Identification and Functional Characterization of a Novel Insecticidal Decapeptide from the Myrmicine Ant Manica rubida

 , ,
, ,
Abstract
:1. Introduction
2. Results
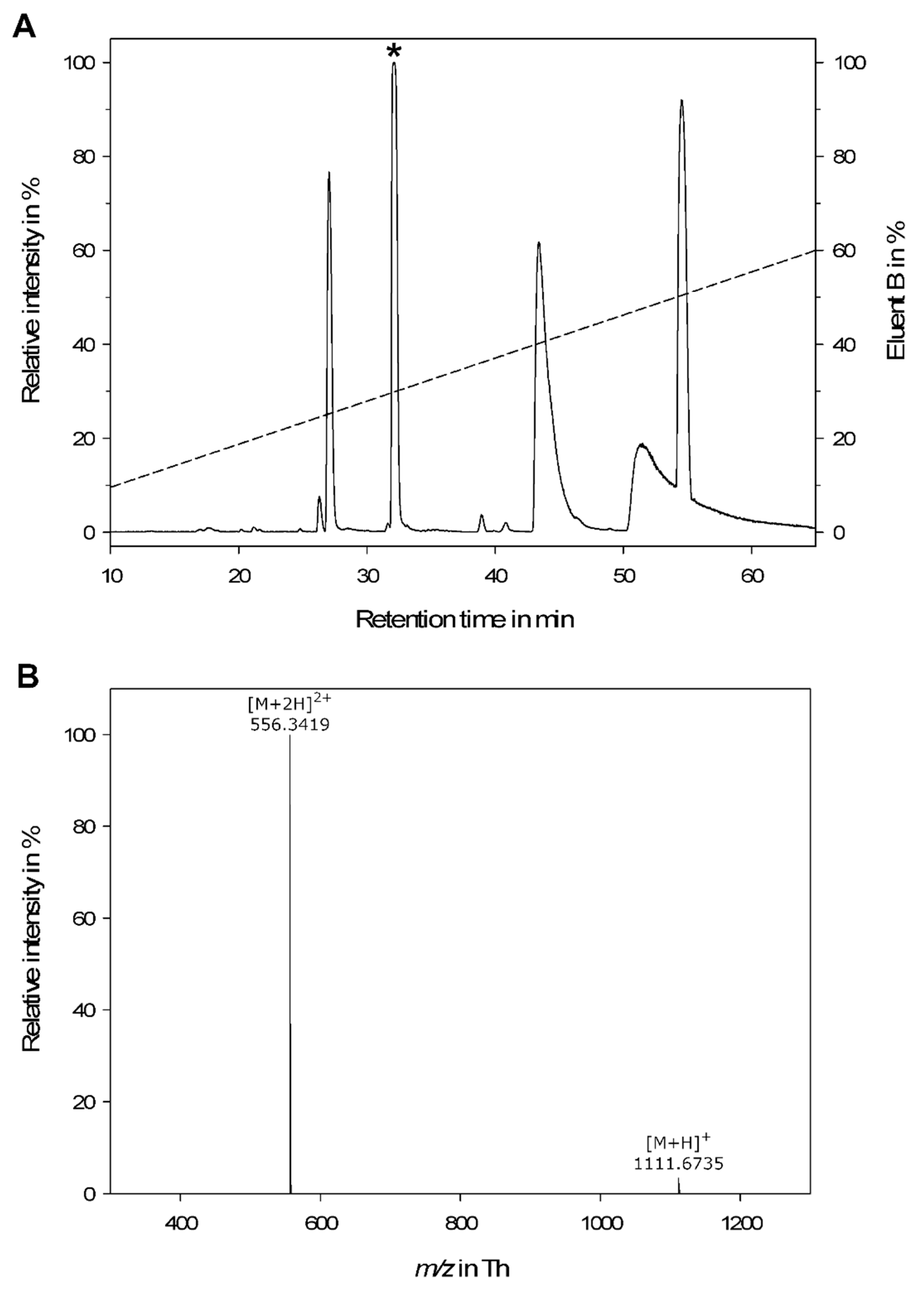
2.1. LC-MS Analysis of Crude Venom
2.2. Peptide Sequencing
2.3. Antimicrobial Activity
2.4. Effects of Crude Venom and Peptidyl Toxins on Aphid Survival and Reproduction
2.5. Effects of Crude Venom and Peptide Toxins on Aphid Behavior and Injection Wound Healing
2.6. Effects of Peptide U-MYRTX-MANr1 on Aphid Susceptibility to Chemical Insecticides
3. Discussion
4. Materials and Methods
4.1. Ant Collection and Taxonomy
4.2. Crude Venom Extraction
4.3. Reduction and Alkylation
4.4. LC-MS Analysis of Venom and Peptides
4.5. Edman Degradation
4.6. Peptide Synthesis
4.7. Determination of Total Protein Concentration for Injection Assays
4.8. Antimicrobial Assay
4.9. Maintenance of Aphids, Injection and Incapacitation Assays
4.10. Feeding Assays and Insecticide Bioassays
4.11. Histological Preparations
4.12. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- AntWeb. Available online: https://www.antweb.org (accessed on 1 July 2019).
- Touchard, A.; Aili, R.S.; Fox, G.E.; Escoubas, P.; Orivel, J.; Nicholson, M.G.; Dejean, A. The Biochemical Toxin Arsenal from Ant Venoms. Toxins 2016, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Aili, S.R.; Touchard, A.; Petitclerc, F.D.R.; Dejean, A.; Orivel, J.R.M.; Padula, M.P.; Escoubas, P.; Nicholson, G.M. Combined peptidomic and proteomic analysis of electrically stimulated and manually dissected venom from the South American bullet ant Paraponera clavata. J. Proteome Res. 2017, 16, 1339–1351. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Mueller, A.; Clayton, D.; Starobova, H.; Hamilton, B.R.; Payne, R.J.; Vetter, I.; King, G.F.; Undheim, E.A.B. A comprehensive portrait of the venom of the giant red bull ant, Myrmecia gulosa, reveals a hyperdiverse hymenopteran toxin gene family. Sci. Adv. 2018, 4, eaau4640. [Google Scholar] [CrossRef] [PubMed]
- Osman, M.F.H.; Brander, J. Weitere Beiträge zur Kenntnis der chemischen Zusammensetzung des Giftes von Ameisen aus der Gattung Formica. Z. Naturforsch. B 1961, 16, 749–753. [Google Scholar] [CrossRef]
- Pluzhnikov, K.A.; Kozlov, S.A.; Vassilevski, A.A.; Vorontsova, O.V.; Feofanov, A.V.; Grishin, E.V. Linear antimicrobial peptides from Ectatomma quadridens ant venom. Biochimie 2014, 107, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Rifflet, A.; Gavalda, S.; Tene, N.; Orivel, J.; Leprince, J.; Guilhaudis, L.; Genin, E.; Vetillard, A.; Treilhou, M. Identification and characterization of a novel antimicrobial peptide from the venom of the ant Tetramorium bicarinatum. Peptides 2012, 38, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Aili, S.R.; Touchard, A.; Escoubas, P.; Padula, M.P.; Orivel, J.; Dejean, A.; Nicholson, G.M. Diversity of peptide toxins from stinging ant venoms. Toxicon 2014, 92, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Kazuma, K.; Masuko, K.; Konno, K.; Inagaki, H. Combined Venom Gland Transcriptomic and Venom Peptidomic Analysis of the Predatory Ant Odontomachus monticola. Toxins 2017, 9, 323. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, W.; Klopp, C.; Verdenaud, M.; Ducancel, F.; Vétillard, A. Profiling the venom gland transcriptome of Tetramorium bicarinatum (Hymenoptera: Formicidae): The first transcriptome analysis of an ant species. Toxicon 2013, 70, 70–81. [Google Scholar] [CrossRef]
- Johnson, S.R.; Copello, J.A.; Evans, M.S.; Suarez, A.V. A biochemical characterization of the major peptides from the venom of the giant neotropical hunting ant Dinoponera australis. Toxicon 2010, 55, 702–710. [Google Scholar] [CrossRef]
- Orivel, J.; Redeker, V.; Le Caer, J.P.; Krier, F.; Revol-Junelles, A.M.; Longeon, A.; Chaffotte, A.; Dejean, A.; Rossier, J. Ponericins, new antibacterial and insecticidal peptides from the venom of the ant Pachycondyla goeldii. J. Biol. Chem. 2001, 276, 17823–17829. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.A.; Shattuck, S.O. The higher classification of the ant subfamily Ponerinae (Hymenoptera: Formicidae), with a review of ponerine ecology and behavior. Zootaxa 2014, 3817, 1–242. [Google Scholar] [CrossRef] [PubMed]
- Lima, D.B.; Torres, A.F.C.; Mello, C.P.; de Menezes, R.; Sampaio, T.L.; Canuto, J.A.; da Silva, J.J.A.; Freire, V.N.; Quinet, Y.P.; Havt, A.; et al. Antimicrobial effect of Dinoponera quadriceps (Hymenoptera: Formicidae) venom against Staphylococcus aureus strains. J. Appl. Microbiol. 2014, 117, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Schluns, H.; Crozier, R.H. Molecular and chemical immune defenses in ants (Hymenoptera: Formicidae). Myrmecol. News 2009, 12, 237–249. [Google Scholar]
- Pimentel, D. Pesticides and Pest Control. In Integrated Pest Management: Innovation-Development Process: Volume 1; Peshin, R., Dhawan, A.K., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 83–87. [Google Scholar]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2005, 144, 31–43. [Google Scholar] [CrossRef]
- Sparks, T.C.; Nauen, R. IRAC: Mode of action classification and insecticide resistance management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [Green Version]
- King, G.F.; Hardy, M.C. Spider-venom peptides: Structure, pharmacology, and potential for control of insect pests. Annu. Rev. Entomol. 2013, 58, 475–496. [Google Scholar] [CrossRef] [PubMed]
- Herzig, V.; Ikonomopoulou, M.; Smith, J.J.; Dziemborowicz, S.; Gilchrist, J.; Kuhn-Nentwig, L.; Rezende, F.O.; Moreira, L.A.; Nicholson, G.M.; Bosmans, F.; et al. Molecular basis of the remarkable species selectivity of an insecticidal sodium channel toxin from the African spider Augacephalus ezendami. Sci. Rep. 2016, 6, 29538. [Google Scholar] [CrossRef]
- Jørgensen, P.S.; Aktipis, A.; Brown, Z.; Carrière, Y.; Downes, S.; Dunn, R.R.; Epstein, G.; Frisvold, G.B.; Hawthorne, D.; Gröhn, Y.T.; et al. Antibiotic and pesticide susceptibility and the Anthropocene operating space. Nat. Sustain. 2018, 1, 632–641. [Google Scholar] [CrossRef]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- European Parliament. Think Thank. Farming without Plant Protection Products. Available online: https://www.europarl.europa.eu/RegData/etudes/IDAN/2019/634416/EPRS_IDA(2019)634416_EN.pdf (accessed on 1 July 2019).
- Windley, M.J.; Herzig, V.; Dziemborowicz, S.A.; Hardy, M.C.; King, G.F.; Nicholson, G.M. Spider-Venom Peptides as Bioinsecticides. Toxins 2012, 4, 191–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, G.F. Tying pest insects in knots: The deployment of spider-venom-derived knottins as bioinsecticides. Pest Manag. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Piek, T.; Duval, A.; Hue, B.; Karst, H.; Lapied, B.; Mantel, P.; Nakajima, T.; Pelhate, M.; Schmidt, J.O. Poneratoxin, a novel peptide neurotoxin from the venom of the ant, Paraponera clavata. Comp. Biochem. Physiol. C Comp. Pharmacol. 1991, 99, 487–495. [Google Scholar] [CrossRef]
- Piek, T.; Hue, B.; Mantel, P.; Terumi, N.; Schmidt, J.O. Pharmacological characterization and chemical fractionation of the venom of the ponerine ant, Paraponera clavata (F.). Comp. Biochem. Physiol. C Comp. Pharmacol. 1991, 99, 481–486. [Google Scholar] [CrossRef]
- Touchard, A.; Brust, A.; Cardoso, F.C.; Chin, Y.K.Y.; Herzig, V.; Jin, A.H.; Dejean, A.; Alewood, P.F.; King, G.F.; Orivel, J.; et al. Isolation and characterization of a structurally unique β-hairpin venom peptide from the predatory ant Anochetus emarginatus. Biochim. Biophys. Acta 2016, 1860, 2553–2562. [Google Scholar] [CrossRef] [PubMed]
- Van Emden, H.F.; Harrington, R. Aphids as Crop Pests; CABI: Wallingford, UK, 2017. [Google Scholar]
- Will, T.; Vilcinskas, A. Aphid-proof plants: Biotechnology-based approaches for aphid control. In Yellow Biotechnology II; Springer: Berlin/Heidelberg, Germany, 2013; pp. 179–203. [Google Scholar]
- Vilcinskas, A. Biology and Ecology of Aphids; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- IRAC—Insecticide Resistance Action Committee. Available online: http://www.irac-online.org/documents/sucking-pests-moa-poster/ (accessed on 1 July 2019).
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Luna-Ramirez, K.; Skaljac, M.; Grotmann, J.; Kirfel, P.; Vilcinskas, A. Orally delivered scorpion antimicrobial peptides exhibit activity against Pea Aphid (Acyrthosiphon pisum) and its bacterial symbionts. Toxins 2017, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Pal, N.; Yamamoto, T.; King, G.F.; Waine, C.; Bonning, B. Aphicidal efficacy of scorpion-and spider-derived neurotoxins. Toxicon 2013, 70, 114–122. [Google Scholar] [CrossRef]
- Xie, M.; Zhang, Y.J.; Zhai, X.M.; Zhao, J.J.; Peng, D.L.; Wu, G. Expression of a scorpion toxin gene BmKit enhances the virulence of Lecanicillium lecanii against aphids. J. Pest Sci. 2015, 88, 637–644. [Google Scholar] [CrossRef]
- Yang, S.; Fitches, E.; Pyati, P.; Gatehouse, J.A. Effect of insecticidal fusion proteins containing spider toxins targeting sodium and calcium ion channels on pyrethroid-resistant strains of peach-potato aphid (Myzus persicae). Pest Manag. Sci. 2015, 71, 951–956. [Google Scholar] [CrossRef]
- Bonning, B.C.; Pal, N.; Liu, S.; Wang, Z.; Sivakumar, S.; Dixon, P.M.; King, G.F.; Miller, W.A. Toxin delivery by the coat protein of an aphid-vectored plant virus provides plant resistance to aphids. Nat. Biotechnol. 2014, 32, 102. [Google Scholar] [CrossRef] [PubMed]
- Heep, J.; Klaus, A.; Kessel, T.; Seip, M.; Vilcinskas, A.; Skaljac, M. Proteomic Analysis of the Venom from the Ruby Ant Myrmica rubra and the Isolation of a Novel Insecticidal Decapeptide. Insects 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, G.C.; Wheeler, J. The Natural History of Manica (Hymenoptera: Formicidae). J. Kans. Entomol. Soc. 1970, 43, 129–162. [Google Scholar]
- Lenoir, A.; Devers, S.; Marchand, P.; Bressac, C.; Savolainen, R. Microgynous queens in the Paleartic ant, Manica rubida: Dispersal morphs or social parasites? J. Insect Sci. 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.J.F.; Bonhoeffer, S. On the evolution of claustral colony founding in ants. Evol. Ecol. Res. 2003, 5, 305–313. [Google Scholar]
- Touchard, A.; Koh, J.M.; Aili, S.R.; Dejean, A.; Nicholson, G.M.; Orivel, J.; Escoubas, P. The complexity and structural diversity of ant venom peptidomes is revealed by mass spectrometry profiling. Rapid Commun. Mass Spectrom. 2015, 29, 385–396. [Google Scholar] [CrossRef]
- Senko, M.W.; Beu, S.C.; McLaffertycor, F.W. Determination of monoisotopic masses and ion populations for large biomolecules from resolved isotopic distributions. J. Am. Soc. Mass Spectrom. 1995, 6, 229–233. [Google Scholar] [CrossRef] [Green Version]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Skaljac, M.; Kirfel, P.; Grotmann, J.; Vilcinskas, A. Fitness costs of infection with Serratia symbiotica are associated with greater susceptibility to insecticides in the pea aphid Acyrthosiphon pisum. Pest Manag. Sci. 2018, 74, 1829–1836. [Google Scholar] [CrossRef]
- Fiedler, K.; Kuhlmann, F.; Schlick-Steiner, B.C.; Steiner, F.M.; Gebauer, G. Stable N-isotope signatures of central European ants—Assessing positions in a trophic gradient. Insectes Soc. 2007, 54, 393–402. [Google Scholar] [CrossRef]
- Tani, N.; Kazuma, K.; Ohtsuka, Y.; Shigeri, Y.; Masuko, K.; Konno, K.; Inagaki, H. Mass Spectrometry Analysis and Biological Characterization of the Predatory Ant Odontomachus monticola Venom and Venom Sac Components. Toxins 2019, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Hink, W.F. Isolation and characterization of myrmexins, six isoforms of venom proteins with anti-inflammatory activity from the tropical ant, Pseudomyrmex triplarinus. Toxicon 2000, 38, 1403–1413. [Google Scholar] [CrossRef]
- Szolajska, E.; Poznanski, J.; Ferber, M.L.; Michalik, J.; Gout, E.; Fender, P.; Bailly, I.; Dublet, B.; Chroboczek, J. Poneratoxin, a neurotoxin from ant venom: Structure and expression in insect cells and construction of a bio-insecticide. Eur. J. Biochem. 2004, 271, 2127–2136. [Google Scholar] [CrossRef] [PubMed]
- Wanandy, T.; Gueven, N.; Davies, N.W.; Brown, S.G.A.; Wiese, M.D. Pilosulins: A review of the structure and mode of action of venom peptides from an Australian ant Myrmecia pilosula. Toxicon 2015, 98, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Rivier, J.; Clark, C.; Ramilo, C.A.; Corpuz, G.P.; Abogadie, F.C.; Mena, E.E.; Hillyard, D.R.; Cruz, L.J. Diversity of Conus neuropeptides. Science 1990, 249, 257–263. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of Peptide Hydrophobicity in the Mechanism of Action of α-Helical Antimicrobial Peptides. Antimicrob. Agents Chemother. 2007, 51, 1398. [Google Scholar] [CrossRef]
- Hale, J.D.F.; Hancock, R.E.W. Alternative mechanisms of action of cationic antimicrobial peptides on bacteria. Expert Rev. Anti-Infect. Ther. 2007, 5, 951–959. [Google Scholar] [CrossRef]
- Pirtskhalava, M.; Gabrielian, A.; Cruz, P.; Griggs, H.L.; Squires, R.B.; Hurt, D.E.; Grigolava, M.; Chubinidze, M.; Gogoladze, G.; Vishnepolsky, B.; et al. DBAASP v.2: An enhanced database of structure and antimicrobial/cytotoxic activity of natural and synthetic peptides. Nucleic Acids Res. 2015, 44, D1104–D1112. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Canofeni, S.; Miele, R.; Mangoni, M.L.; Barra, D. Temporins, Antimicrobial Peptides from the European Red Frog Rana temporaria. Eur. J. Biochem. 1996, 242, 788–792. [Google Scholar] [CrossRef]
- Luna-Ramirez, K.; Tonk, M.; Rahnamaeian, M.; Vilcinskas, A. Bioactivity of Natural and Engineered Antimicrobial Peptides from Venom of the Scorpions Urodacus yaschenkoi and U. manicatus. Toxins 2017, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.A.; Separovic, F. How membrane-active peptides get into lipid membranes. Acc. Chem. Res. 2016, 49, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Dushay, M.S. Insect hemolymph clotting. Cell. Mol. Life Sci. 2009, 66, 2643–2650. [Google Scholar] [CrossRef] [PubMed]
- Altincicek, B.; Gross, J.; Vilcinskas, A. Wounding-mediated gene expression and accelerated viviparous reproduction of the pea aphid Acyrthosiphon pisum. Insect Mol. Biol. 2008, 17, 711–716. [Google Scholar] [CrossRef]
- Er, A.; Sak, O.; Ergin, E.; Uçkan, F.; Rivers, D.B. Venom-induced immunosuppression: An overview of hemocyte-mediated responses. Psyche A J. Entomol. 2011, 2011, 276376. [Google Scholar] [CrossRef]
- Herzig, V.; De Araujo, D.A.; Greenwood, P.K.; Chin, K.Y.; Windley, J.M.; Chong, Y.; Muttenthaler, M.; Mobli, M.; Audsley, N.; Nicholson, M.G.; et al. Evaluation of Chemical Strategies for Improving the Stability and Oral Toxicity of Insecticidal Peptides. Biomedicines 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Keymanesh, K.; Soltani, S.; Sardari, S. Application of antimicrobial peptides in agriculture and food industry. World J. Microbiol. Biotechnol. 2009, 25, 933–944. [Google Scholar] [CrossRef]
- Marcos, J.F.; Muñoz, A.; Pérez-Payá, E.; Misra, S.; López-García, B. Identification and Rational Design of Novel Antimicrobial Peptides for Plant Protection. Annu. Rev. Phytopathol. 2008, 46, 273–301. [Google Scholar] [CrossRef]
- Seifert, B. Die Ameisen Mittel-und Nordeuropas; Lutra Verlag-u. Vertriebsges: Görlitz/Tauer, Germany, 2007. [Google Scholar]
- Anthis, N.J.; Clore, G.M. Sequence-specific determination of protein and peptide concentrations by absorbance at 205 nm. Protein Sci. 2013, 22, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Scopes, R.K. Measurement of protein by spectrophotometry at 205 nm. Anal. Biochem. 1974, 59, 277–282. [Google Scholar] [CrossRef]
- Hirsch, R.; Wiesner, J.; Marker, A.; Pfeifer, Y.; Bauer, A.; Hammann, P.E.; Vilcinskas, A. Profiling antimicrobial peptides from the medical maggot Lucilia sericata as potential antibiotics for MDR Gram-negative bacteria. J. Antimicrob. Chemother. 2018, 74, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Will, T.; Schmidtberg, H.; Skaljac, M.; Vilcinskas, A. Heat shock protein 83 plays pleiotropic roles in embryogenesis, longevity, and fecundity of the pea aphid Acyrthosiphon pisum. Dev. Genes Evol. 2017, 227, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sapountzis, P.; Duport, G.; Balmand, S.; Gaget, K.; Jaubert-Possamai, S.; Febvay, G.; Charles, H.; Rahbé, Y.; Colella, S.; Calevro, F. New insight into the RNA interference response against cathepsin-L gene in the pea aphid, Acyrthosiphon pisum: Molting or gut phenotypes specifically induced by injection or feeding treatments. Insect Biochem. Mol. Biol. 2014, 51, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Will, T.; Vilcinskas, A. The structural sheath protein of aphids is required for phloem feeding. Insect Biochem. Mol. Biol. 2015, 57, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, A.; Van Damme, E.J.M.; Smagghe, G.; Cullen, E. Evaluation of the susceptibility of the pea aphid, Acyrthosiphon pisum, to a selection of novel biorational insecticides using an artificial diet. J. Insect Sci. 2009, 9. [Google Scholar] [CrossRef] [PubMed]
- Akey, D.H.; Beck, S.D. Continuous Rearing of the Pea Aphid, Acyrthosiphon pisum, on a Holidic Diet. Ann. Entomol. Soc. Am. 1971, 64, 353–356. [Google Scholar] [CrossRef]
- IRAC—Insecticide Resistance Action Committee. Susceptibility Test Method 019. Available online: https://www.irac-online.org/methods/aphids-adultnymphs/ (accessed on 1 July 2019).
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Immunomedicine Group. Sequence Identity and Similarity (SIAS). Available online: http://bio.med.ucm.es/Tools/sias.html (accessed on 1 July 2019).




{kind=link}
{kind=link}
{kind=link}
| No. | RT [min] | Int. [%] | MWcrude [Da] | MWred. [Da] | MWalk. [Da] | S-S | Length a |
|---|---|---|---|---|---|---|---|
| 1 | 26.3 | 8 | 1314.71 | 1314.71 | 1314.72 | 0 | 12 |
| 2 | 26.3 | 7 | 1352.66 | 1352.66 | 1352.66 | 0 | 12 |
| 3 | 26.3 | 4 | 1336.69 | 1336.70 | 1336.70 | 0 | 12 |
| 4 | 27.0 | 77 | 1434.80 | 1436.83 | 1550.87 | 1 | 13 |
| 5 | 27.1 | 8 | 1208.64 | 1210.66 | 1324.70 | 1 | 11 |
| 6 | 27.1 | 3 | 1472.75 | 1474.77 | 1588.81 | 1 | 13 |
| 7 | 32.1 | 15 | 1132.64 | 1132.65 | 1132.65 | 0 | 10 |
| 8 | 32.1 | 28 | 1093.64 | 1093.64 | 1093.64 | 0 | 10 |
| 9 | 32.1 | 17 | 1148.61 | 1148.61 | 1148.61 | 0 | 10 |
| 10 | 32.1 | 7 | 920.50 | 920.50 | 920.50 | 0 | 8 |
| 11 | 32.1 | 4 | 994.57 | 994.57 | 994.57 | 0 | 9 |
| 12 | 32.1 | 100 | 1110.67 | 1110.68 | 1110.66 | 0 | 10 |
| 13 | 38.9 | 4 | 1136.68 | 1136.68 | 1136.68 | 0 | 10 |
| 14 | 43.4 | 62 | 2739.63 | 2739.64 | 2739.64 | 0 | 25 |
| 15 | 43.4 | 12 | 2569.52 | 2569.53 | 2569.53 | 0 | 23 |
| 16 | 43.6 | 17 | 2978.60 | 2978.61 | 2978.61 | 0 | 27 |
| 17 | 44.7 | 5 | 2850.72 | 2850.72 | 2850.73 | 0 | 26 |
| 18 | 46.3 | 3 | 2987.79 | 2987.79 | 2987.79 | 0 | 27 |
| 19 | 51.5 | 3 | 2823.76 | 2823.77 | 2823.77 | 0 | 25 |
| 20 | 51.5 | 19 | 2840.79 | 2840.80 | 2840.80 | 0 | 26 |
| 21 | 54.5 | 92 | 2174.27 | 2174.27 | 2174.28 | 0 | 20 |
| 22 | 54.6 | 3 | 2218.22 | 2218.23 | 2218.23 | 0 | 20 |
| 23 | 54.6 | 45 | 2196.24 | 2196.25 | 2196.25 | 0 | 20 |
| 24 | 54.6 | 18 | 2212.21 | 2212.21 | 2212.21 | 0 | 20 |
| 25 | 54.6 | 4 | 2042.10 | 2042.11 | 2042.11 | 0 | 18 |
| 26 | 55.2 | 7 | 2840.79 | 2840.80 | 2840.80 | 0 | 26 |
| Treatment | Concentration (mg·mL−1) | % Survival | Significance a | |
|---|---|---|---|---|
| Bovine serum albumin (BSA) b | 16 | 78.3 | ns | |
| 4 | 84.7 | ns | ||
| 1 | 91.7 | p < 0.05 | ||
| Crude venom | Manica rubida | 1 | 0.0 | p < 0.0001 |
| 0.1 | 33.3 | p < 0.0001 | ||
| 0.01 | 66.7 | ns | ||
| Myrmica rubra | 1 | 20.0 | p < 0.0001 | |
| 0.1 | 71.7 | ns | ||
| 0.01 | 66.7 | ns | ||
| Peptides | U-MYRTX-MANr1 (Manica rubida) | 16 | 6.7 | p < 0.0001 |
| 4 | 30.0 | p < 0.0001 | ||
| 1 | 75.0 | ns | ||
| U-MYRTX-MYRrub1 (Myrmica rubra) | 16 | 41.7 | p < 0.0001 | |
| 4 | 60.0 | p < 0.01 | ||
| 1 | 80.0 | ns | ||
| Treatment | Immediately Post-Injection | 1 h Post-Injection |
|---|---|---|
| Control (water) | Immediate righting; responsiveness < 10 s; fast and active movement | No visible impact on vitality or fitness |
| U-MYRTX-MANr1 16 mg·mL−1 | Delayed righting Response; ~1 min; Disoriented and slow movement (mild paralysis) | Moderate reduction of vitality and fitness; continuous mild paralysis |
| Crude venom 1 mg·mL−1 | Delayed or no righting (>3 min); very limited movement (strong paralysis) | Extreme reduction of vitality and fitness (~40% with no physiological reactions); continuous strong paralysis; necrosis and extensive loss of hemolymph |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heep, J.; Skaljac, M.; Grotmann, J.; Kessel, T.; Seip, M.; Schmidtberg, H.; Vilcinskas, A. Identification and Functional Characterization of a Novel Insecticidal Decapeptide from the Myrmicine Ant Manica rubida. Toxins 2019, 11, 562. https://doi.org/10.3390/toxins11100562
Heep J, Skaljac M, Grotmann J, Kessel T, Seip M, Schmidtberg H, Vilcinskas A. Identification and Functional Characterization of a Novel Insecticidal Decapeptide from the Myrmicine Ant Manica rubida. Toxins. 2019; 11(10):562. https://doi.org/10.3390/toxins11100562
Chicago/Turabian StyleHeep, John, Marisa Skaljac, Jens Grotmann, Tobias Kessel, Maximilian Seip, Henrike Schmidtberg, and Andreas Vilcinskas. 2019. "Identification and Functional Characterization of a Novel Insecticidal Decapeptide from the Myrmicine Ant Manica rubida" Toxins 11, no. 10: 562. https://doi.org/10.3390/toxins11100562
APA StyleHeep, J., Skaljac, M., Grotmann, J., Kessel, T., Seip, M., Schmidtberg, H., & Vilcinskas, A. (2019). Identification and Functional Characterization of a Novel Insecticidal Decapeptide from the Myrmicine Ant Manica rubida. Toxins, 11(10), 562. https://doi.org/10.3390/toxins11100562