Channel Formation by LktA of Mannheimia (Pasteurella) haemolytica in Lipid Bilayer Membranes and Comparison of Channel Properties with Other RTX-Cytolysins


Abstract
:1. Introduction
2. Results
2.1. Purification of LktA
2.2. LktA Obtained by Polyethylene Glycol 4000 Precipitation Had an Extremely Low Membrane Activity
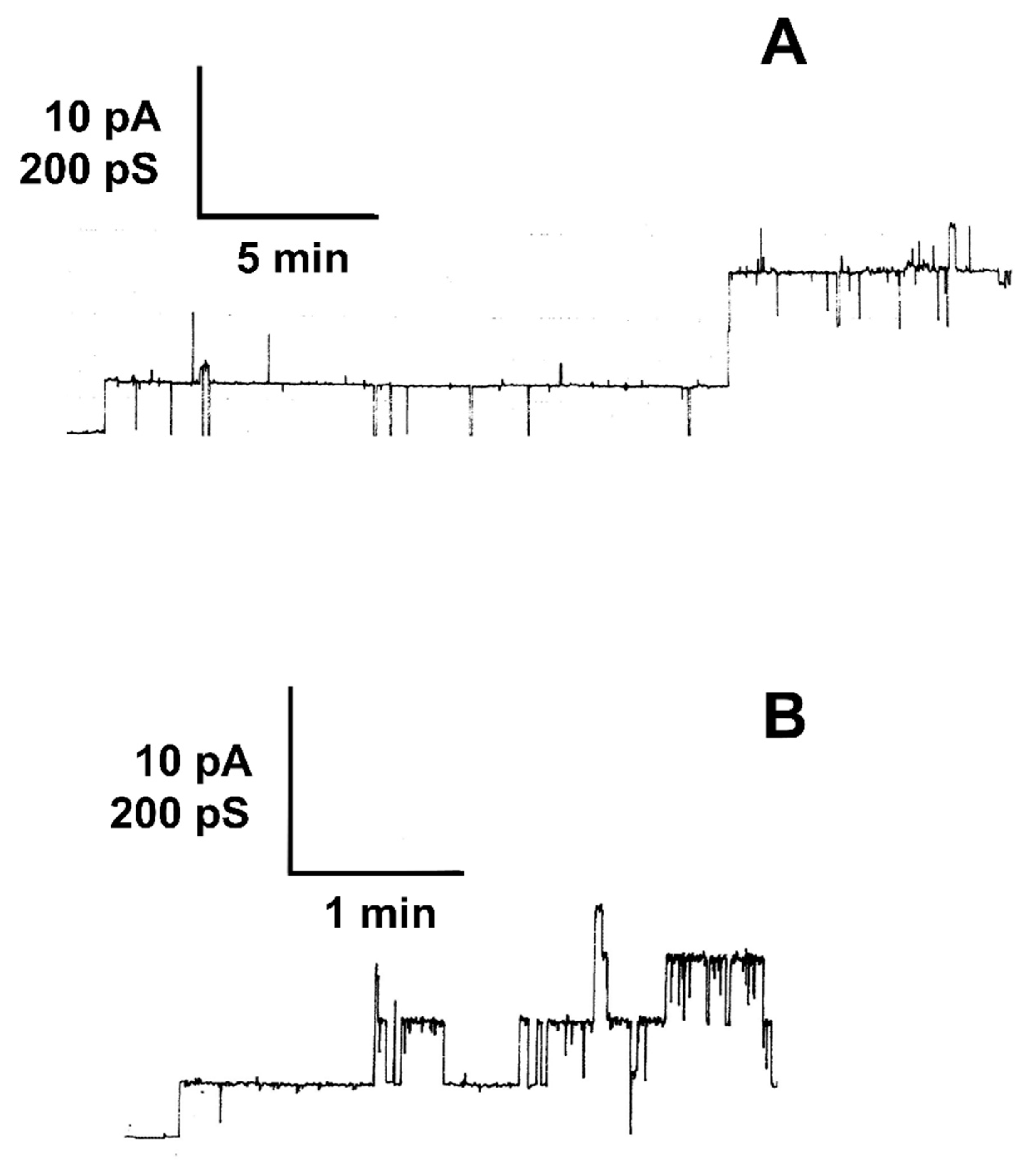
2.3. LktA Dissolved in 6 M Urea Formed Small Ion-Permeable Channels
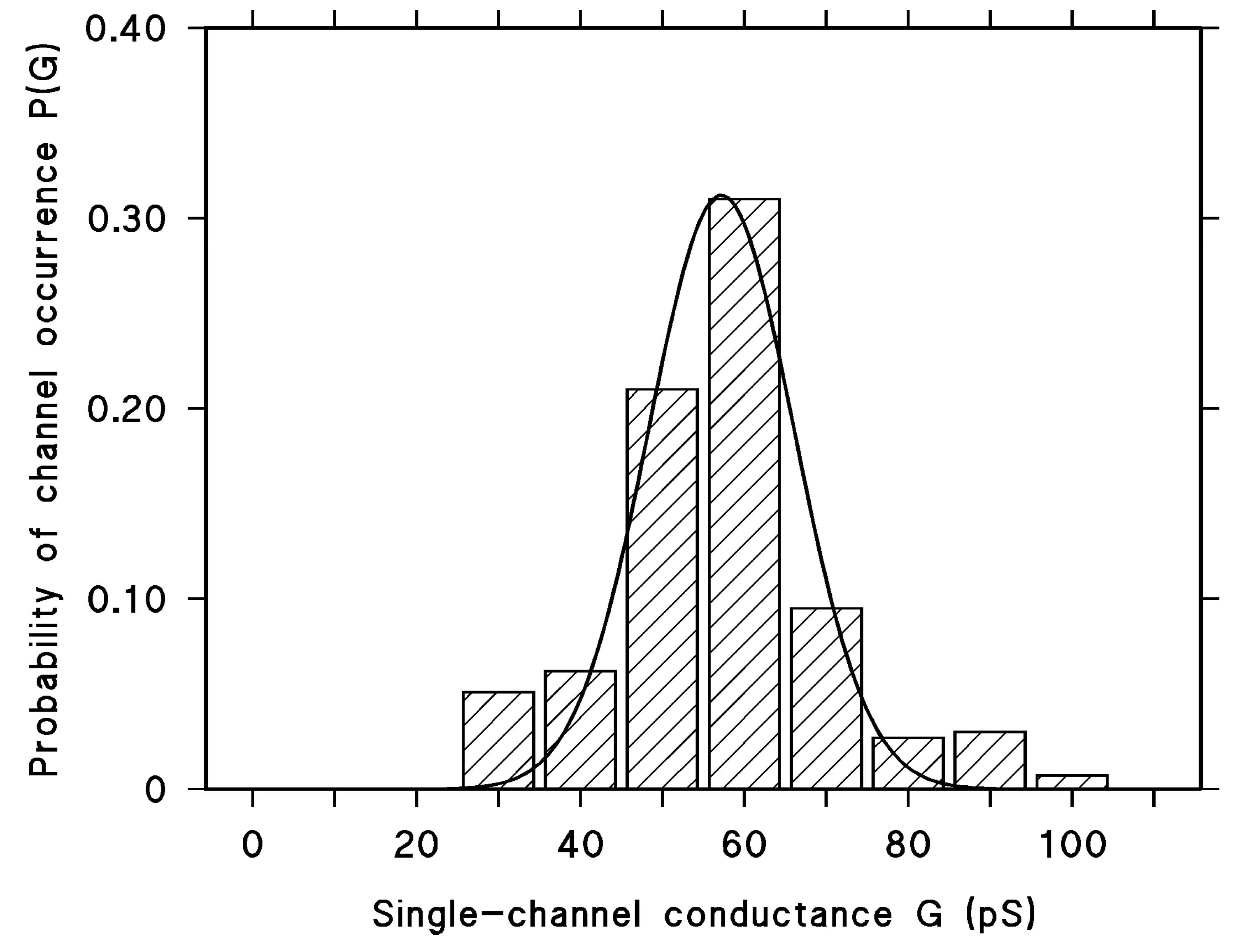
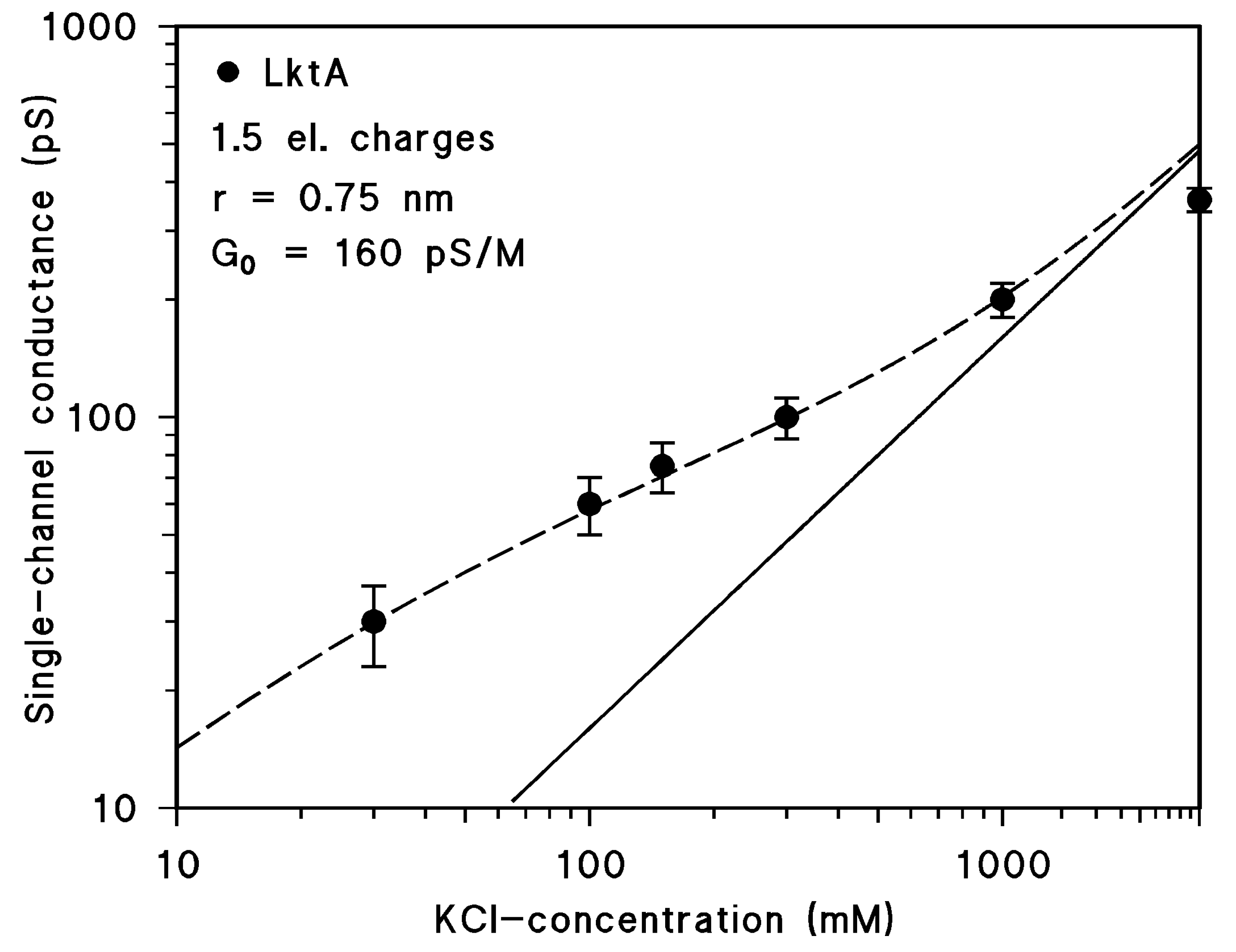
2.4. Results of Single-Channel Analysis
2.5. Selectivity of the LktA Channel
3. Discussion
3.1. Purification of LktA from E. coli AA525
3.2. Single-Channel Analysis
3.3. Negative Point Charges Control Ion Transport through LktA Channels
3.4. Comparison with Channel Properties of Other RTX-Toxins
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Culture Conditions
4.2. Isolation and Purification of LktA
4.3. SDS‑PAGE
4.4. Lipid Bilayer Experiments
4.5. Effects of Point Charges on LktA Conductance
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lo, R.Y.; Sathiamoorthy, S.; Shewen, P.E. Analysis of in vivo expressed genes in Mannheimia haemolytica A1. FEMS Microbiol. Lett. 2006, 265, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.A.; Abdelsalam, E.B. A review on pneumonic pasteurellosis (respiratory mannheimiosis) with emphasis on pathogenesis, virulence mechanisms and predisposing factors. Bulg. J. Vet. Med. 2008, 11, 139–160. [Google Scholar]
- Oppermann, T.; Schwarz, S.; Busse, N.; Czermak, P. A fast and simple assay to quantify bacterial leukotoxin activity. EJBT 2016, 24, 38–42. [Google Scholar] [CrossRef]
- Gilmour, N.J.L.; Gilmour, J.S. Pasteurellosis of sheep. In Pasteurella and Pasteurellosis; Adlam, C.F., Rutter, J.M., Eds.; Academic Press: London, UK, 1989; pp. 223–261. [Google Scholar]
- Highlander, S.K. Molecular genetic analysis of virulence in Mannheimia (Pasteurella) haemolytica. Front. Biosci. 2001, 6, D1128–D1150. [Google Scholar] [PubMed]
- Davies, R.L.; Donachie, W. Intra-specific diversity and host specificity within Pasteurella haemolytica based on variation of capsular polysaccharide, lipopolysaccharide and outer-membrane proteins. Microbiology 1996, 142, 1895–1907. [Google Scholar] [CrossRef] [PubMed]
- Davies, R.L.; Arkinsaw, S.; Selander, R.K. Evolutionary genetics of Pasteurella haemolytica isolates recovered from cattle and sheep. Infect. Immun. 1997, 65, 3585–3593. [Google Scholar] [PubMed]
- Ewers, C.; Lübke-Becker, A.; Wiele, L.H. Mannheimia haemolytica und die Pathogenese der Enzootischen Bronchopneumonie. Berl. Münch. Tierärztl. Wschr. 2004, 117, 97–115. [Google Scholar]
- Potter, A.A.; Ready, K.; Gilchrist, J. Purification of fimbriae from Pasteurella haemolytica A-1. Microb. Pathog. 1988, 4, 311–316. [Google Scholar] [CrossRef]
- Highlander, S.K.; Chidambaram, M.; Engler, M.J.; Weinstock, G.M. DNA sequence of the Pasteurella haemolytica leukotoxin gene cluster. DNA 1989, 8, 15–28. [Google Scholar] [CrossRef]
- Potter, M.D.; Lo, R.Y. Cloning and characterization of a gene from Pasteurella haemolytica A1 involved in lipopolysaccharide biosynthesis. FEMS Microbiol. Lett. 1995, 129, 75–81. [Google Scholar]
- Davies, R.L.; Baillie, S. Cytotoxic activity of Mannheimia haemolytica strains in relation to diversity of the leukotoxin structural gene lktA. Vet. Microbiol. 2003, 92, 263–279. [Google Scholar] [CrossRef]
- Lo, R.Y. Genetic analysis of virulence factors of Mannheimia (Pasteurella) haemolytica A1. Vet. Microbiol. 2001, 83, 23–35. [Google Scholar] [CrossRef]
- Rice, J.; Carrasco-Medina, L.; Hodgins, D.; Shewen, P. Mannheimia haemolytica and bovine respiratory disease. Anim. Health Res. Rev. 2007, 8, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, L.O.; Maheswaran, S.K.; Weiss, D.J.; Ames, T.R. Immunohistochemical localization of Pasteurella haemolytica A1-derived endotoxin, leukotoxin, and capsular polysaccharide in experimental bovine Pasteurella pneumonia. Vet. Pathol. 1990, 27, 150–161. [Google Scholar] [CrossRef]
- Lo, R.Y.; McKerral, L.J.; Hills, T.L.; Kostrzynska, M. Analysis of the capsule biosynthetic locus of Mannheimia (Pasteurella) haemolytica A1 and proposal of a nomenclature system. Infect. Immun. 2001, 69, 4458–4464. [Google Scholar] [CrossRef]
- Highlander, S.K.; Fedorova, N.D.; Dusek, D.M.; Panciera, R.; Alvarez, L.E.; Rinehart, C. Inactivation of Pasteurella (Mannheimia) haemolytica leukotoxin causes partial attenuation of virulence in a calf challenge model. Infect. Immun. 2000, 68, 3916–3922. [Google Scholar] [CrossRef]
- Jeyaseelan, S.; Sreevatsan, S.; Maheswaran, S.K. Role of Mannheimia haemolytica leukotoxin in the pathogenesis of bovine pneumonic pasteurellosis. Anim. Health Res. Rev. 2002, 3, 69–82. [Google Scholar] [CrossRef]
- Linhartová, I.; Bumba, L.; Mašín, J.; Basler, M.; Osička, R.; Kamanová, J.; Procházková, K.; Adkins, I.; Hejnová-Holubová, J.; Sadílková, L.; et al. RTX proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef]
- Welch, R.A. Pore-forming cytolysins of gram-negative bacteria. Mol. Microbiol. 1991, 5, 521–528. [Google Scholar] [CrossRef]
- Welch, R.A.; Forestier, C.; Lobo, A.; Pellett, S.; Thomas, W., Jr.; Rowe, G. The synthesis and function of the Escherichia coli hemolysin and related RTX exotoxins. FEMS Microbiol. Immunol. 1992, 5, 29–36. [Google Scholar] [CrossRef]
- Omaleki, L.; Browning, G.F.; Barber, S.R.; Allen, J.L.; Srikumaran, S.; Markham, P.F. Sequence diversity, cytotoxicity and antigenic similarities of the leukotoxin of isolates of Mannheimia species from mastitis in domestic sheep. Vet Microbiol. 2014, 174, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Davies, R.L.; Campbell, S.; Whittam, T.S. Mosaic Structure and Molecular Evolution of the Leukotoxin Operon (lktCABD) in Mannheimia (Pasteurella) haemolytica, Mannheimia glucosida, and Pasteurella trehalosi. J. Bacteriol. 2002, 184, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Baumann, U.; Wu, S.; Flaherty, K.M.; McKay, D.B. Three-dimensional structure of the alkaline protease of Pseudomonas aeruginosa: A two-domain protein with a calcium binding parallel beta roll motif. EMBO J. 1993, 12, 3357–3364. [Google Scholar] [CrossRef] [PubMed]
- Döbereiner, A.; Schmid, A.; Ludwig, A.; Goebel, W.; Benz, R. The effects of calcium and other polyvalent cations on channel formation by Escherichia coli alpha-hemolysin in red blood cells and lipid bilayer membranes. Eur. J. Biochem. 1996, 240, 454–460. [Google Scholar] [CrossRef]
- Highlander, S.K.; Engler, M.J.; Weinstock, G.M. Secretion and expression of the Pasteurella haemolytica Leukotoxin. J. Bacteriol. 1990, 172, 2343–2350. [Google Scholar] [CrossRef]
- Lally, E.T.; Hill, R.B.; Kieba, I.R.; Korostoff, J. The interaction between RTX toxins and target cells. Trends Microbiol. 1999, 7, 356–361. [Google Scholar] [CrossRef]
- Benz, R.; Maier, E.; Gentschev, I. TolC of Escherichia coli functions as an outer membrane channel. Zent. Bakteriol. 1993, 278, 187–196. [Google Scholar] [CrossRef]
- Koronakis, V.; Sharff, A.; Koronakis, E.; Luisi, B.; Hughes, C. Crystal structure of the bacterial membrane protein TolC central to multidrug efflux and protein export. Nature 2000, 405, 914–919. [Google Scholar] [CrossRef]
- Mackman, N.; Nicaud, J.M.; Gray, L.; Holland, I.B. Genetical and functional organisation of the Escherichia coli haemolysin determinant 2001. Mol. Gen. Genet. 1985, 201, 282–288. [Google Scholar] [CrossRef]
- Holland, I.B.; Schmitt, L.; Young, J. Type 1 protein secretion in bacteria, the ABC-transporter dependent pathway (review). Mol. Membr. Biol. 2005, 22, 29–39. [Google Scholar] [CrossRef]
- Sun, Y.; Clinkenbeard, K.D.; Clarke, C.; Cudd, L.; Highlander, S.K.; Dabo, S.M. Pasteurella haemolytica leukotoxin induced apoptosis of bovine lymphocytes involves DNA fragmentation. Vet. Microbiol. 1999, 65, 153–166. [Google Scholar] [CrossRef]
- Sun, Y.; Clinkenbeard, K.D.; Cudd, L.A.; Clarke, C.R.; Clinkenbeard, P.A. Correlation of Pasteurella haemolytica leukotoxin binding with susceptibility to intoxication of lymphoid cells from various species. Infect. Immun. 1999, 67, 6264–6269. [Google Scholar] [PubMed]
- Jeyaseelan, S.; Hsuan, S.L.; Kannan, M.S.; Walcheck, B.; Wang, J.F.; Kehrli, M.E.; Lally, E.T.; Sieck, G.C.; Maheswaran, S.K. Lymphocyte function-associated antigen 1 is a receptor for Pasteurella haemolytica leukotoxin in bovine leukocytes. Infect. Immun. 2000, 68, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Thumbikat, P.; Dileepan, T.; Kannan, M.S.; Maheswaran, S.K. Characterization of Mannheimia (Pasteurella) haemolytica leukotoxin interaction with bovine alveolar macrophage beta2 integrins. Vet. Res. 2005, 36, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Dileepan, T.; Thumbikat, P.; Walcheck, B.; Kannan, M.S.; Maheswaran, S.K. Recombinant expression of bovine LFA-1 and characterization of its role as a receptor for Mannheimia haemolytica leukotoxin. Microb. Pathog. 2005, 38, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Clinkenbeard, K.D.; Mosier, D.A.; Confer, A.W. Transmembrane pore size and role of cell swelling in cytotoxicity caused by Pasteurella haemolytica leukotoxin. Infect. Immun. 1989, 57, 420–425. [Google Scholar]
- Sun, Y.; Clinkenbeard, K.D.; Ownby, C.L.; Cudd, L.; Clarke, C.R.; Highlander, S.K. Ultrastructural characterization of apoptosis in bovine lymphocytes exposed to Pasteurella haemolytica leukotoxin. Am. J. Vet. Res. 2000, 61, 51–55. [Google Scholar] [CrossRef]
- Singh, K.; Ritchey, J.W.; Confer, A.W. Mannheimia haemolytica: Bacterial–host interactions in bovine pneumonia. Vet. Pathol. 2011, 48, 338–348. [Google Scholar] [CrossRef]
- Benz, R.; Maier, E.; Ladant, D.; Ullmann, A.; Sebo, P. Adenylate cyclase toxin (CyaA) of Bordetella pertussis. Evidence for the formation of small ion-permeable channels and comparison with HlyA of Escherichia coli. J. Biol. Chem. 1994, 269, 27231–27239. [Google Scholar]
- Bárcena-Uribarri, I.; Benz, R.; Winterhalter, M.; Zakharian, E.; Balashova, N. Pore forming activity of the potent RTX-toxin produced by pediatric pathogen Kingella kingae: Characterization and comparison to other RTX-family members. Biochim. Biophys. Acta 2015, 1848, 1536–1544. [Google Scholar] [CrossRef]
- Benz, R. Channel formation by RTX-toxins of pathogenic bacteria: Basis of their biological activity. Biochim. Biophys. Acta 2016, 1858, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Benz, R.; Schmid, A.; Wagner, W.; Goebel, W. Pore formation by the Escherichia coli hemolysin: Evidence for an association-dissociation equilibrium of the pore-forming aggregates. Infect. Immun. 1989, 57, 887–895. [Google Scholar] [PubMed]
- Benz, R.; Hardie, K.R.; Hughes, C. Pore formation in artificial membranes by the secreted hemolysins of Proteus vulgaris and Morganella morganii. Eur. J. Biochem. 1994, 220, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Maier, E.; Reinhard, N.; Benz, R.; Frey, J. Channel-forming activity and channel size of the RTX toxins ApxI, ApxII, and ApxIII of Actinobacillus pleuropneumoniae. Infect. Immun. 1996, 64, 4415–4423. [Google Scholar] [PubMed]
- Betsou, F.; Sebo, P.; Guiso, N. CyaC-mediated activation is important not only for toxic but also for protective activities of Bordetella pertussis adenylate cyclase-hemolysin. Infect. Immun. 1993, 61, 3583–3589. [Google Scholar]
- Knapp, O.; Maier, E.; Polleichtner, G.; Masín, J.; Sebo, P.; Benz, R. Channel formation in model membranes by the adenylate cyclase toxin of Bordetella pertussis: Effect of calcium. Biochemistry 2003, 42, 8077–8084. [Google Scholar] [CrossRef]
- Saza, H.; Bartolomé, B.; Serra, J.L.; de la Cruz, F.; Goñi, F.M. Alpha-haemolysin from E. coli. Purification and self-aggregation properties. FEBS Lett. 1991, 280, 195–198. [Google Scholar]
- Castellan, E.G. The ionic current in aqueous solutions. In Physical Chemistry, 3rd ed.; Addison-Wesley Publishing Company: Reading, MA, USA, 1983; pp. 769–780. [Google Scholar]
- Benz, R.; Janko, K.; Lauger, P. Ionic selectivity of pores formed by the matrix protein (porin) of Escherichia coli. Biochim. Biophys. Acta 1979, 551, 238–247. [Google Scholar] [CrossRef]
- Masin, J.; Basler, M.; Knapp, O.; El-Azami-El-Idrissi, M.; Maier, E.; Konopasek, I.; Benz, R.; Leclerc, C.; Sebo, P. Acylation of lysine 860 allows tight binding and cytotoxicity of Bordetella adenylate cyclase on CD11b-expressing cells. Biochemistry 2005, 44, 12759–12766. [Google Scholar] [CrossRef]
- Schmidt, H.; Maier, E.; Karch, H.; Benz, R. Pore-forming properties of the plasmid-encoded hemolysin of enterohemorrhagic Escherichia coli O157:H7. Eur. J. Biochem. 1996, 241, 594–601. [Google Scholar] [CrossRef]
- Osickova, A.; Osicka, R.; Maier, E.; Benz, R.; Sebo, P. An amphipathic α-helix including glutamates 509 and 516 is crucial for membrane translocation of adenylate cyclase toxin and modulates formation and cation selectivity of its membrane channels. J. Biol. Chem. 1999, 274, 37644–37650. [Google Scholar] [PubMed]
- Valeva, A.; Siegel, I.; Wylenzek, M.; Wassenaar, T.M.; Weis, S.; Heinz, N.; Schmitt, R.; Fischer, C.; Reinartz, R.; Bhakdi, S.; et al. Putative identification of an amphipathic alpha-helical sequence in hemolysin of Escherichia coli (HlyA) involved in transmembrane pore formation. Biol. Chem. 2008, 389, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- Benz, R.; Maier, E.; Bauer, S.; Ludwig, A. The deletion of several amino acid stretches of Escherichia coli alpha-hemolysin (HlyA) suggests that the channel-forming domain contains beta-strands. PLoS ONE 2014, 9, e112248. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.; Benz, R.; Goebel, W. Oligomerization of Escherichia coli haemolysin (HlyA) is involved in pore formation. Mol. Gen. Genet. 1993, 241, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Vojtova-Vodolanova, J.; Basler, M.; Osicka, R.; Knapp, O.; Maier, E.; Cerny, J.; Benada, O.; Benz, R.; Sebo, P. Oligomerization is involved in pore formation by Bordetella adenylate cyclase toxin. FASEB J. 2009, 23, 2831–2843. [Google Scholar] [CrossRef] [PubMed]
- Trias, J.; Benz, R. Characterization of the channel formed by the mycobacterial porin in lipid bilayer membranes. Demonstration of voltage gating and of negative point charges at the channel mouth. J. Biol. Chem. 1993, 268, 6234–6240. [Google Scholar]
- Scheffer, J.; König, W.; Hacker, J.; Goebel, W. Bacterial adherence and hemolysin production from Escherichia coli induces histamine and leukotriene release from various cells. Infect. Immun. 1985, 50, 271–278. [Google Scholar]
- Vázquez, R.F.; Millone, M.A.D.; Pavinatto, F.J.; Herlax, V.S.; Bakás, L.S.; Oliveira, O.N., Jr.; Vela, M.E.; Maté, S.M. Interaction of acylated and unacylated forms of E. coli alpha-hemolysin with lipid monolayers: A PM-IRRAS study. Colloids Surf. B Biointerfaces 2017, 158, 76–83. [Google Scholar] [CrossRef]
- Ludwig, A.; Garcia, F.; Bauer, S.; Jarchau, T.; Benz, R.; Hoppe, J.; Goebel, W. Analysis of the in vivo activation of hemolysin (HlyA) from Escherichia coli. J. Bacteriol. 1996, 178, 5422–5430. [Google Scholar] [CrossRef]
- Bhakdi, S.; Tranum-Jensen, J. Membrane damage by pore-forming bacterial cytolysins. Microb. Pathog. 1986, 1, 5–14. [Google Scholar] [CrossRef]
- Ehrmann, I.E.; Gray, M.C.; Gordon, V.M.; Gray, L.S.; Hewlett, E.L. Hemolytic activity of adenylate cyclase toxin from Bordetella pertussis. FEBS Lett. 1991, 278, 79–83. [Google Scholar] [PubMed]
- Lear, J.D.; Furblur, U.G.; Lally, E.T.; Tanaka, J.C. Actinobacillus actinomycetemcomitans leukotoxin forms large conductance, voltage-gated ion channels when incorporated into planar lipid bilayers. Biochim. Biophys. Acta 1995, 1238, 34–41. [Google Scholar] [CrossRef]
- Forestier, C.; Welch, R.A. Identification of RTX toxin target cell specificity domains by use of hybrid genes. Infect. Immun. 1991, 59, 4212–4220. [Google Scholar] [PubMed]
- Eberspächer, B.; Hugo, F.; Bhakdi, S. Quantitative study of the binding and hemolytic efficiency of Escherichia coli hemolysin. Infect. Immun. 1989, 57, 983–988. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Benz, R.; Janko, K.; Boos, W.; Lauger, P. Formation of large, ion-permeable membrane channels by the matrix protein (porin) of Escherichia coli. Biochim. Biophys. Acta 1978, 511, 305–319. [Google Scholar] [CrossRef]
- Janko, K.; Benz, R. Properties of lipid bilayer membranes made from lipids containing phytanic acid. Biochim. Biophys. Acta 1977, 470, 8–16. [Google Scholar] [CrossRef]
- Scholfield, C.R. Composition of Soybean Lecithin. J. Am. Oil Chem. Soc. 1981, 58, 889–892. [Google Scholar] [CrossRef]
- Benz, R.; Hancock, R.E. Mechanism of ion transport through the anion-selective channel of the Pseudomonas aeruginosa outer membrane. J. Gen. Physiol. 1987, 89, 275–295. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Electrolyte/Ion | Concentration (M) | G (pS) | Limiting Molar Conductivity (S·cm2·mol−1) |
|---|---|---|---|
| KCl | 0.03 | 30 ± 6 | |
| 0.10 | 57 ± 9 | ||
| 0.15 | 75 ± 11 | ||
| 0.30 | 100 ± 12 | ||
| 1.00 | 200 ± 20 | ||
| 3.00 | 360 ± 25 | ||
| LiCl | 1.00 | 85 ± 11 | |
| KCH3COO (pH 7) | 1.00 | 180 ± 18 | |
| K+ | 73.5 | ||
| Li+ | 38.7 | ||
| Cl− | 76.4 | ||
| CH3COO− | 40.9 |
| Electrolyte | Permeability Ratios Pcation/Panion | Vm (mV) |
|---|---|---|
| KCl | 5.0 | 31.0 ± 2.4 |
| LiCl | 4.6 | 29.8 ± 1.8 |
| KCH3COO (pH 7) | 7.3 | 38.1 ± 2.3 |
| Toxin | SCC | IS | VG | PD |
|---|---|---|---|---|
| LktA | 75 pS | Cation‑select. | No voltage dependence (this study) | 1.5 nm (d) (this study) |
| M. haemolytica | (0.15 M KCl) (this study) | PK/PCl = 5.0 (this study) | ||
| HlyA | 500 pS | Cation‑select. PK/PCl = 9 [43] | <100 mV [43] | 1.4–3 nm(b) [62] |
| E. coli | (0.15 M KCl) [43] | |||
| EHEC-Hly | 500 pS | Cation‑select. PK/PCl = 13 [52] | No data | 2.6 nm (b,c) [52] |
| E. coli EHEC | (0.15 M KCl) [52] | |||
| RtxA | 400 pS | Cation‑select. | 30–40 mV [41] | 1.9 nm (a) [41] |
| K. kingae | (0.1 M KCl) [41] | PK/PCl = 3.5 [41] | ||
| ApxI | 540 pS | Cation‑select. | No data | 2.4 nm (c) [45] |
| A. pleuropneumoniae | (0.15 M KCl) [45] | PK/PCl = 5.7 [45] | ||
| ApxII | 620 pS | Cation‑select. | No data | 2.5 nm (c) [45] |
| A. pleuropneumoniae | (0.15 M KCl) [45] | PK/PCl = No data [45] | ||
| ApxIII | 95 pS | Cation‑select. PK/PCl = 9.6 [45] | No data | 1.8 nm (c) [45] |
| A. pleuropneumoniae | (0.15 M KCl) [45] | |||
| CyaA | 10 pS | Cation‑select. PK/PCl = 9–11 [40] | No data | 0.6–0.8 nm (b,c) [40,63] |
| B. pertussis | (0.1 M KCl) [40] | |||
| Rtx | 500 pS | Cation‑select. (PK/PCl = 9.5) [44] | ≥40 mV [44] | 2 nm (d) [44] |
| P. vulgaris | (0.15 M KCl) [44] | |||
| Rtx | 520 pS | Cation‑select. (PK/PCl = 10) [44] | ≥40 mV [44] | 2 nm (d) [44] |
| M. morganii | (0.15 M KCl) [44] | |||
| Leukotoxin | 406 pS | No data | No data | No data |
| Actinobacillus actinomycetemcomitans | (0.14 M NaCl, 0.01 M CaCl2) [64] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benz, R.; Piselli, C.; Potter, A.A. Channel Formation by LktA of Mannheimia (Pasteurella) haemolytica in Lipid Bilayer Membranes and Comparison of Channel Properties with Other RTX-Cytolysins. Toxins 2019, 11, 604. https://doi.org/10.3390/toxins11100604
Benz R, Piselli C, Potter AA. Channel Formation by LktA of Mannheimia (Pasteurella) haemolytica in Lipid Bilayer Membranes and Comparison of Channel Properties with Other RTX-Cytolysins. Toxins. 2019; 11(10):604. https://doi.org/10.3390/toxins11100604
Chicago/Turabian StyleBenz, Roland, Claudio Piselli, and Andrew A. Potter. 2019. "Channel Formation by LktA of Mannheimia (Pasteurella) haemolytica in Lipid Bilayer Membranes and Comparison of Channel Properties with Other RTX-Cytolysins" Toxins 11, no. 10: 604. https://doi.org/10.3390/toxins11100604
APA StyleBenz, R., Piselli, C., & Potter, A. A. (2019). Channel Formation by LktA of Mannheimia (Pasteurella) haemolytica in Lipid Bilayer Membranes and Comparison of Channel Properties with Other RTX-Cytolysins. Toxins, 11(10), 604. https://doi.org/10.3390/toxins11100604