Distribution of Tetrodotoxin in the New Zealand Clam, Paphies australis, Established Using Immunohistochemistry and Liquid Chromatography-Tandem Quadrupole Mass Spectrometry

 , and
, and
Abstract
:1. Introduction
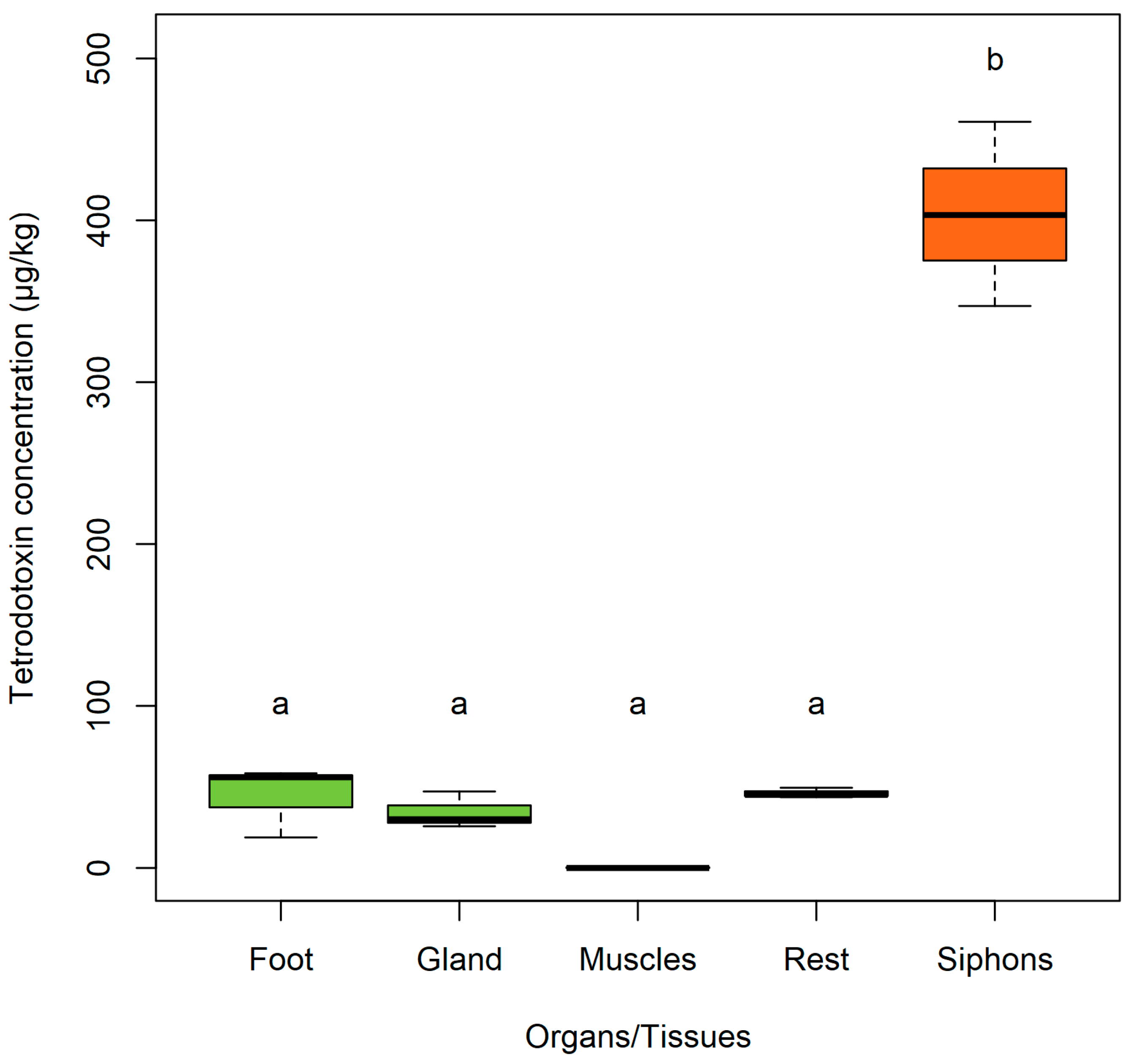
2. Results and Discussion
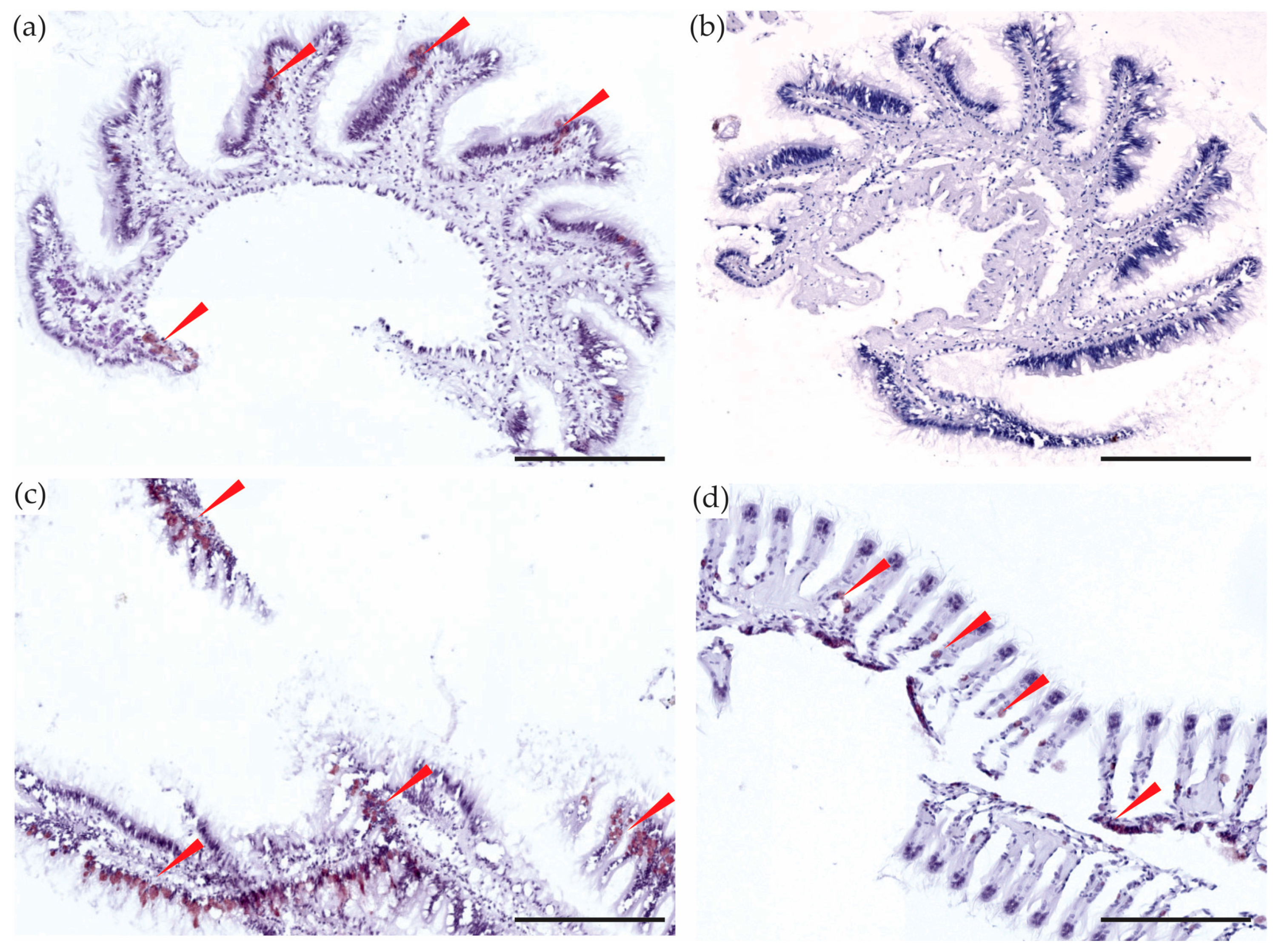
2.1. Siphons
2.2. Foot
2.3. Adductor Muscles
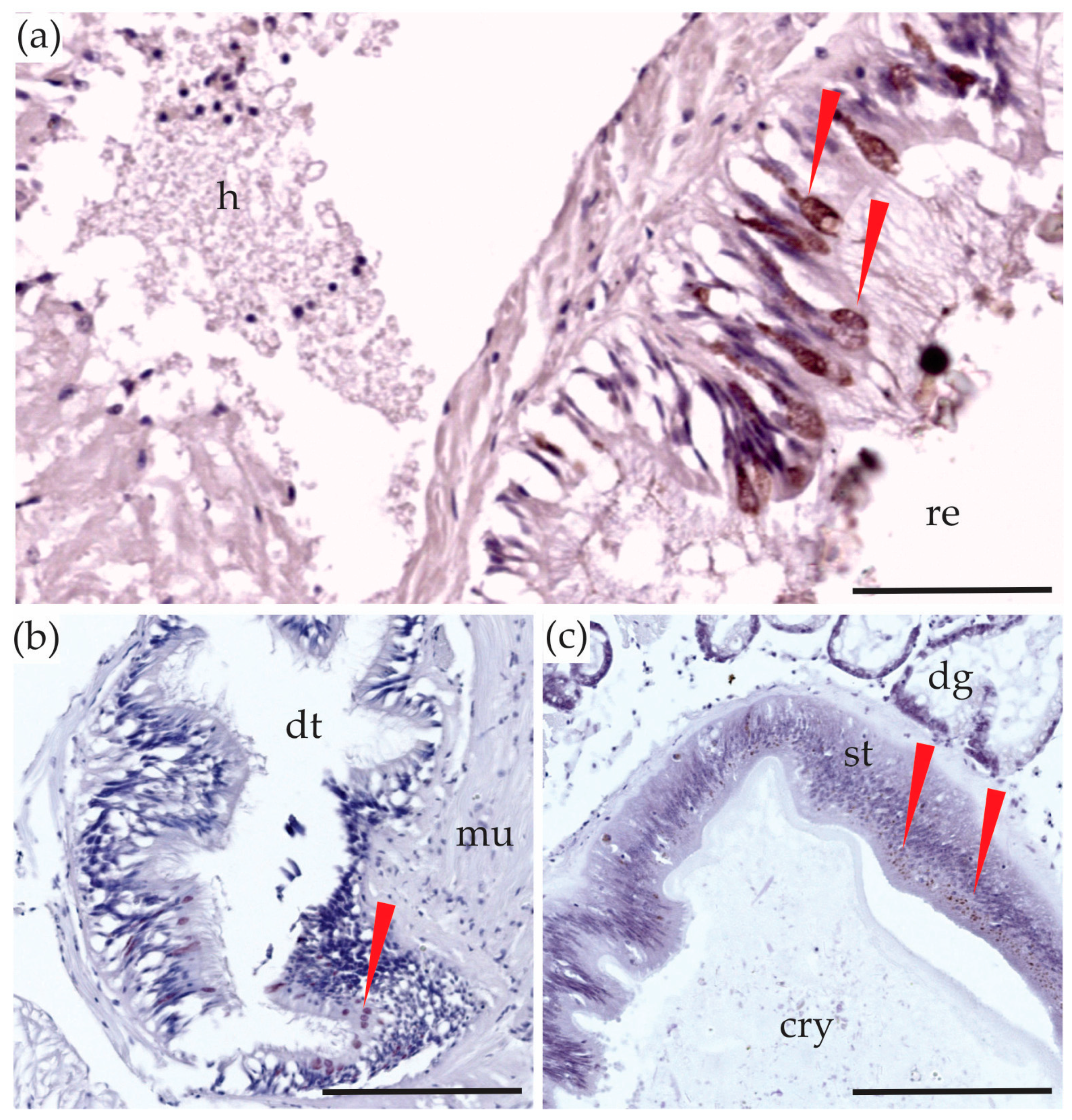
2.4. Digestive System
2.5. The ‘rest’
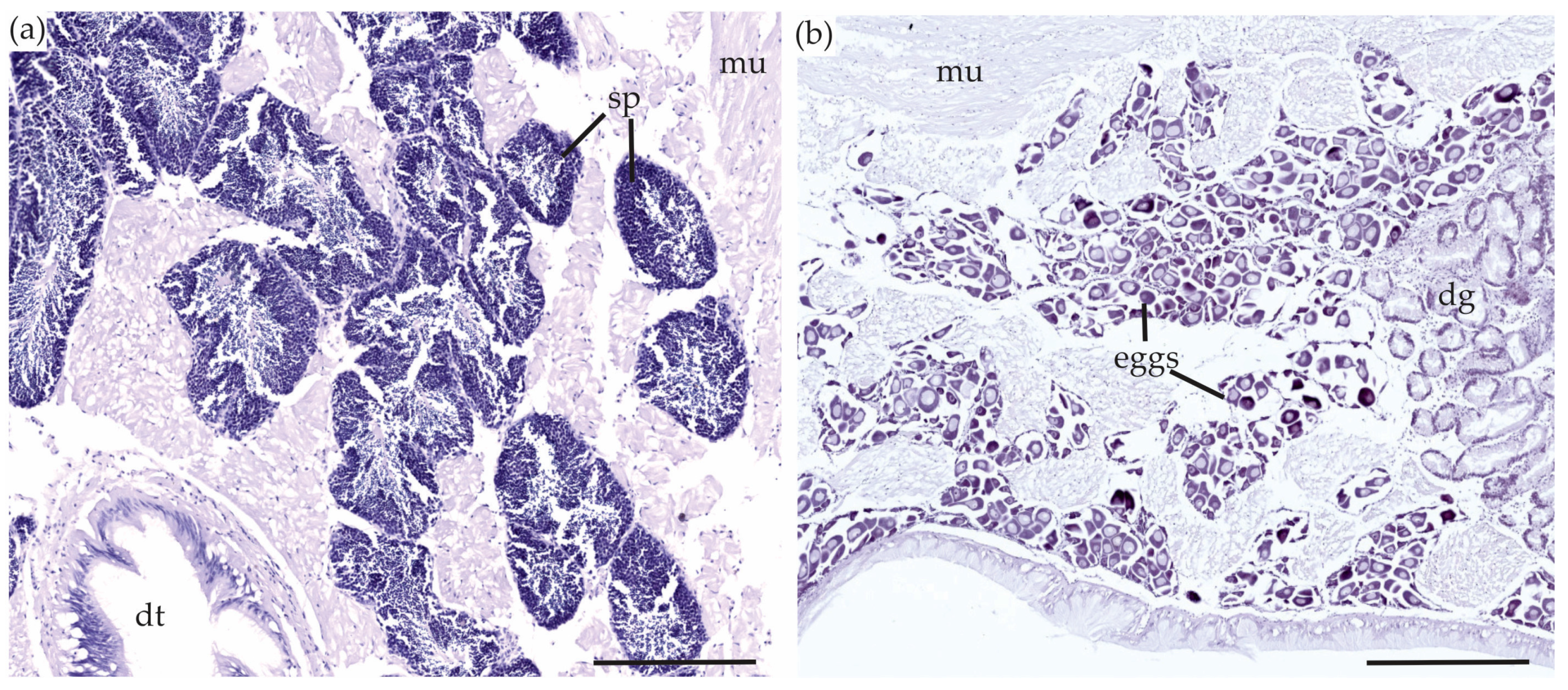
2.5.1. Gonads
2.5.2. Mantle
2.5.3. Gills and Labial Palps
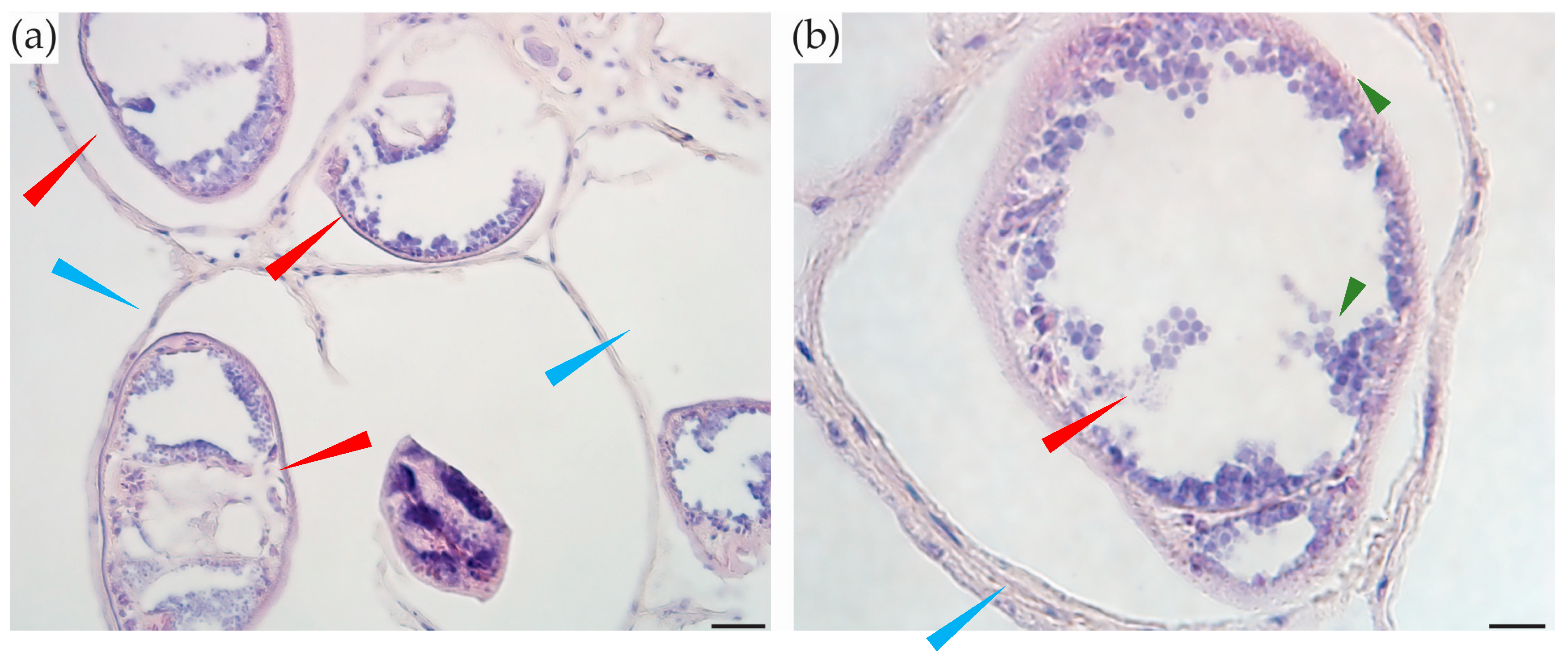
2.6. Parasites
3. Materials and Methods
3.1. Paphies australis Collection
3.2. Sample Processing
3.2.1. Sectioning for Immunohistochemistry
3.2.2. Dissection for Tetrodotoxin Analysis
3.2.3. Immunohistochemistry
3.2.4. Tetrodotoxin Analysis
3.2.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cestèle, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Noguchi, T.; Ebesu, J.S. Puffer poisoning: epidemiology and treatment. J. Toxicol. Toxin Rev. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Tahara, Y. Uber das tetrodongift. Biochemistry 1910, 30, 255–275. [Google Scholar]
- Goto, T.; Kishi, Y.; Takahashi, S.; Hirata, Y. Tetrodotoxin. Tetrahedron 1965, 21, 2059–2088. [Google Scholar] [CrossRef]
- Tsuda, K.; Ikuma, S.; Kawamura, M.; Tachikawa, R.; Sakai, K.; Tamura, C.; Amakasu, O. Tetrodotoxin. VII. On the structures of tetrodotoxin and its derivatives. Chem. Pharm. Bull. 1964, 12, 1357–1374. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, F.A. Tetrodotoxin. Sci. Am. 1967, 217, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Kodama, M.; Noguchi, T.; Maruyama, J.; Ogata, T.; Hashimoto, K. Release of tetrodotoxin and paralytic shellfish poison from puffer liver by RNase. J. Biochem. 1983, 93, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Isbister, G.K.; Son, J.; Wang, F.; Maclean, C.J.; Lin, C.S.; Ujma, J.; Balit, C.R.; Smith, B.; Milder, D.G.; Kiernan, M.C. Puffer fish poisoning: A potentially life-threatening condition. Med. J. Aust. 2002, 177, 650–653. [Google Scholar] [PubMed]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origins and biosynthesis of tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Bane, V.; Lehane, M.; Dikshit, M.; O‘Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O. Tetrodotoxin–distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Yasumura, D.; Yotsu, M.; Michishita, T.; Endo, A.; Kotaki, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric. Biol. Chem. 1986, 50, 793–795. [Google Scholar]
- Wu, Z.; Yang, Y.; Xie, L.; Xia, G.; Hu, J.; Wang, S.; Zhang, R. Toxicity and distribution of tetrodotoxin-producing bacteria in puffer fish Fugu rubripes collected from the Bohai Sea of China. Toxicon 2005, 46, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Mebs, D.; Yasumoto, T. Tetrodotoxin and its analogues in extracts from the toad Atelopus oxyrhynchus (family: Bufonidae). Toxicon 1992, 30, 1489–1492. [Google Scholar] [CrossRef]
- Daly, J.W.; Padgett, W.L.; Saunders, R.L.; Cover, J.F., Jr. Absence of tetrodotoxins in a captive-raised riparian frog, Atelopus varius. Toxicon 1997, 35, 705–709. [Google Scholar] [CrossRef]
- Cardall, B.L.; Brodie, E.D.; Hanifin, C.T. Secretion and regeneration of tetrodotoxin in the rough-skin newt (Taricha granulosa). Toxicon 2004, 44, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Kodama, M.; Sato, S.; Sakamoto, S.; Ogata, T. Occurrence of tetrodotoxin in Alexandrium tamarense, a causative dinoflagellate of paralytic shellfish poisoning. Toxicon 1996, 34, 1101–1105. [Google Scholar] [CrossRef]
- McNabb, P.S.; Taylor, D.I.; Ogilvie, S.C.; Wilkinson, L.; Anderson, A.; Hamon, D.; Wood, S.A.; Peake, B.M. First detection of tetrodotoxin in the bivalve Paphies australis by liquid chromatography coupled to triple quadrupole mass spectrometry with and without precolumn reaction. J. AOAC Int. 2014, 97, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.; Powell, A.; Schofield, A.; Lees, D.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Eur. Surveill. 2015, 20, 2–8. [Google Scholar] [CrossRef]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First detection of tetrodotoxin in Greek shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779–1807. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, H.K.; Alexander, J.; BarregAard, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B. Risks for public health related to the presence of tetrodotoxin (TTX) and TTX analogues in marine bivalves and gastropods. Eur. Food Saf. Auth. J. 2017, 15, 4752. [Google Scholar]
- Zhang, X.; Yan, Z.; Wang, Y.; Jiang, T.; Wang, J.; Sun, X.; Guo, Y. Immunoaffinity chromatography purification and ultrahigh performance liquid chromatography tandem mass spectrometry determination of tetrodotoxin in marine organisms. J. Agric. Food Chem. 2015, 63, 3129–3134. [Google Scholar] [CrossRef] [PubMed]
- Boundy, M.J.; Selwood, A.I.; Harwood, D.T.; McNabb, P.S.; Turner, A.D. Development of a sensitive and selective liquid chromatography–mass spectrometry method for high throughput analysis of paralytic shellfish toxins using graphitised carbon solid phase extraction. J. Chromatogr. A 2015, 1387, 1–12. [Google Scholar] [CrossRef] [PubMed]
- McNabb, P.; Selwood, A.I.; Holland, P.T. Multiresidue method for determination of algal toxins in shellfish: Single-laboratory validation and interlaboratory study. J. AOAC Int. 2005, 88, 761–772. [Google Scholar] [PubMed]
- Tanu, M.; Mahmud, Y.; Takatani, T.; Kawatsu, K.; Hamano, Y.; Arakawa, O.; Noguchi, T. Localization of tetrodotoxin in the skin of a brackishwater puffer Tetraodon steindachneri on the basis of immunohistological study. Toxicon 2002, 40, 103–106. [Google Scholar] [CrossRef]
- Mahmud, Y.; Okada, K.; Takatani, T.; Kawatsu, K.; Hamano, Y.; Arakawa, O.; Noguchi, T. Intra-tissue distribution of tetrodotoxin in two marine puffers Takifugu vermicularis and Chelonodon patoca. Toxicon 2003, 41, 13–18. [Google Scholar] [CrossRef]
- Salvitti, L.R.; Wood, S.A.; Winsor, L.; Cary, S.C. Intracellular immunohistochemical detection of tetrodotoxin in Pleurobranchaea maculata (Gastropoda) and Stylochoplana sp. (Turbellaria). Mar. Drugs 2015, 13, 756–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanu, M.B.; Mahmud, Y.; Arakawa, O.; Takatani, T.; Kajihara, H.; Kawatsu, K.; Hamano, Y.; Asakawa, M.; Miyazawa, K.; Noguchi, T. Immunoenzymatic visualization of tetrodotoxin (TTX) in Cephalothrix species (Nemertea: Anopla: Palaeonemertea: Cephalotrichidae) and Planocera reticulata (Platyhelminthes: Turbellaria: Polycladida: Planoceridae). Toxicon 2004, 44, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Magarlamov, T.Y.; Shokur, O.A.; Chernyshev, A.V. Distribution of tetrodotoxin in the ribbon worm Lineus alborostratus (Takakura, 1898)(nemertea): Immunoelectron and immunofluorescence studies. Toxicon 2016, 112, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Rivera, V.R.; Poli, M.A.; Bignami, G.S. Prophylaxis and treatment with a monoclonal antibody of tetrodotoxin poisoning in mice. Toxicon 1995, 33, 1231–1237. [Google Scholar] [CrossRef]
- Kawatsu, K.; Hamano, Y.; Yoda, T.; Terano, Y.; Shibata, T. Rapid and highly sensitive enzyme immunoassay for quantitative determination of tetrodotoxin. Jpn. J. Med. Sci. Biol. 1997, 50, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Smolowitz, R.; Doucette, G. Immunohistochemical localization of saxitoxin in the siphon epithelium of the butter clam, Saxidomus giganteus. Biol. Bull. 1995, 189, 229–230. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, L.; White, D.; Adamson, J. Temporal variation and tissue localization of paralytic shellfish toxins in the New Zealand Tuatua (surf-clam), Paphies subtriangulata. J. Shellfish Res. 1996, 15, 735–740. [Google Scholar]
- Kvitek, R.G. Paralytic shellfish toxins sequestered by bivalves as a defense against siphon-nipping fish. Mar. Biol. 1991, 111, 369–374. [Google Scholar] [CrossRef]
- Hine, P.M. The inter-relationships of bivalve haemocytes. Fish Shellfish Immunol. 1999, 9, 367–385. [Google Scholar] [CrossRef]
- Stanley, S.M. Post-Paleozoic adaptive radiation of infaunal bivalve molluscs: A consequence of mantle fusion and siphon formation. J. Paleontol. 1968, 42, 214–229. [Google Scholar]
- Gosling, E. Bivalve Molluscs: Biology, Ecology and Culture; Wiley-Blackwell: Oxford, UK, 2008; Volume 1, p. 448. [Google Scholar]
- Zwarts, L.; Wanink, J. Siphon size and burying depth in deposit-and suspension-feeding benthic bivalves. Mar. Biol. 1989, 100, 227–240. [Google Scholar] [CrossRef]
- De Goeij, P.; Luttikhuizen, P.C.; van der Meer, J.; Piersma, T. Facilitation on an intertidal mudflat: The effect of siphon nipping by flatfish on burying depth of the bivalve Macoma balthica. Oecologia 2001, 126, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Kamermans, P.; Huitema, H.J. Shrimp (Crangon crangon L.) browsing upon siphon tips inhibits feeding and growth in the bivalve Macoma balthica (L.). J. Exp. Mar. Biol. Ecol. 1994, 175, 59–75. [Google Scholar] [CrossRef]
- Kvitek, R.; Bretz, C. Harmful algal bloom toxins protect bivalve populations from sea otter predation. Mar. Ecol. Prog. Ser. 2004, 271, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Stace, G. Burrowing ability and life position of Toheroa (Paphies ventricosa: Mesodesmatidae), an unusually large, deep-burrowing ocean beach bivalve endemic to New Zealand. Jpn. J. Malacol. 1995, 54, 67–76. [Google Scholar]
- Bricelj, V.M.; Shumway, S.E. Paralytic shellfish toxins in bivalve molluscs: occurrence, transfer kinetics, and biotransformation. Rev. Fish. Sci. 1998, 6, 315–383. [Google Scholar] [CrossRef]
- Hwang, D.F.; Tsai, Y.H.; Cheng, C.A.; Jeng, S.S. Comparison of paralytic toxins in aquaculture of purple clam in Taiwan. Toxicon 1992, 30, 669–672. [Google Scholar] [CrossRef]
- Pereira, P.; Dias, E.; Franca, S.; Pereira, E.; Carolino, M.; Vasconcelos, V. Accumulation and depuration of cyanobacterial paralytic shellfish toxins by the freshwater mussel Anodonta cygnea. Aquat. Toxicol. 2004, 68, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, J.; Opie, A.M. On the role of octopine dehydrogenase in the adductor muscles of bivalve molluscs. Comp. Biochem. Physiol. Part B Biochem. Mol. Bol. 1978, 61, 85–92. [Google Scholar] [CrossRef]
- Shumway, S.E.; Cembella, A.D. The impact of toxic algae on scallop culture and fisheries. Rev. Fish. Sci. 1993, 1, 121–150. [Google Scholar] [CrossRef]
- Helm, M.M.; Bourne, N.; Lovatelli, A. Hatchery Culture of Bivalves: A Practical Manual, 2004. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/docrep/007/y5720e/y5720e00.htm (accessed on 11 June 2018).
- Harada, T.; Oshima, Y.; Kamiya, H.; Yasumoto, T. Confirmation of paralytic shellfish toxins in the dinoflagellate Pyrodinium bahamense var. compressa and bivalves in Palau. Nippon Suisan Gakkaishi 1982, 48, 821–825. [Google Scholar] [CrossRef]
- Mafra, L.L., Jr.; Bricelj, V.M.; Fennel, K. Domoic acid uptake and elimination kinetics in oysters and mussels in relation to body size and anatomical distribution of toxin. Aquat. Toxicol. 2010, 100, 17–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Xie, P. Seasonal dynamics of the hepatotoxic microcystins in various organs of four freshwater bivalves from the large eutrophic lake Taihu of subtropical China and the risk to human consumption. Environ. Toxicol. 2005, 20, 572–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoi, S.; Suzuki, M.; Asahina, K.; Sawayama, E.; Nishikubo, J.; Oyama, H.; Takei, M.; Shiibashi, N.; Takatani, T.; Arakawa, O. Role of maternal tetrodotoxin in survival of larval pufferfish. Toxicon 2018, 148, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Purchon, R.D. The Biology of The Mollusca; Pergamon Press: Oxford, UK, 1968; Volume 1, p. 560. [Google Scholar]
- Howard, D.W.; Lewis, E.J.; Keller, B.J.; Smith, C.S. Histological Techniques for Marine Bivalve Mollusks and Crustaceans, 2nd ed.; NOAA: Oxford, MD, USA, 2004; Volume 5, p. 218.
- Salvitti, L.; Wood, S.A.; Taylor, D.I.; McNabb, P.; Cary, S.C. First identification of tetrodotoxin (TTX) in the flatworm Stylochoplana sp.; a source of TTX for the sea slug Pleurobranchaea maculata. Toxicon 2015, 95, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Boundy, M.J.; Rapkova, M.D. Development and single-laboratory validation of a liquid chromatography tandem mass spectrometry method for quantitation of Tetrodotoxin in mussels and oysters. J. AOAC Int. 2017, 100, 1469–1482. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: www.r-project.org (accessed on 22 May 2018).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Step | Solution | Time (min) |
|---|---|---|
| 1. | 3% H2O2/10% methanol | 10 |
| 2. | 1 × PBS | 10 × 3 |
| 3. | Normal Goat Serum | 20 |
| 4. | 1 × PBS | 10 × 3 |
| 5. | mAB T20G10 * | Overnight at 4 °C |
| 6. | 1 × PBS | 10 × 3 |
| 7. | Biotinylated secondary antibody (anti-rabbit IgG) * | 60 |
| 8. | 1 × PBS | 10 × 3 |
| 9. | VECTASTAIN® ABC reagent * | 60 |
| 10. | 1 × PBS | 10 × 3 |
| 11. | DAB * | 5–10 |
| 12. | Deionized H2O | 5 |
| 13. | Counterstain (Gill’s II Hematoxylin) | 5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biessy, L.; Smith, K.F.; Boundy, M.J.; Webb, S.C.; Hawes, I.; Wood, S.A. Distribution of Tetrodotoxin in the New Zealand Clam, Paphies australis, Established Using Immunohistochemistry and Liquid Chromatography-Tandem Quadrupole Mass Spectrometry. Toxins 2018, 10, 282. https://doi.org/10.3390/toxins10070282
Biessy L, Smith KF, Boundy MJ, Webb SC, Hawes I, Wood SA. Distribution of Tetrodotoxin in the New Zealand Clam, Paphies australis, Established Using Immunohistochemistry and Liquid Chromatography-Tandem Quadrupole Mass Spectrometry. Toxins. 2018; 10(7):282. https://doi.org/10.3390/toxins10070282
Chicago/Turabian StyleBiessy, Laura, Kirsty F. Smith, Michael J. Boundy, Stephen C. Webb, Ian Hawes, and Susanna A. Wood. 2018. "Distribution of Tetrodotoxin in the New Zealand Clam, Paphies australis, Established Using Immunohistochemistry and Liquid Chromatography-Tandem Quadrupole Mass Spectrometry" Toxins 10, no. 7: 282. https://doi.org/10.3390/toxins10070282
APA StyleBiessy, L., Smith, K. F., Boundy, M. J., Webb, S. C., Hawes, I., & Wood, S. A. (2018). Distribution of Tetrodotoxin in the New Zealand Clam, Paphies australis, Established Using Immunohistochemistry and Liquid Chromatography-Tandem Quadrupole Mass Spectrometry. Toxins, 10(7), 282. https://doi.org/10.3390/toxins10070282