Inhibitory Effect of Metalloproteinase Inhibitors on Skin Cell Inflammation Induced by Jellyfish Nemopilema nomurai Nematocyst Venom

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Effect of NnNV and NnNV-I on the Viability of HaCaT and CCC-ESF-1
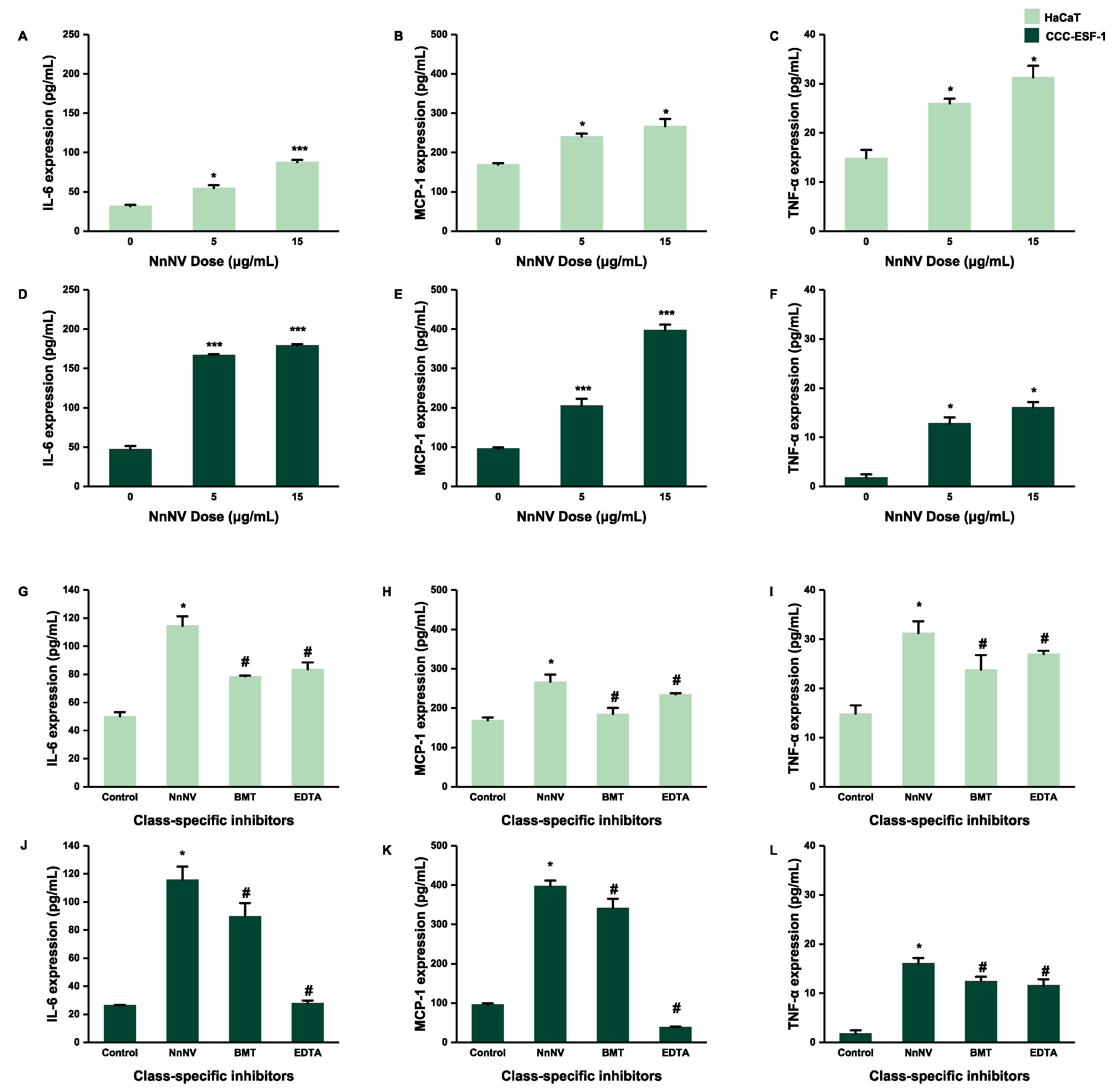
2.2. Effect of NnNV on the Gene Expression of IL-6, MCP-1, and TNF-α in HaCaT and CCC-ESF-1 at Different Times
2.3. Effect of NnNV and NnNV-I on the Gene Expression of IL-6, MCP-1 and TNF-α in HaCaT and CCC-ESF-1
2.4. Effect of NnNV and NnNV-I on the Protein Expression of IL-6, MCP-1 and TNF-α in HaCaT and CCC-ESF-1
3. Discussion
4. Materials and Methods
4.1. Jellyfish Collection
4.2. Nematocyst Isolation and Venom Extraction
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. RNA Isolation and Quantitative Real Time PCR (qRT-PCR)
4.6. ELISA Detection of Cytokines
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| NnNV | jellyfish Nemopilema nomurai nematocyst venom |
| NnNV-I | the mixtures of jellyfish Nemopilema nomurai nematocyst venom and matrix metalloproteinase inhibitors batimastat or ethylenediaminetetraacetic acid |
| BMT | batimastat |
| EDTA | ethylenediaminetetraacetic acid |
| PBS | phosphate-buffered saline |
| FBS | fetal bovine serum |
| MMP | matrix metalloproteinase |
| VEB | venom extraction buffer |
References
- Purcell, J.E.; Uye, S.; Lo, W.T. Anthropogenic Causes of Jellyfish Blooms and Their Direct Consequences for Humans: A Review. Mar. Ecol. Prog. Ser. 2007, 350, 153–174. [Google Scholar] [CrossRef]
- Je, P. Climate Effects on Formation of Jellyfish and Ctenophore Blooms: A Review. J. Mar. Biol. Assoc. UK 2005, 85, 461–476. [Google Scholar]
- Mariottini, G.L.; Pane, L. Mediterranean Jellyfish Venoms: A Review on Scyphomedusae. Mar. Drugs 2010, 8, 1122–1152. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, L.; Seys, J.; Mees, J. To Pee, or Not to Pee: A Review on Envenomation and Treatment in European Jellyfish Species. Mar. Drugs 2016, 14, 127. [Google Scholar] [CrossRef] [PubMed]
- Pyo, M.J.; Lee, H.; Bae, S.K.; Heo, Y.; Choudhary, I.; Yoon, W.D.; Kang, C.; Kim, E. Modulation of Jellyfish Nematocyst Discharges and Management of Human Skin Stings in Nemopilema Nomurai and Carybdea Mora. Toxicon 2016, 109, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, M.; Uye, S.; Burnett, J.; Mianzan, H. Stings of Edible Jellyfish (Rhopilema Hispidum, Rhopilema Esculentum and Nemopilema Nomurai) in Japanese Waters. Toxicon 2006, 48, 713. [Google Scholar] [CrossRef] [PubMed]
- Glatstein, M.; Adir, D.; Galil, B.; Scolnik, D.; Rimon, A.; Pivkolevy, D.; Hoyte, C. Pediatric Jellyfish Envenomation in the Mediterranean Sea. Eur. J. Emerg. Med. 2017, 25, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Birsa, L.M.; Verity, P.G.; Lee, R.F. Evaluation of the Effects of Various Chemicals on Discharge of and Pain Caused by Jellyfish Nematocysts. Comp. Biochem. Physiol.—C Toxicol. Pharmacol. 2010, 151, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Morabito, R.; Marino, A.; Dossena, S.; La Spada, G. Nematocyst Discharge in Pelagia Noctiluca (Cnidaria, Scyphozoa) Oral Arms Can Be Affected by Lidocaine, Ethanol, Ammonia and Acetic Acid. Toxicon 2014, 83, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yu, H.; Xue, W.; Yue, Y.; Liu, S.; Xing, R.; Li, P. Jellyfish Venomics and Venom Gland Transcriptomics Analysis of Stomolophus Meleagris to Reveal the Toxins Associated with Sting. J. Proteomics 2014, 106, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Takeya, H.; Onikura, A.; Nikai, T.; Sugihara, H.; Iwanaga, S. Primary Structure of a Hemorrhagic Metalloproteinase, HT-2, Isolated from the Venom of Crotalus ruber ruber. J. Biochem. 1990, 108, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.W.; Serrano, S.M. Structural Considerations of the Snake Venom Metalloproteinases, Key Members of the M12 Reprolysin Family of Metalloproteinases. Toxicon 2005, 45, 969–985. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Rucavado, A. Snake Venom Metalloproteinases: Their Role in the Pathogenesis of Local Tissue Damage. Biochimie 2000, 82, 841–850. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the Environment: Regulation of Local and Global Homeostasis by the Skin’s Neuroendocrine System. Adv. Anat. Embryol. Cell Biol. 2012, 212, v-115. [Google Scholar] [PubMed]
- Rucavado, A.; Escalante, T.; Shannon, J.; Gutiérrez, J.M.; Fox, J.W. Proteomics of Wound Exudate in Snake Venom-Induced Pathology: Search for Biomarkers to Assess Tissue Damage and Therapeutic Success. J. Proteome Res. 2011, 10, 1987–2005. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Yu, H.; Li, R.; Xing, R.; Liu, S.; Li, K.; Wang, X.; Chen, X.; Li, P. Functional Elucidation of Nemopilema Nomurai and Cyanea Nozakii Nematocyst Venoms’ Lytic Activity Using Mass Spectrometry and Zymography. Toxins 2017, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, D.; Liu, G.; Zhang, X.; Wang, Q.; Zheng, J.; Zhou, Y.; He, Q.; Zhang, L. Protective Effects of Batimastat against Hemorrhagic Injuries in Delayed Jellyfish Envenomation Syndrome Models. Toxicon 2015, 108, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Manna, P.R.; Tuckey, R.C. On the Role of Skin in the Regulation of Local and Systemic Steroidogenic Activities. Steroids 2015, 103, 72–88. [Google Scholar] [CrossRef] [PubMed]
- Locksley, R.M.; Killeen, N.; Lenardo, M.J. The TNF and TNF Receptor Superfamilies: Integrating Mammalian Biology. Cell 2001, 104, 487–501. [Google Scholar] [CrossRef]
- Sehgal, P.B.; Wang, L.; Rayanade, R.; Pan, H.; Margulies, L. Interleukin-6-Type Cytokines. Ann. N.Y. Acad. Sci. 1995, 762, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.S.; Angelo, L.S.; Kurzrock, R. Interleukin-6 and Its Receptor in Cancer: Implications for Translational Therapeutics. Cancer 2007, 110, 1911–1928. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.-H.; Jin, F.; Kwak, C.-H.; Abekura, F.; Park, J.-Y.; Park, N.G.; Chang, Y.-C.; Lee, Y.-C.; Chung, T.-W.; Ha, K.-T.; et al. Jellyfish Extract Induces Apoptotic Cell Death through the P38 Pathway and Cell Cycle Arrest in Chronic Myelogenous Leukemia K562 Cells. PeerJ 2017, 5, e2895. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Fan, L.L.; Zhong, T.; Li, G.; Xia, X.; Long, H.; Huang, D.; Shu, W. The Nematocysts Venom of Chrysaora Helvola Brandt Leads to Apoptosis-like Cell Death Accompanied by Uncoupling of Oxidative Phosphorylation. Toxicon 2016, 110, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Stella, V.J.; He, Q. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2008, 35, 495–516. [Google Scholar]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S. The Toxicogenomic Multiverse: Convergent Recruitment of Proteins into Animal Venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [PubMed]
- Undheim, E.A.; Jones, A.; Clauser, K.R.; Holland, J.W.; Pineda, S.S.; King, G.F.; Fry, B.G. Clawing through Evolution: Toxin Diversification and Convergence in the Ancient Lineage Chilopoda (Centipedes). Mol. Biol. Evol. 2014, 31, 2124–2148. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Casewell, N.R.; Yanagihara, A.A.; Nouwens, A.; Cribb, B.W.; Whitehead, D.; Jackson, T.N.W.; Ali, S.A.; Wagstaff, S.C.; Koludarov, I.; et al. Firing the Sting: Chemically Induced Discharge of Cnidae Reveals Novel Proteins and Peptides from Box Jellyfish (Chironex Fleckeri) Venom. Toxins 2015, 7, 936–950. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Jung, E.-s.; Kang, C.; Yoon, W.D.; Kim, J.S.; Kim, E. Scyphozoan Jellyfish Venom Metalloproteinases and Their Role in the Cytotoxicity. Toxicon 2011, 58, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yu, H.; Liu, S.; Xing, R.; Guo, Z.; Li, P. Factors Affecting the Protease Activity of Venom from Jellyfish Rhopilema Esculentum Kishinouye. Bioorg. Med. Chem. Lett. 2005, 15, 5370–5374. [Google Scholar] [PubMed]
- Li, R.; Yu, H.; Yue, Y.; Li, P. Combined Proteome and Toxicology Approach Reveals the Lethality of Venom Toxins from Jellyfish Cyanea Nozakii. J. Proteome Res. 2018, 17, 3904–3913. [Google Scholar] [PubMed]
- Remigante, A.; Costa, R.; Morabito, R.; La Spada, G.; Marino, A.; Dossena, S. Impact of Scyphozoan Venoms on Human Health and Current First Aid Options for Stings. Toxins 2018, 10, 133. [Google Scholar] [CrossRef] [PubMed]
- Bloom, D.A.; Burnett, J.W.; Alderslade, P. Partial Purification of Box Jellyfish (Chironex Fleckeri) Nematocyst Venom Isolated at the Beachside. Toxicon 1998, 36, 1075–1085. [Google Scholar] [CrossRef]
- Carrette, T.; Seymour, J. A Rapid and Repeatable Method for Venom Extraction from Cubozoan Nematocysts. Toxicon 2004, 44, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yu, H.; Yue, Y.; Liu, S.; Xing, R.; Chen, X.; Wang, X.; Li, P. In Depth Analysis of the in Vivo Toxicity of Venom from the Jellyfish Stomolophus Meleagris. Toxicon 2014, 92, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Pairs | |
|---|---|
| β-actin | F: 5′-GGCACCACACCTTCTACAATGAGC-3′ |
| R: 5′-GATAGCACAGCCTGGATAGCAACG-3′ | |
| IL-6 | F: 5′-GTCCAGTTGCCTTCTCCC-3′ |
| R: 5′-GCCTCTTTGCTGCTTTCA-3′ | |
| MCP-1 | F: 5′-CTTCTGTGCCTGCTGCTC-3′ |
| R: 5′-TGCTGCTGGTGATTCTTCT-3′ | |
| TNF-α | F: 5′-CGTGGAGCTGGCCGAGGAG-3′ |
| R: 5′-AGGAAGGAGAAGAGGCTGAGGAAC-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, A.; Yu, H.; Li, R.; Liu, S.; Xing, R.; Li, P. Inhibitory Effect of Metalloproteinase Inhibitors on Skin Cell Inflammation Induced by Jellyfish Nemopilema nomurai Nematocyst Venom. Toxins 2019, 11, 156. https://doi.org/10.3390/toxins11030156
Li A, Yu H, Li R, Liu S, Xing R, Li P. Inhibitory Effect of Metalloproteinase Inhibitors on Skin Cell Inflammation Induced by Jellyfish Nemopilema nomurai Nematocyst Venom. Toxins. 2019; 11(3):156. https://doi.org/10.3390/toxins11030156
Chicago/Turabian StyleLi, Aoyu, Huahua Yu, Rongfeng Li, Song Liu, Ronge Xing, and Pengcheng Li. 2019. "Inhibitory Effect of Metalloproteinase Inhibitors on Skin Cell Inflammation Induced by Jellyfish Nemopilema nomurai Nematocyst Venom" Toxins 11, no. 3: 156. https://doi.org/10.3390/toxins11030156
APA StyleLi, A., Yu, H., Li, R., Liu, S., Xing, R., & Li, P. (2019). Inhibitory Effect of Metalloproteinase Inhibitors on Skin Cell Inflammation Induced by Jellyfish Nemopilema nomurai Nematocyst Venom. Toxins, 11(3), 156. https://doi.org/10.3390/toxins11030156