eds.; CRC Press: Boca Raton, FL, USA, 1996; p. 111. [Google Scholar]
- Vasconcelos, V.; Oliveira, S.; Teles, F.O. Impact of a toxic and non-toxic strain of Microcystis aeruginosa on the crayfish Procambarus clarkii. Toxicon 2001, 39, 1461–1470. [Google Scholar] [CrossRef]
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Kaldre, K.; Haugjärv, K.; Liiva, M.; Gross, R. The effect of two different feeds on growth, carapace colour, maturation and mortality in marbled crayfish (Procambarus fallax f. virginalis). Aquac. Int. 2015, 23, 185–194. [Google Scholar] [CrossRef]
- Wang, W.; Ishikawa, M.; Koshio, S.; Yokoyama, S.; Hossain, M.S.; Moss, A.S. Effects of dietary astaxanthin supplementation on juvenile kuruma shrimp, Marsupenaeus japonicus. Aquaculture 2018, 491, 197–204. [Google Scholar] [CrossRef]
- Berticat, O.; Nègre-Sadargues, G.; Castillo, R. The metabolism of astaxanthin during the embryonic development of the crayfish Astacus leptodactylus Eschscholtz (Crustacea, Astacidea). Comp. Biochem. Physiol. Part B Biochem. 2000, 127, 309–318. [Google Scholar] [CrossRef]
- Xu, H.B.; Sui, H.X.; Gao, S.R. Primary experimental study on bioaccumulation of microcystin in Cyprinus carpio L. Chin. J. Food Hyg. 2003, 15, 202–204. [Google Scholar]
- Li, X.G.; Zhou, G.; Zhou, J.L. Preliminary study on the Accumulation and Biodepuration of Microcystins in Tilapia. J. Hydroecol. 2010, 3, 67–70. [Google Scholar]
- Krishnamurthy, T.; Carmichael, W.W.; Sarver, E.W. Toxic peptides from freshwater cyanobacteria (blue-green-algae). I. Isolation, purification and characterization of peptides from Microcystis aeruginosa and Anabaena flos-aquae. Toxicon 1986, 24, 865–873. [Google Scholar] [CrossRef]
- Hansson, L.A.; Gustafsson, S.; Rengefors, K.; Bomark, L. Cyanobacterial chemical warfare affects zooplankton community composition. Fresh Water Biol. 2007, 52, 1290–1301. [Google Scholar] [CrossRef]
- Codd, G.A.; Poon, G.K. Cyanobacterial toxins. In Biochemistry of the Algae and Cyanobacteria; Rogers, L.J., Gallon, J.R., Eds.; Oxford Science Publishers, Clarendon Press: Oxford, UK, 1998; pp. 283–296. [Google Scholar]
- Chen, J.; Xie, P. Tissue distributions and seasonal dynamics of the hepatotoxic microcystins-LR and-RR in two freshwater shrimps, Palaemon modestus and Macrobrachium nipponensis, from a large shallow, eutrophic lake of the subtropical China. Toxicon 2005, 45, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Haney, J.F. Foodweb transfer, accumulation, and depuration of microcystins, a cyanobacterial toxin, in pumpkinseed sunfish (Lepomis gibbosus). Toxicon 2006, 48, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Elena, T.; Silvia, B.; Sara, B. Depuration of microcystin-LR from the red swamp crayfish Procambarus clarkii with assessment of its food quality. Aquaculture 2008, 285, 90–95. [Google Scholar]
- Huang, C.H.; Chang, R.J.; Huang, S.L.; Chen, W.L. Dietary vitamin E supplementation affects tissue lipid peroxidation of hybrid tilapia. Oreochromis niloticus×O. aureus. Comp. Biochem. Physiol. Part B 2003, 134, 265–270. [Google Scholar] [CrossRef]
- Garciaguerrero, M.; Racotta, I.S.; Villarreal, H. Variation in lipid, protein, and carbohydrate content during the embryonic development of the crayfish Cherax Quadricarinatus (Decapoda: Parastacidae). J. Crustacean Biol. 2003, 23, 1–6. [Google Scholar] [CrossRef]
- Cazenave, J.; Bistoni, M.A.; Pesce, S.F.; Wunderlin, D.A. Differential detoxification and antioxidant response in diverse organs of Corydoras paleatus experimentally exposed to microcystin-RR. Aquatic Toxicol. 2006, 76, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Prieto, A.I.; Pichardo, S.; Jos, A.; Moreno, I.; Camean, A.M. Time-dependent oxidative stress responses after acute exposure to toxic cyanobacterial cells containing microcystins in tilapia fish (Oreochromis niloticus) under laboratory conditions. Aquatic Toxicol. 2007, 84, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Petit, H.; Negre-Sadargues, G.; Castillo, R.; Trilles, J.P. The effects of dietary astaxanthin on growth and moulting cycle of postlarval stages of the prawn, Penaeus Japonicus (Crustacea, Decapoda). Comp. Biochem. Physiol. Part A Physiol. 1997, 117, 539–544. [Google Scholar] [CrossRef]
- Carfoot, T.H. Animal Energetics; Academic Press: New York, NY, USA, 1987; pp. 407–515. [Google Scholar]
- Levine, D.M.; Sulkin, S.D. Partitioning and utilization of energy during the larval development of the xanthid crab, Rithropanopeus harrisii (Gould). J. Exp. Mar. Biol. Ecol. 1979, 40, 247–257. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ax Content mg/g | 0 | 3 | 6 | 9 | 12 |
|---|---|---|---|---|---|
| Survival ratio % | 77.78 ± 4.81 a | 83.33 ± 8.33 ab | 86.11 ± 12.73 a,b | 94.44 ± 4.81 b | 97.22 ± 4.81 b |
| Astaxanthin Content mg/g | C 2 | G 3 | F 4 | E 5 | U 6 | R 7 |
|---|---|---|---|---|---|---|
| 0 | 4304.24 ± 177.48 a | 612.47 ± 95.0 a | 1027.15 ± 105.80 a | 120.56 ± 14.02 a | 182.17 ± 14.02 a | 2361.89 ± 146.96 b |
| 3 | 4413.37 ± 151.21 a | 678.24 ± 107.11 a | 1079.89 ± 77.55 a | 141.24 ± 23.59 a | 186.47 ± 13.59 a | 2327.53 ± 139.19 b |
| 6 | 4315.56 ± 147.14 a | 845.24 ± 125.24 a,b | 1024.54 ± 86.78 a | 155.73 ± 16.10 a | 196.59 ± 16.10 a | 2093.46 ± 170.37 a,b |
| 9 | 4532.36 ± 284.82 a | 1048.12 ± 76.37 b | 1263.78 ± 97.73 a | 221.47 ± 31.68 b | 201.21 ± 21.68 a | 1797.78 ± 114.54 a |
| 12 | 4557.24 ± 147.97 a | 1175.11 ± 92.45 b | 1243.45 ± 107.73 a | 233.45 ± 21.68 b | 214.53 ± 21.68 a | 1690.70 ± 114.54 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, Z.; Zhang, Y.; Sun, L. Effects of Dietary Astaxanthin Supplementation on Energy Budget and Bioaccumulation in Procambarus clarkii (Girard, 1852) Crayfish under Microcystin-LR Stress. Toxins 2018, 10, 277. https://doi.org/10.3390/toxins10070277
An Z, Zhang Y, Sun L. Effects of Dietary Astaxanthin Supplementation on Energy Budget and Bioaccumulation in Procambarus clarkii (Girard, 1852) Crayfish under Microcystin-LR Stress. Toxins. 2018; 10(7):277. https://doi.org/10.3390/toxins10070277
Chicago/Turabian StyleAn, Zhenhua, Yingying Zhang, and Longshen Sun. 2018. "Effects of Dietary Astaxanthin Supplementation on Energy Budget and Bioaccumulation in Procambarus clarkii (Girard, 1852) Crayfish under Microcystin-LR Stress" Toxins 10, no. 7: 277. https://doi.org/10.3390/toxins10070277
APA StyleAn, Z., Zhang, Y., & Sun, L. (2018). Effects of Dietary Astaxanthin Supplementation on Energy Budget and Bioaccumulation in Procambarus clarkii (Girard, 1852) Crayfish under Microcystin-LR Stress. Toxins, 10(7), 277. https://doi.org/10.3390/toxins10070277