Imbalance in the Blood Concentrations of Selected Steroids in Pre-pubertal Gilts Depending on the Time of Exposure to Low Doses of Zearalenone


Abstract
:1. Introduction
2. Results
2.1. Experimental Feed
2.2. Clinical Observations
2.3. The Effect of Various Doses of Zearalenone on Hormone Secretion
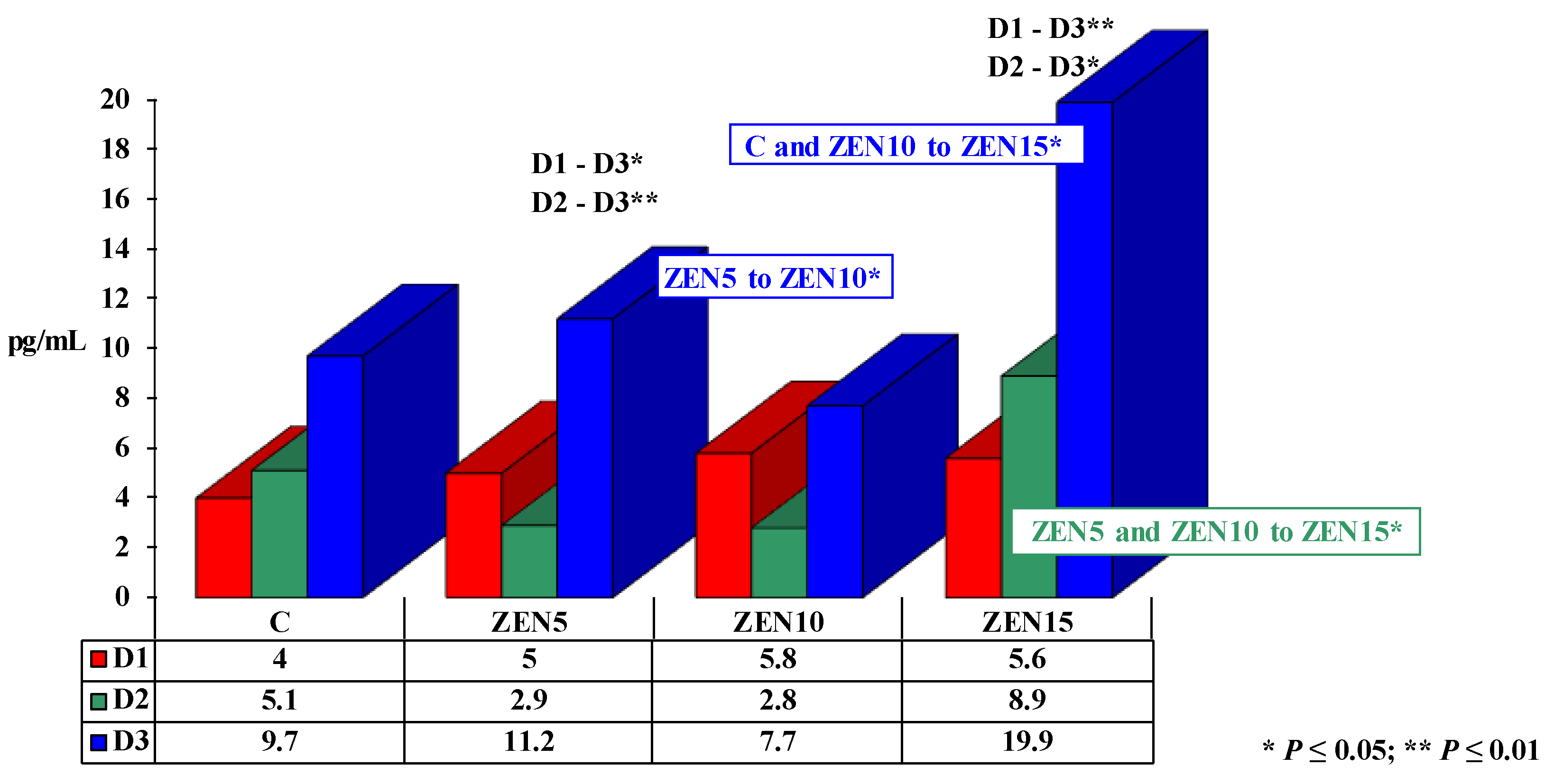
2.3.1. The Effect of Estradiol
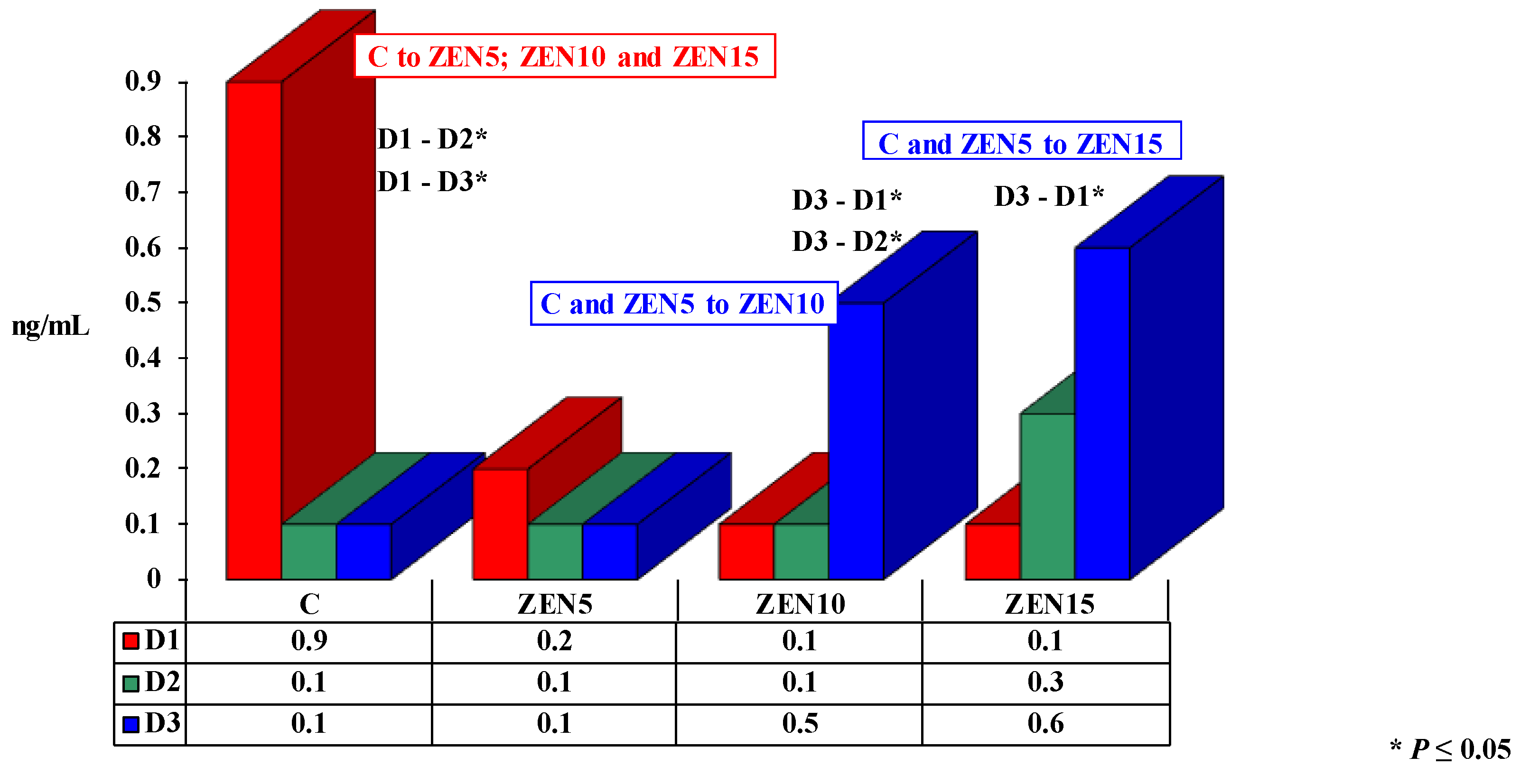
2.3.2. The Effect of Progesterone
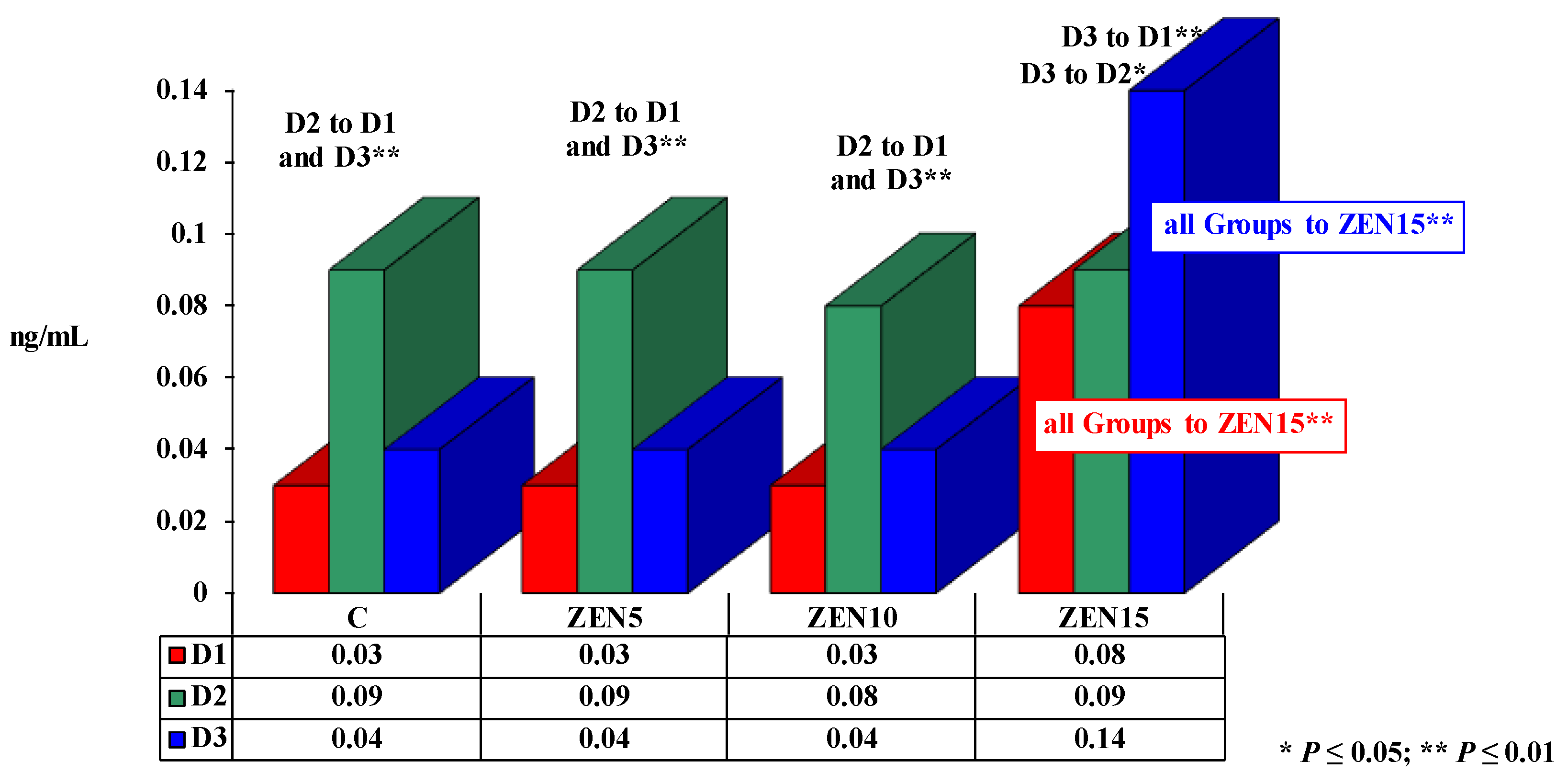
2.3.3. The Effect of Testosterone
2.3.4. Pearson’s Correlation Coefficient (r)
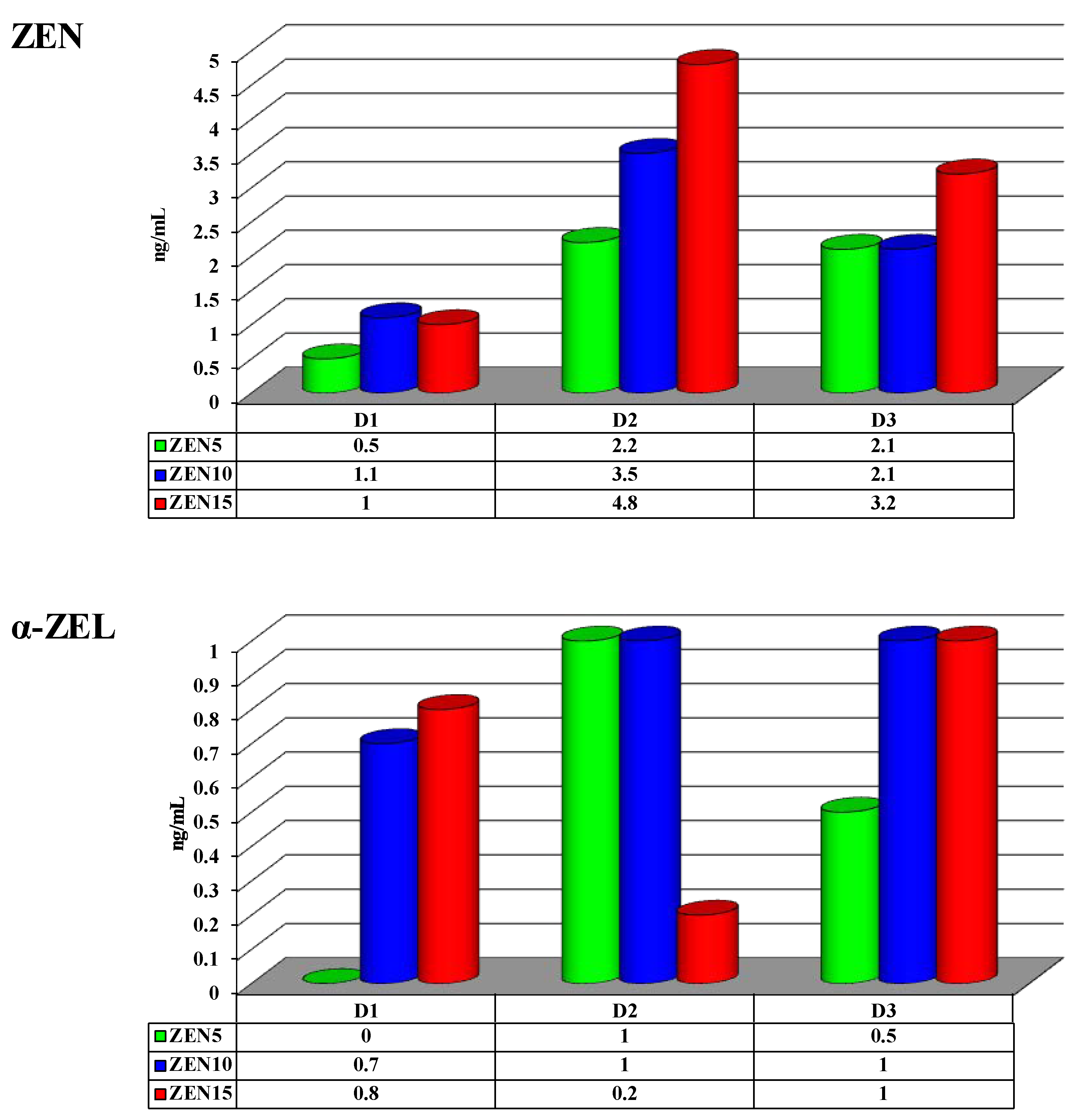
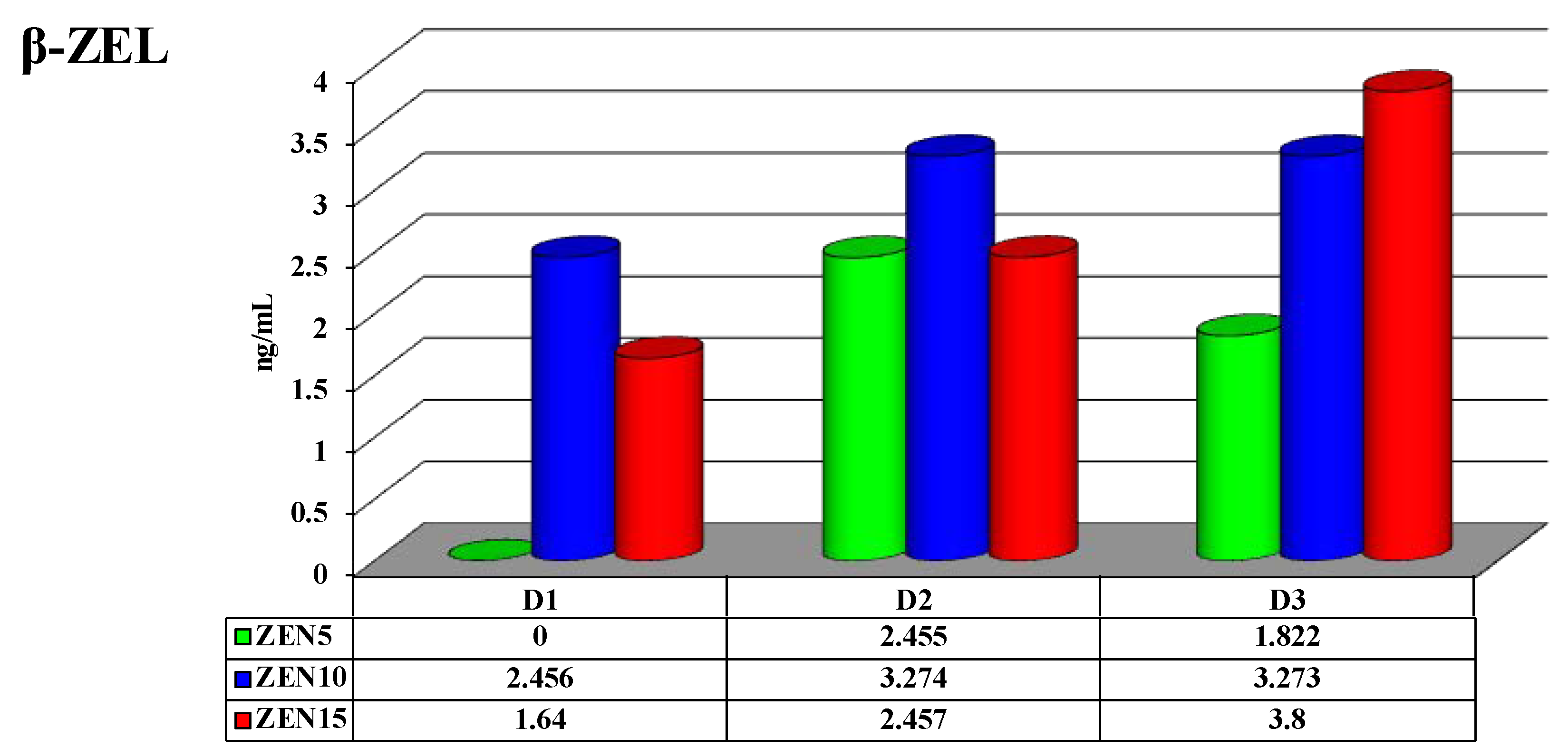
2.4. Concentrations of Zearalenone and its Metabolites in Peripheral Blood
3. Discussion
3.1. Estradiol
3.2. Progesterone
3.3. Testosterone
3.4. Zearalenone (ZEN) and its Metabolites
3.5. Summary
- -
- a minor increase was noted in peripheral blood levels of E2 and “free ZEN”, proportionally to the ZEN dose and analytical date;
- -
- the concentration of E2 in peripheral blood decreased on D1 in all experimental groups and it increased on D2 and D3 in selected experimental groups (in group ZEN15 on D2, and in groups ZEN10 and ZEN15 on D3);
- -
- testosterone levels increased significantly on all analytical dates in response to the LOAEL dose, and the concentrations of E2 and T were not correlated or were bound by weak linear correlations.
4. Materials and Methods
4.1. General Information
4.2. Experimental Animals and Feed
4.3. Determination of Mycotoxins in Feed
4.4. Blood Sampling
4.5. Determination of Hormone Concentrations
4.5.1. Estradiol
4.5.2. Progesterone and Testosterone
4.5.3. Statistical Analysis
4.6. Extraction Procedure
4.6.1. Quantification of ZEN and Its Metabolites
4.6.2. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Zheng, W.; Feng, N.; Wang, Y.; Noll, L.; Xu, S.; Liu, X.; Lu, N.; Zou, H.; Gu, J.; Yuan, Y.; et al. Effects of zearalenone and its derivatives on the synthesis and secretion of mammalian sex steroid hormones: A review. Food Chem. Toxicol. 2019, 126, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.M.; Bell, R.J.; Green, S.; Davis, S.R. Effects of testosterone therapy for women: A systematic review and meta-analysis protocol. Syst. Rev. 2019, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, H.-K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, 4851. [Google Scholar] [CrossRef]
- Pastoor, T.P.; Bachman, A.N.; Bell, D.R.; Cohen, S.M.; Dellarco, M.; Dewhurst, I.C.; Doe, J.E.; Doerrer, N.G.; Embry, M.R.; Hines, R.N.; et al. A 21st century roadmap for human health risk assessment. Crit. Rev. Toxicol. 2014, 44, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.-H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Cieplińska, K.; Gajęcka, M.; Dąbrowski, M.; Rykaczewska, A.; Zielonka, Ł.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Gajęcki, M.T. Time-dependent changes in the intestinal microbiome of gilts exposed to low zearalenone doses. Toxins 2019, 11, 296. [Google Scholar] [CrossRef]
- Cieplińska, K.; Gajęcka, M.; Nowak, A.; Dąbrowski, M.; Zielonka, Ł.; Gajęcki, M.T. The gentoxicity of caecal water in gilts exposed to low doses of zearalenone. Toxins 2018, 10, 350. [Google Scholar] [CrossRef]
- Rykaczewska, A.; Gajęcka, M.; Dąbrowski, M.; Wiśniewska, A.; Szcześniewska, J.; Gajęcki, M.T.; Zielonka, Ł. Growth performance, selected blood biochemical parameters and body weight of pre-pubertal gilts fed diets supplemented with different doses of zearalenone (ZEN). Toxicon 2018, 152, 84–94. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global Mycotoxin Occurrence in Feed: A Ten-Year Survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef]
- Pereira, C.S.; Cunha, S.C.; Fernandes, J.O. Prevalent Mycotoxins in Animal Feed: Occurrence and Analytical Methods. Toxins 2019, 11, 290. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Paradigm lost, paradigm found: The re-emergence of hormesis as a fundamental dose response model in the toxicological sciences. Environ. Pollut. 2005, 138, 378–411. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. The effect of low monotonic doses of zearalenone on selected reproductive tissues in pre-pubertal female dogs—A review. Molecules 2015, 20, 20669–20687. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. Activity of zearalenone in the porcine intestinal tract. Molecules 2017, 22, 18. [Google Scholar] [CrossRef] [PubMed]
- Luisetto, M.; Almukhtar, N.; Ahmadabadi, B.N.; Hamid, G.A.; Mashori, G.R.; Khan, K.R.; Khan, F.A.; Cabianca, L. Endogenus Toxicology: Modern Physio-Pathological Aspects and Relationship with New Therapeutic Strategies. An Integrative Discipline Incorporating Concepts from Different Research Discipline Like Biochemistry, Pharmacology and Toxicology. Clin. Pathol. 2019, 3, 000113. [Google Scholar] [CrossRef]
- He, J.; Wei, C.; Li, Y.; Liu, Y.; Wang, Y.; Pan, J.; Liu, J.; Wu, Y.; Cui, S. Zearalenone and alpha-zearalenol inhibit the synthesis and secretion of pig follicle stimulating hormone via the non-classical estrogen membrane receptor GPR30. Mol. Cell. Endocrinol. 2018, 461, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowski, M.; Obremski, K.; Gajęcka, M.; Gajęcki, M.T.; Zielonka, Ł. Changes in the Subpopulations of Porcine Peripheral Blood Lymphocytes Induced by Exposure to Low Doses of Zearalenone (ZEN) and Deoxynivalenol (DON). Molecules 2016, 21, 557. [Google Scholar] [CrossRef]
- Chen, L.; Giesy, J.P.; Xie, P. The dose makes the poison. Sci. Total Environ. 2018, 621, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.J.; Short, K.L.; Hooper, S.B. The science of steroids. Semin. Fetal Neonat. Med. 2019, 24, 170–175. [Google Scholar] [CrossRef]
- Przybylska-Gornowicz, B.; Lewczuk, B.; Prusik, M.; Hanuszewska, M.; Petrusewicz-Kosińska, M.; Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. The Effects of Deoxynivalenol and Zearalenone on the Pig Large Intestine. A Light and Electron Microscopy Study. Toxins 2018, 10, 148. [Google Scholar] [CrossRef]
- Przybylska-Gornowicz, B.; Tarasiuk, M.; Lewczuk, B.; Prusik, M.; Ziółkowska, N.; Zielonka, Ł.; Gajęcki, M.; Gajęcka, M. The Effects of Low Doses of Two Fusarium Toxins, Zearalenone and Deoxynivalenol, on the Pig Jejunum. A Light and Electron Microscopic Study. Toxins 2015, 7, 4684–4705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, M.S.; Ebel, E.D. Estimating correlation of prevalence at two locations in the farm-to-table continuum using qualitative test data. Int. J. Food Microbiol. 2017, 245, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Benagiano, M.; Bianchi, P.; D’Elios, M.M.; Brosens, I.; Benagiano, G. Autoimmune diseases: Role of steroid hormones. Best Pract. Res. Clin. Obstet. Gynaecol. 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Przybylska-Gornowicz, B. The low doses effect of experimental zearalenone (ZEN) intoxication on the presence of Ca2+ in selected ovarian cells from pre-pubertal bitches. Pol. J. Vet. Sci. 2012, 15, 711–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkenfeld, S.R.; Linc, C.; Frigo, D.E. Communication between genomic and non-genomic signaling events coordinate steroid hormone actions. Steroids 2018, 133, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Otrocka-Domagała, I. Immunocytochemical expression of 3β- and 17β-hydroxysteroid dehydrogenase in bitch ovaries exposed to low doses of zearalenone. Pol. J. Vet. Sci. 2013, 16, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Lawrenz, B.; Melado, L.; Fatemi, H. Premature progesterone rise in ART-cycles. Reprod. Biol. 2018, 18, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Schoevers, E.J.; Santos, R.R.; Colenbrander, B.; Fink-Gremmels, J.; Roelen, B.A.J. Transgenerational toxicity of Zearalenone in pigs. Reprod. Toxicol. 2012, 34, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Rivera, H.M.; Stincic, T.L. Estradiol and the control of feeding behavior. Steroids 2018, 133, 44–52. [Google Scholar] [CrossRef]
- Jiang, X.; Lu, N.; Xue, Y.; Liu, S.; Lei, H.; Tu, W.; Lu, Y.; Xia, D. Crude fiber modulates the fecal microbiome and steroid hormones in pregnant Meishan sows. Gen. Comp. Endocrinol. 2019, 277, 141–147. [Google Scholar] [CrossRef]
- Jakimiuk, E.; Kuciel-Lisieska, G.; Zwierzchowski, W.; Gajęcka, M.; Obremski, K.; Zielonka, Ł.; Skorska-Wyszyńska, E.; Gajęcki, M. Morphometric changes of the reproductive system in gilts during zearalenone mycotoxicosis. Med. Weter. 2006, 62, 99–102. [Google Scholar]
- Goldstein, I.; Kim, N.N.; Clayton, A.H.; DeRogatis, L.R.; Giraldi, A.; Parish, S.J.; Pfaus, J.; Simon, J.A.; Kingsberg, S.A.; Meston, C.; et al. Hypoactive Sexual Desire Disorder: International Society for the Study of Women’s Sexual Health (ISSWSH) Expert Consensus Panel Review. Mayo Clin. Proc. 2017, 92, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Blitek, A.; Szymanska, M.; Pieczywek, M.; Morawska-Pucinska, E. Luteal P4 synthesis in early pregnant gilts after induction of estrus with PMSG/hCG. Anim. Reprod. Sci. 2016, 166, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Kanakis, G.A.; Tsametis, C.P.; Goulis, DG. Measuring testosterone in women and men. Maturitas 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- White, S.F.; Lee, Y.; Phan, J.M.; Moody, S.N.; Shirtcliff, E.A. Putting the flight in “fight-or-flight”: Testosterone reactivity to skydiving is modulated by autonomic activation. Biol. Psychol. 2019, 143, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Bhasin, S.; Jasuja, R. Reproductive and Nonreproductive Actions of Testosterone. Enc. Endocr. Dis. 2019, 2, 721–734. [Google Scholar]
- Van Anders, S.M.; Steiger, J.; Goldey, K.L. Effects of gendered behavior on testosterone in women and men. Proc. Natl. Acad. Sci. USA 2015, 112, 13805–13810. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, V.; Scordo, M.; Yoon, R.S.; Liporace, F.A.; Greene, L.W. Revisiting the role of testosterone: Are we missing something? Rev. Urol. 2017, 19, 16–24. [Google Scholar] [CrossRef]
- Yang, D.; Jiang, T.; Lin, P.; Chen, H.; Wang, L.; Wang, N.; Zhao, F.; Tang, K.; Zhou, D.; Wang, A.; et al. Apoptosis inducing factor gene depletion inhibits zearalenone-induced cell death in a goat Leydig cell line. Reprod. Toxicol. 2017, 67, 129–139. [Google Scholar] [CrossRef]
- Gajęcka, M.; Rybarczyk, L.; Zwierzchowski, W.; Jakimiuk, E.; Zielonka, Ł.; Obremski, K.; Gajęcki, M. The effect of experimental, long-term exposure to low-dose zearalenone mycotoxicosis on the histological condition of ovaries in sexually immature gilts. Theriogenology 2011, 75, 1085–1094. [Google Scholar] [CrossRef]
- Hennig-Pauka, I.; Koch, F.J.; Schaumberger, S.; Woechtl, B.; Novak, J.; Sulyok, M.; Nagl, V. Current challenges in the diagnosis of zearalenone toxicosis as illustrated by a field case of hyperestrogenism in suckling piglets. Porc. Health Manag. 2018, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, Ł.; Waśkiewicz, A.; Beszterda, M.; Kostecki, M.; Dąbrowski, M.; Obremski, K.; Goliński, P.; Gajęcki, M. Zearalenone in the Intestinal Tissues of Immature Gilts Exposed per os to Mycotoxins. Toxins 2015, 7, 3210–3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demaegdt, H.; Daminet, B.; Evrard, A.; Scippo, M.L.; Muller, M.; Pussemier, L.; Callebaut, A.; Vandermeiren, K. Endocrine activity of mycotoxins and mycotoxin mixtures. Food Chem. Toxicol. 2016, 96, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Sławuta, P.; Nicpoń, J.; Kołacz, R.; Kiełbowicz, Z.; Zielonka, Ł.; Dąbrowski, M.; Szweda, W.; Gajęcki, M.; Nicpoń, J. Zearalenone and its metabolites in the tissues of female wild boars exposed per os to mycotoxins. Toxicon 2016, 114, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Parveen, M.; Zhu, Y.; Kiyama, R. Expression profiling of the genes responding to zearalenone and its analogues using estrogen-responsive genes. FEBS Lett. 2009, 583, 2377–2384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajęcka, M.; Zielonka, Ł.; Dąbrowski, M.; Mróz, M.; Gajęcki, M. The effect of low doses of zearalenone and its metabolites on progesterone and 17β-estradiol concentrations in peripheral blood and body weights of pre-pubertal female Beagle dogs. Toxicon 2013, 76, 260–269. [Google Scholar] [CrossRef]
- Heberer, T.; Lahrssen-Wiederholt, M.; Schafft, H.; Abraham, K.; Pzyrembel, H.; Henning, K.J.; Schauzu, M.; Braeunig, J.; Goetz, M.; Niemann, L.; et al. Zero tolerances in food and animal feed—Are there any scientific alternatives? A European point of view on an international controversy. Toxicol. Lett. 2007, 175, 118–135. [Google Scholar] [CrossRef]
- Smith, D.; Combes, R.; Depelchin, O.; Jacobsen, S.D.; Hack, R.; Luft, J.; Lammens, L.; von Landenberg, F.; Phillips, B.; Pfister, R.; et al. Optimising the design of preliminary toxicity studies for pharmaceutical safety testing in the dog. Regul. Toxicol. Pharmacol. 2005, 41, 95–101. [Google Scholar] [CrossRef]
- Gajęcki, M. The Effect of Experimentally Induced Fusarium Mycotoxicosis on Selected Diagnostic and Morphological Parameters of the Porcine Digestive Tract; Development Project NR12-0080-10 entitled; The National Centre for Research and Development: Warsaw, Poland, 30 November 2013; pp. 1–180.
- Kowalski, A.; Kaleczyc, J.; Gajęcki, M.; Zieliński, H. Adrenaline, noradrenaline and cortisol levels in pigs during blood collection (In Polish). Med. Weter. 1996, 52, 716–718. [Google Scholar]
- Gajęcka, M.; Woźny, M.; Brzuzan, P.; Zielonka, Ł.; Gajęcki, M. Expression of CYPscc and 3β-HSD mRNA in bitches ovary after long-term exposure to zearalenone. Bull. Vet. Inst. Pulawy 2011, 55, 777–780. [Google Scholar]
- Stanczyk, F.Z.; Xu, X.; Sluss, P.M.; Brinton, L.A.; McGlynn, K.A. Do metabolites account for higher serum steroid hormone levels measured by RIA compared to mass spectrometry? Clin. Chim. Acta 2018, 484, 223–225. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analytical Date | Group C | Group ZEN5 | Group ZEN10 | Group ZEN15 |
|---|---|---|---|---|
| D1 | 0.105 | 0.445 | −0.483 | 0.0 |
| D2 | 0.767 | 0.0 | 0.0 | 0.594 |
| D3 | 0.244 | −0.231 | −0.147 | 0.511 |
| Analytical Date | Group C | Group ZEN5 | Group ZEN10 | Group ZEN15 |
|---|---|---|---|---|
| D1 | 0.326 | 0.408 | −0.596 | −0.015 |
| D2 | 0.076 | 0.452 | 0.069 | 0.117 |
| D3 | −0.203 | −0.492 | −0.128 | −0.472 |
| Analytical Date | Group C | Group ZEN5 | Group ZEN10 | Group ZEN15 |
|---|---|---|---|---|
| D1 | 0.220 | 0.998 | 0.730 | 0.0 |
| D2 | 0.576 | 0.0 | 0.0 | 0.694 |
| D3 | 0.522 | 0.0 | 0.148 | −0.372 |
| Parameters | Composition Declared by The Manufacturer (%) |
|---|---|
| Soybean meal | 16 |
| Wheat | 55 |
| Barley | 22 |
| Wheat bran | 4.0 |
| Chalk | 0.3 |
| Zitrosan | 0.2 |
| Vitamin-mineral premix1 | 2.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rykaczewska, A.; Gajęcka, M.; Onyszek, E.; Cieplińska, K.; Dąbrowski, M.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Babuchowski, A.; Gajęcki, M.T.; Zielonka, Ł. Imbalance in the Blood Concentrations of Selected Steroids in Pre-pubertal Gilts Depending on the Time of Exposure to Low Doses of Zearalenone. Toxins 2019, 11, 561. https://doi.org/10.3390/toxins11100561
Rykaczewska A, Gajęcka M, Onyszek E, Cieplińska K, Dąbrowski M, Lisieska-Żołnierczyk S, Bulińska M, Babuchowski A, Gajęcki MT, Zielonka Ł. Imbalance in the Blood Concentrations of Selected Steroids in Pre-pubertal Gilts Depending on the Time of Exposure to Low Doses of Zearalenone. Toxins. 2019; 11(10):561. https://doi.org/10.3390/toxins11100561
Chicago/Turabian StyleRykaczewska, Anna, Magdalena Gajęcka, Ewa Onyszek, Katarzyna Cieplińska, Michał Dąbrowski, Sylwia Lisieska-Żołnierczyk, Maria Bulińska, Andrzej Babuchowski, Maciej T. Gajęcki, and Łukasz Zielonka. 2019. "Imbalance in the Blood Concentrations of Selected Steroids in Pre-pubertal Gilts Depending on the Time of Exposure to Low Doses of Zearalenone" Toxins 11, no. 10: 561. https://doi.org/10.3390/toxins11100561
APA StyleRykaczewska, A., Gajęcka, M., Onyszek, E., Cieplińska, K., Dąbrowski, M., Lisieska-Żołnierczyk, S., Bulińska, M., Babuchowski, A., Gajęcki, M. T., & Zielonka, Ł. (2019). Imbalance in the Blood Concentrations of Selected Steroids in Pre-pubertal Gilts Depending on the Time of Exposure to Low Doses of Zearalenone. Toxins, 11(10), 561. https://doi.org/10.3390/toxins11100561