Proanthocyanidins Alleviates AflatoxinB1-Induced Oxidative Stress and Apoptosis through Mitochondrial Pathway in the Bursa of Fabricius of Broilers

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
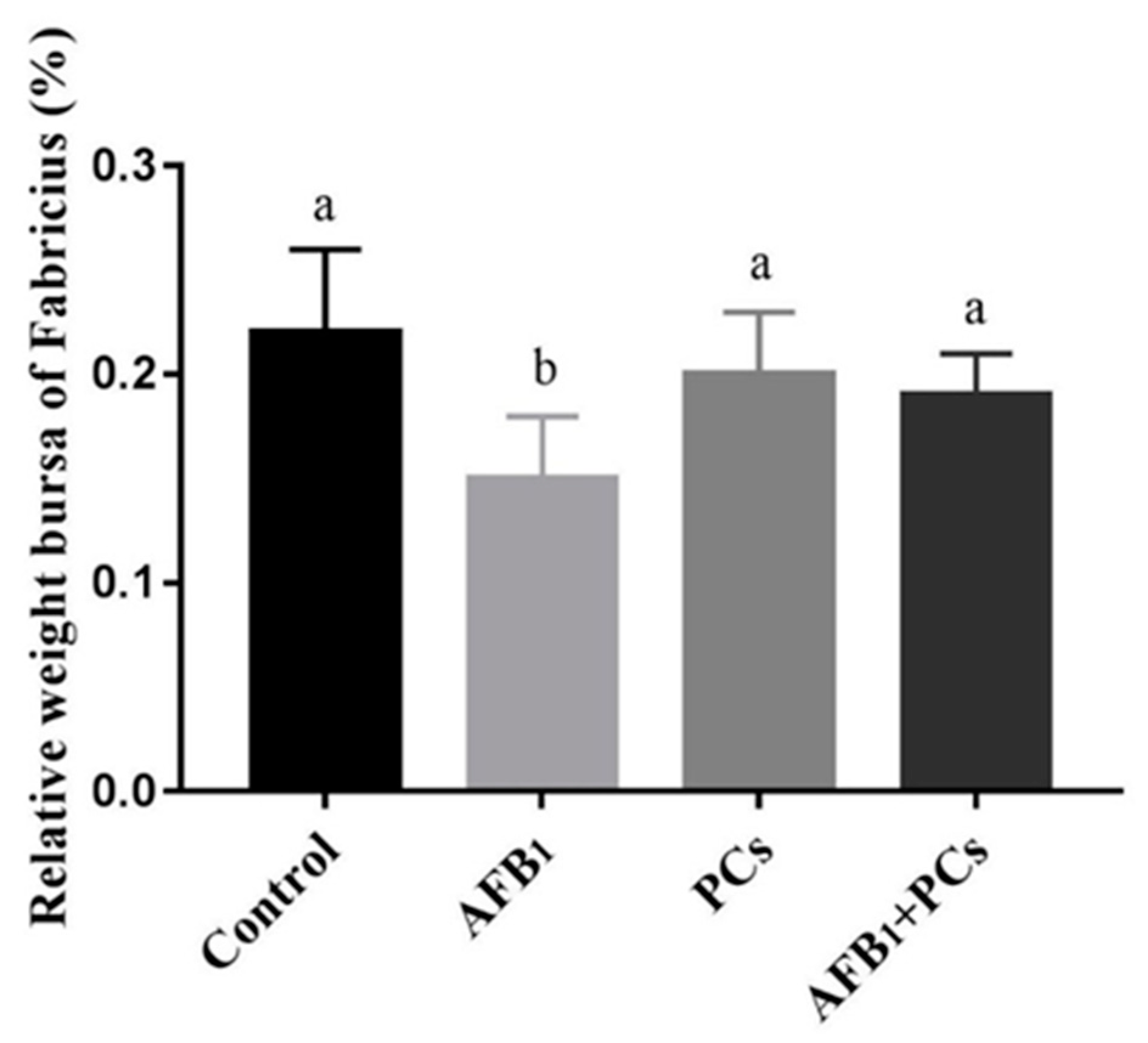
2.1. The Relative Weight of Bursa of Fabricius
2.2. Pathological Observation
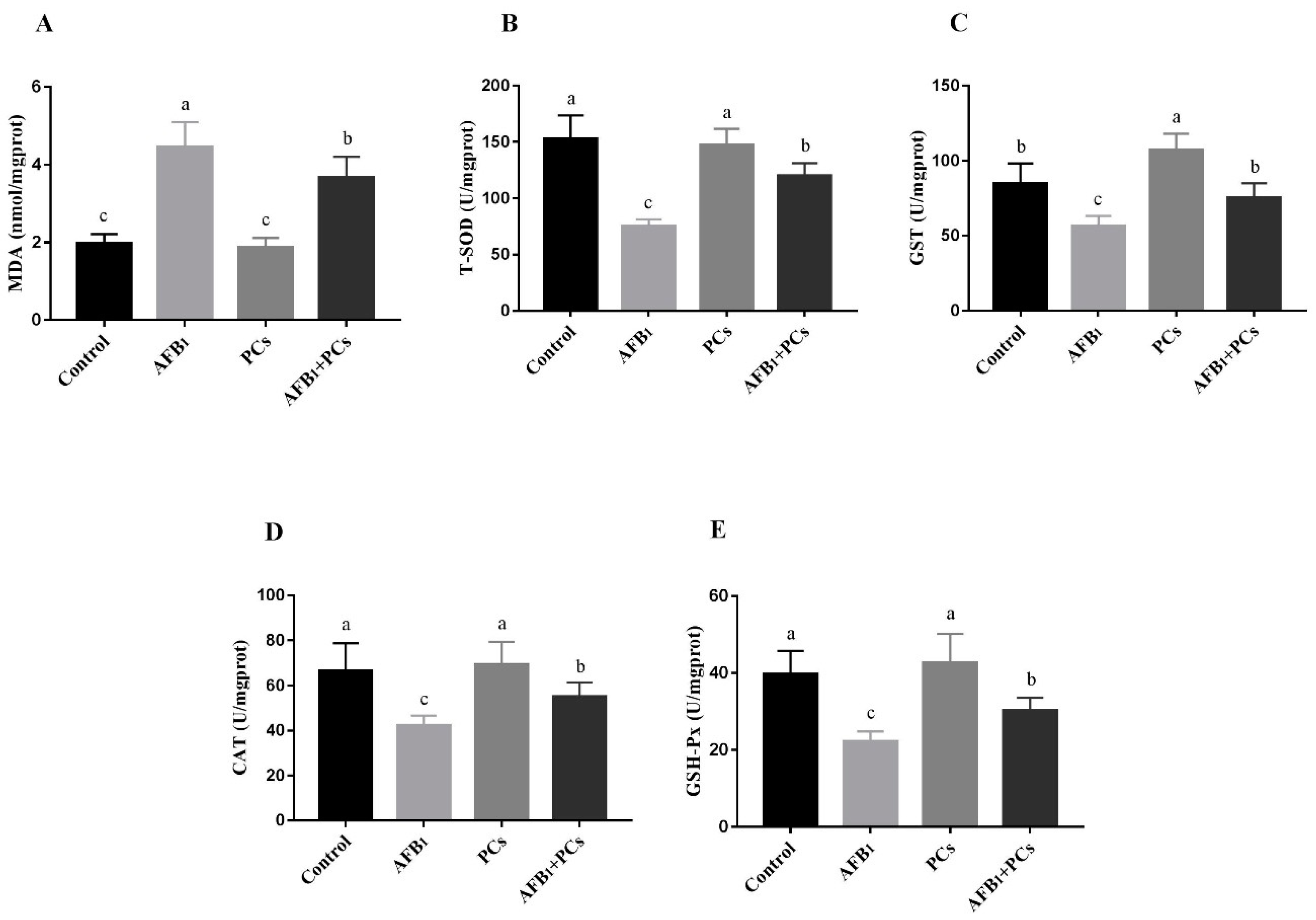
2.3. Determination of Oxidative Stress Markers
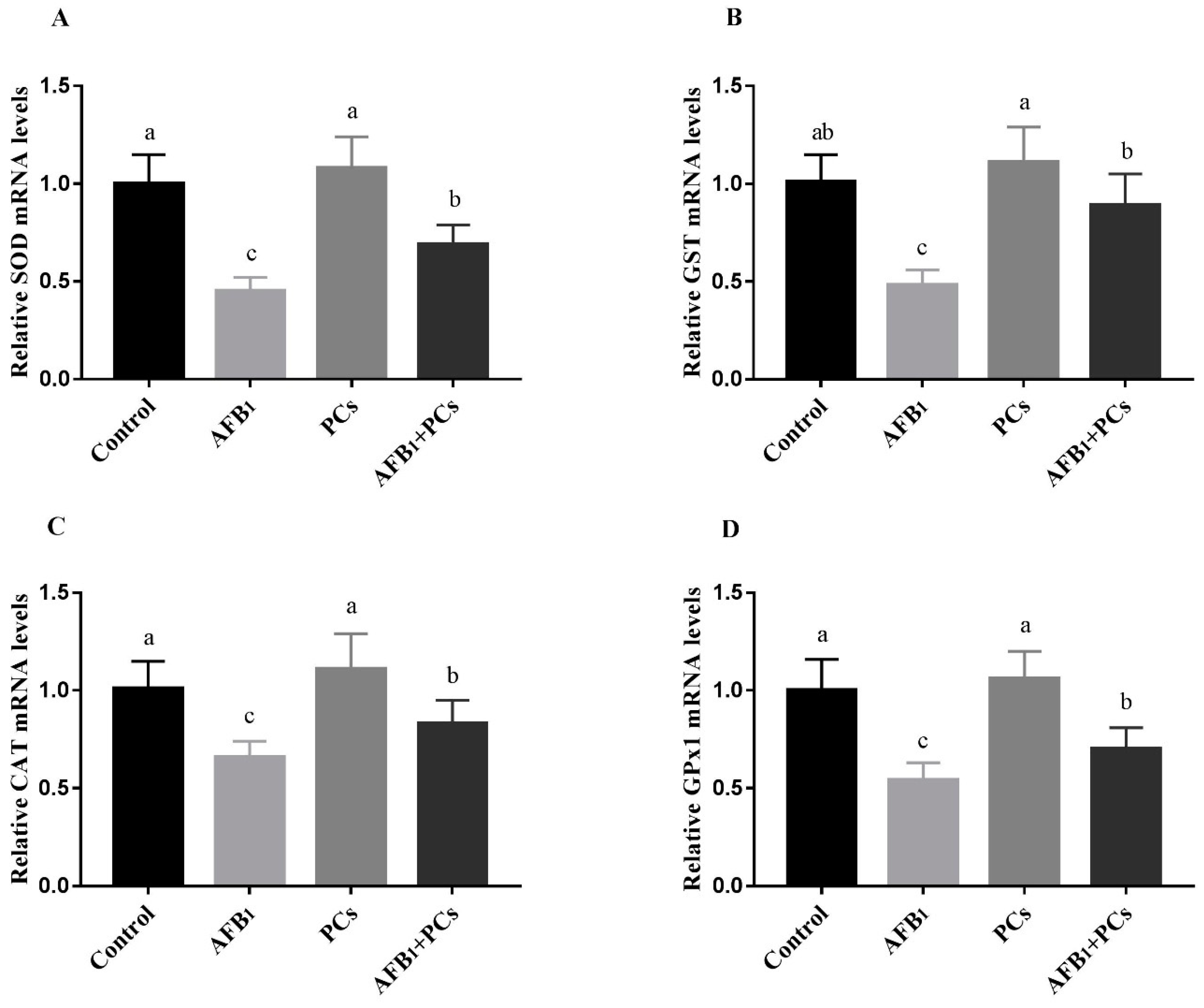
2.4. Relative mRNA Expression of Antioxidant-Related Genes
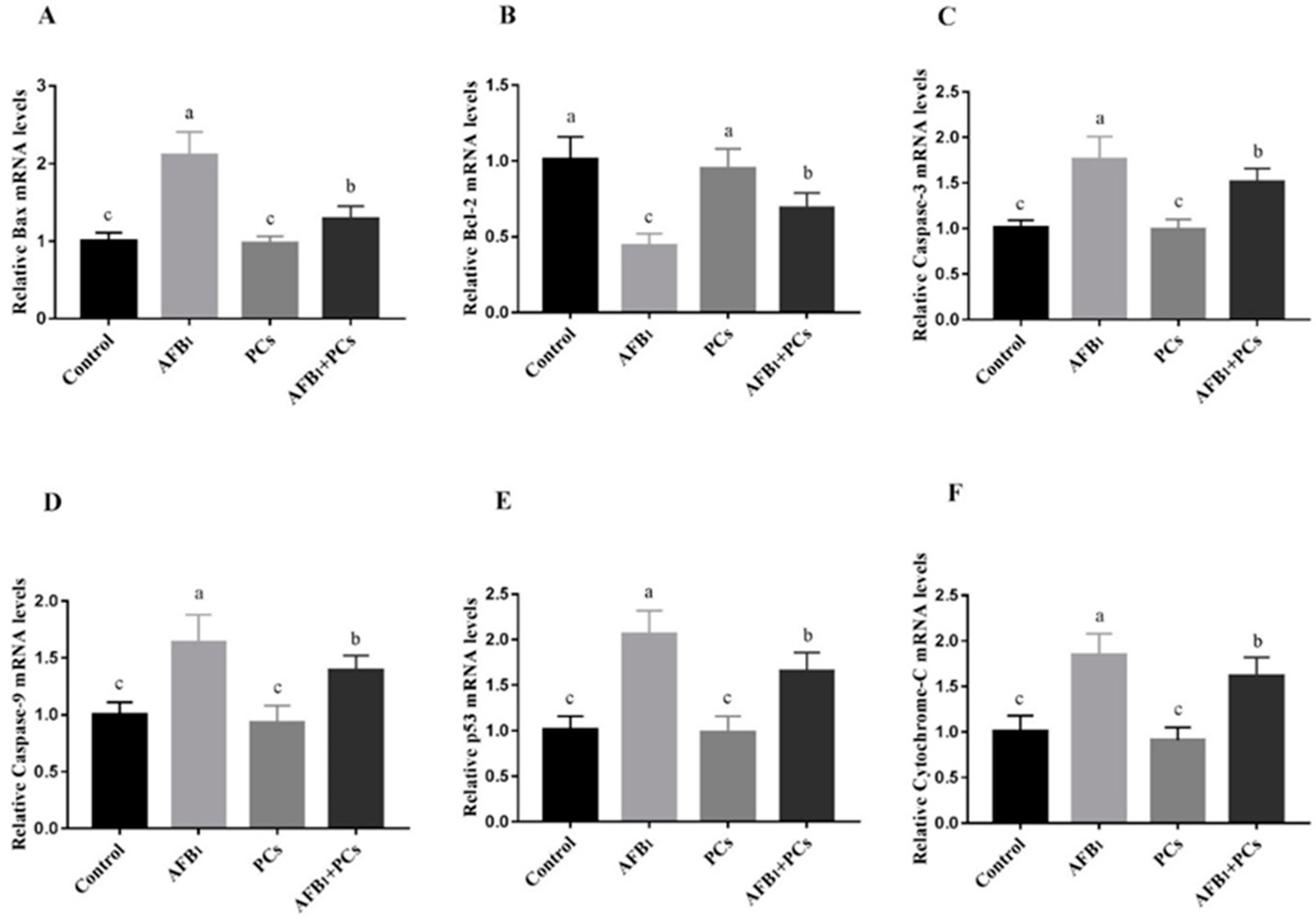
2.5. Relative mRNA Expression Levels of Apoptosis-Associated Genes
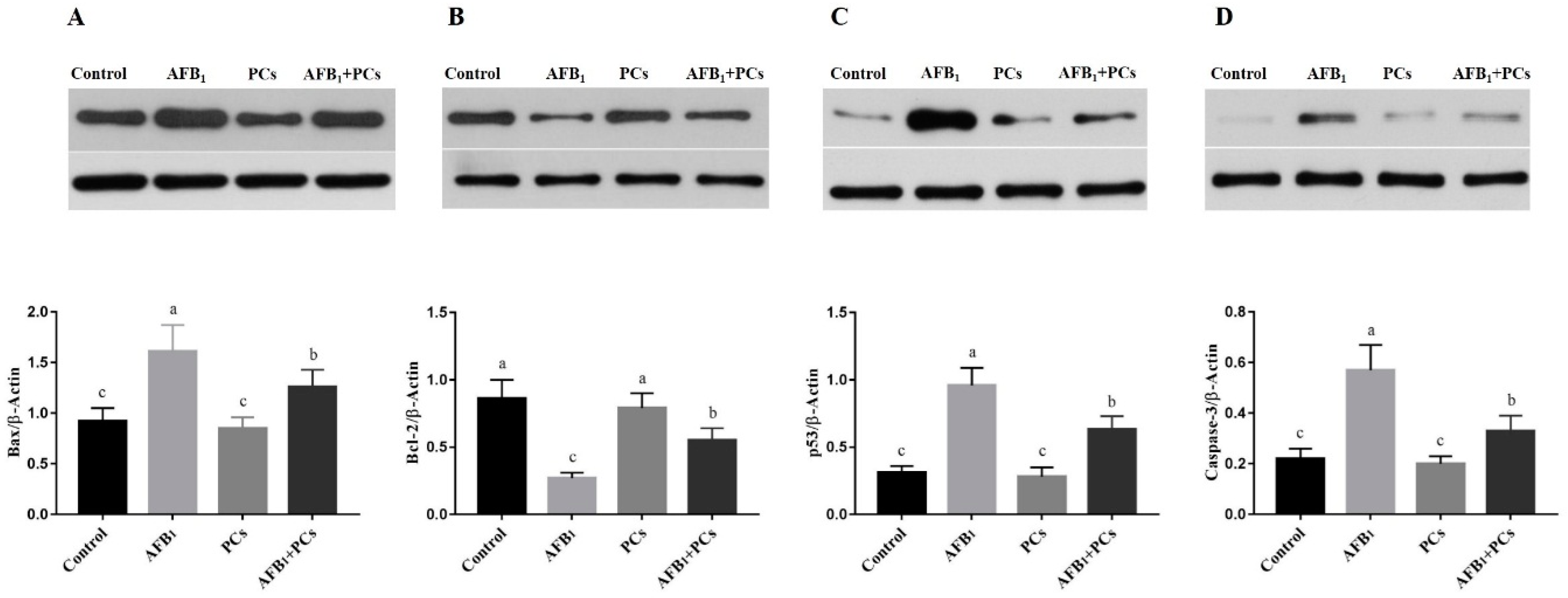
2.6. Protein Expression Levels of Apoptosis-Associated Genes
3. Discussion
4. Conclusions
5. Material and Methods
5.1. Animal Ethics
5.2. Aflatoxin B1 Production and Analysis
5.3. Bird, Diets, and Management
5.4. Collection of Samples
5.5. Pathological Observation
5.6. Determination of Oxidative Stress
5.7. Total RNA Extraction and Quantitative Real-Time PCR
5.8. Western Blot Analysis
5.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Marroquin-Cardona, A.G.; Johnson, N.M.; Phillips, T.D.; Hayes, A.W. Mycotoxins in a changing global environment--a review. Food Chem. Toxicol. 2014, 69, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Mary, V.S.; Valdehita, A.; Navas, J.M.; Rubinstein, H.R.; Fernandezcruz, M.L. Effects of aflatoxin B1, fumonisin B1 and their mixture on the aryl hydrocarbon receptor and cytochrome P450 1A induction. Food Chem. Toxicol. 2015, 75, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.W. IARC-Monographs on the Evaluation of the Carcinogenic Risk of Chemicals to Humans; International Agency for Research on Cancer: Lyon, France, 1986. [Google Scholar]
- Trebak, F.; Alaoui, A.; Alexandre, D.; El Ouezzani, S.; Anouar, Y.; Chartrel, N.; Magoul, R. Impact of aflatoxin B1 on hypothalamic neuropeptides regulating feeding behavior. Neurotoxicology 2015, 49, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.Y.; Qi, M.; Zhao, L.; Zhu, M.K.; Guo, J.; Liu, J.; Gu, C.Q.; Rajput, S.A.; Krumm, C.S.; Qi, D.S. Curcumin prevents aflatoxin B₁ hepatoxicity by inhibition of cytochrome p450 isozymes in chick liver. Toxins (Basel) 2016, 8, 327. [Google Scholar] [CrossRef] [PubMed]
- Stettler, P.M.; Sengstag, C. Liver carcinogen aflatoxin B1 as an inducer of mitotic recombination in a human cell line. Mol. Carcinog. 2001, 31, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Meissonnier, G.M.; Pinton, P.; Laffitte, J.; Cossalter, A.M.; Gong, Y.Y.; Wild, C.P.; Bertin, G.; Galtier, P.; Oswald, I.P. Immunotoxicity of aflatoxin B1: Impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol. Appl. Pharmacol. 2008, 231, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahhab, A.W.; Aly, S.E. Antioxidants and radical scavenging properties of vegetable extracts in rats fed aflatoxin-contaminated diet. J. Agric. Food Chem. 2003, 51, 2409–2414. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F. Natural Antioxidants in Avian Nutrition and Reproduction, 1st ed.; Nottingham University Press: Nottingham, UK, 2002. [Google Scholar]
- Shen, H.M.; Shi, C.Y.; Lee, H.P.; Ong, C.N. Aflatoxin B1-induced lipid peroxidation in rat liver. Toxicol. Appl. Pharmacol. 1994, 127, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Samali, A.; Fulda, S.; Gorman, A.M.; Hori, O.; Srinivasula, S.M. Cellular stress responses: Cell survival and cell death. Int. J. Cell Biol. 2010, 2010, 245803. [Google Scholar] [CrossRef] [PubMed]
- Brahmi, D.; Bouaziz, C.; Ayed, Y.; Mansour, H.B.; Zourgui, L.; Bacha, H. Chemopreventive effect of cactus Opuntia ficus indicaon oxidative stress and genotoxicity of aflatoxin B1. Nutr. Metab. 2011, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Bbosa, G.S.; Kitya, D.; Odda, J.; Ogwal-Okeng, J. Aflatoxins metabolism, effects on epigenetic mechanisms and their role in carcinogenesis. Health (N. Y.) 2013, 5, 14–34. [Google Scholar] [CrossRef]
- Chen, K.; Fang, J.; Peng, X.; Cui, H.; Chen, J.; Wang, F.; Chen, Z.; Zuo, Z.; Deng, J.; Lai, W.; Zhou, Y. Effect of selenium supplementation on aflatoxin B1-induced histopathological lesions and apoptosis in bursa of Fabricius in broilers. Food Chem. Toxicol. 2014, 74, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Shu, G.; Peng, X.; Fang, J.; Chen, K.; Cui, H.; Chen, Z.; Zuo, Z.; Deng, J.; Geng, Y.; Lai, W. Protective effects of sodium selenite against aflatoxin B1-induced oxidative stress and apoptosis in broiler spleen. Int. J. Environ. Res. Public Health 2013, 10, 2834–2844. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Ma, Q.; Zhao, L.; Jia, R.; Zhang, J.; Ji, C.; Wang, X. Protective effects of sporoderm-broken spores of ganderma lucidum on growth performance, antioxidant capacity and immune function of broiler chickens exposed to low level of aflatoxin B1. Toxins (Basel) 2016, 8, 278. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Zhang, K.; Bai, S.; Ding, X.; Zeng, Q.; Yang, J.; Fang, J.; Chen, K. Histological lesions, cell cycle arrest, apoptosis and T cell subsets changes of spleen in chicken fed aflatoxin-contaminated corn. Int. J. Environ. Res. Public Health 2014, 11, 8567–8580. [Google Scholar] [CrossRef] [PubMed]
- Ariga, T. The antioxidative function, preventive action on disease and utilization of proanthocyanidins. Biofactors 2004, 21, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Chen, X.; Wang, N.; Wang, M.; Pan, J.; Tong, J.; Li, P.; Yang, S.; He, J. Proanthocyanidins protect epithelial cells from zearalenone-induced apoptosis via inhibition of endoplasmic reticulum stress-induced apoptosis pathways in mouse small intestines. Molecules 2018, 23, 1508. [Google Scholar] [CrossRef] [PubMed]
- Uchino, R.; Madhyastha, H.; Dhungana, S.; Nakajima, Y.; Omura, S.; Maruyama, M. NFkappaB-dependent regulation of urokinase plasminogen activator by proanthocyanidin-rich grape seed extract: Effect on invasion by prostate cancer cells. Blood Coagul. Fibrinolysis 2010, 21, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Attia, S.M.; Bakheet, S.A.; Al-Easheed, N.M. Proanthocyanidins produce significant attenuation of doxorubicin-induced mutagenicity via suppression of oxidative stress. Oxid. Med. Cell. Longev. 2010, 3, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Kim, H.J.; Jeong, I.; Hong, Y.J.; Kim, M.J.; Rhie, D.J.; Jo, Y.H.; Hahn, S.J.; Yoon, S.H. Grape seed proanthocyanidin extract inhibits glutamate-induced cell death through inhibition of calcium signals and nitric oxide formation in cultured rat hippocampal neurons. BMC Neurosci. 2011, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Sen, C.K.; Ray, S.D.; Das, D.K.; Bagchi, M.; Preuss, H.G.; Vinson, J.A. Molecular mechanisms of cardioprotection by a novel grape seed proanthocyanidin extract. Mutat. Res. 2003, 523–524, 87–97. [Google Scholar] [CrossRef]
- Potworowski, E.F. T and B lymphocytes. Organ and age distribution in the chicken. Immunology 1972, 23, 199–204. [Google Scholar] [PubMed]
- Serra, A.; Macia, A.; Romero, M.; Angles, N.; Morello, J.R.; Motilva, M.J. Distribution of procyanidins and their metabolites in rat plasma and tissues after an acute intake of hazelnut extract. Food Funct. 2011, 2, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Masteller, E.L.; Lee, K.P.; Carlson, L.M.; Thompson, C.B. Expression of sialyl Lewis (x) and Lewis (x) defines distinct stages of chicken B cell maturation. J. Immunol. 1995, 155, 5550–5556. [Google Scholar] [PubMed]
- Santin, E.; Paulillo, A.C.; Maiorka, A.; Nakaghi, L.; Macari, M.; Fisher da Silva, A.V.; Alessi, A.C. Evaluation of the efficacy of saccharomyces cerevisiae cell wall to ameliorate the toxic effects of aflatoxin in broilers. Int. J. Poult. Sci. 2003, 2, 341–344. [Google Scholar]
- Kujoth, G.C.; Hiona, A.; Pugh, T.D.; Someya, S.; Panzer, K.; Wohlgemuth, S.E.; Hofer, T.; Seo, A.Y.; Sullivan, R.; Jobling, W.A. Mitochondrial DNA mutations, oxidative stress, and apoptosis in mammalian aging. Science 2005, 309, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.F.; Liu, J.; Wasser, S.; Shen, H.; Tan, C.E.L.; Ong, C.N. Inhibition of ebselen on aflatoxin B1-induced hepatocarcinogenesis in Fischer 344 rats. Carcinogenesis 2000, 21, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nut. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Yener, Z.; Celik, I.; Ilhan, F.; Bal, R. Effects of Urtica dioica L. Seed on lipid peroxidation, antioxidants and liver pathology in aflatoxin-induced tissue injury in rats. Food Chem. Toxicol. 2009, 47, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. AJME 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Liu, X.F.; Zhang, L.M.; Guan, H.N.; Zhang, Z.W.; Xu, S.W. Effects of oxidative stress on apoptosis in manganese-induced testicular toxicity in cocks. Food Chem. Toxicol. 2013, 60, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, N.; Ma, L.; Duan, Y.; Wang, J.; Zhao, X.; Wang, S.; Wang, H.; Hong, F. Oxidative injury in the mouse spleen caused by lanthanides. J. Alloy. Compd. 2010, 489, 708–713. [Google Scholar] [CrossRef]
- Rajput, S.A.; Sun, L.; Zhang, N.; Khalil, M.M.; Gao, X.; Ling, Z.; Zhu, L.; Khan, F.A.; Zhang, J.; Qi, D. Ameliorative effects of grape seed proanthocyanidin extract on growth performance, immune function, antioxidant capacity, biochemical constituents, liver histopathology and aflatoxin residues in broilers exposed to aflatoxin B1. Toxins (Basel) 2017, 9, 371. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Zhao, L.; Ji, C.; Li, X.; Jia, R.; Xi, L.; Zhang, J.; Ma, Q. Protective effects of Bacillus subtilis ANSB060 on serum biochemistry, histopathological changes and antioxidant enzyme activities of broilers fed moldy peanut meal naturally contaminated with aflatoxins. Toxins (Basel) 2015, 7, 3330–3343. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Ling, Z.; Ni-Ya, Z.; Karrow, N.A.; Christopher, S.K.; De-Sheng, Q.; Lv-Hui, S. Aflatoxin B1 metabolism: Regulation by phase I and II metabolizing enzymes and chemoprotective agents. Mutat. Res.-/Rev. Mutat. 2018, 778, 778–779. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.A.; Ribeiro, J.M. Apoptosis and disease. The Lancet 1993, 341, 1251–1254. [Google Scholar] [CrossRef]
- Antonsson, B. Mitochondria and the Bcl-2 family proteins in apoptosis signaling pathways. Mol. Cell. Biochem. 2004, 256–257, 141–155. [Google Scholar] [CrossRef]
- Siddiqui, W.A.; Ahad, A.; Ahsan, H. The mystery of Bcl2 family: Bcl-2 proteins and apoptosis: An update. Arch. Toxicol. 2015, 89, 289–317. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, T.; Reed, J.C. Tumor suppressor p53 is a direct transcriptional activator of the human bax gene. Cell 1995, 80, 293–300. [Google Scholar] [PubMed]
- Jin, Y.; Zhang, S.; Tao, R.; Huang, J.; He, X.; Qu, L.; Fu, Z. Oral exposure of mice to cadmium (II), chromium (VI) and their mixture induce oxidative-and endoplasmic reticulum-stress mediated apoptosis in the livers. Environ. Toxicol. 2016, 31, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (ΔΨm) in apoptosis; an update. Apoptosis 2003, 8, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Reed, J.C. Mitochondria and apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Muhammad, I.; Sun, X.; Han, M.; Hamid, S.; Zhang, X. Protective role of curcumin in ameliorating AFB1-induced apoptosis via mitochondrial pathway in liver cells. Mol. Biol. Rep. 2018, 45, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Chen, K.; Chen, J.; Fang, J.; Cui, H.; Zuo, Z.; Deng, J.; Chen, Z.; Geng, Y.; Lai, W. Aflatoxin B1 affects apoptosis and expression of Bax, Bcl-2, and Caspase-3 in thymus and bursa of Fabricius in broiler chickens. Environ. Toxicol. 2016, 31, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, L.; Zhang, N.; Zhang, J.; Guo, J.; Li, C.; Rajput, S.A.; Qi, D. Effects of nutrients in substrates of different grains on aflatoxin B1 production by Aspergillus flavus. Biomed. Res. Int. 2016, 2016. [Google Scholar]
- Rajput, S.A.; Sun, L.; Zhang, N.-Y.; Khalil, M.M.; Ling, Z.; Chong, L.; Wang, S.; Rajput, I.R.; Bloch, D.M.; Khan, F.A. Grape seed proanthocyanidin extract alleviates aflatoxin B1-induced immunotoxicity and oxidative stress via modulation of NF-κBand Nrf2 signaling pathways in broilers. Toxins (Basel) 2019, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, J.; Zhang, B.; Wu, K.; Yang, A.; Li, C.; Zhang, J.; Zhang, C.; Rajput, S.A.; Zhang, N. Deoxynivalenol impairs porcine intestinal host defense peptide expression in weaned piglets and IPEC-J2 cells. Toxins (Basel) 2018, 10, 541. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.H.; Lei, M.Y.; Zhang, N.Y.; Gao, X.; Li, C.; Krumm, C.S.; Qi, D.S. Individual and combined cytotoxic effects of aflatoxin B1, zearalenone, deoxynivalenol and fumonisin B1 on BRL 3A rat liver cells. Toxicon 2015, 95, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Xiao, Z.; Li, C.; Zhang, J.; Zhu, L.; Sun, L.; Zhang, N.; Khalil, M.M.; Rajput, S.A.; Qi, D. Prenatal exposure to zearalenone disrupts reproductive potential and development via hormone-related genes in male rats. Food Chem. Toxicol. 2018, 116, 11–19. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | (%) |
|---|---|
| Corn | 58.3 |
| Soybean meal | 30.2 |
| Fish meal | 5.6 |
| Soybean oil | 2.3 |
| Dicalcium phosphate | 1.2 |
| Lime stone | 1.00 |
| Salt | 0.2 |
| Methionine | 0.2 |
| Premix 1 | 1.00 |
| Total | 100.00 |
| Calculated chemical composition | |
| Crude protein | 21.87 |
| Metabolisable energy (MJ/kg) | 13.45 |
| Lysine | 1.14 |
| Methionine | 0.40 |
| Methionine + Cystine | 0.94 |
| Calcium | 0.95 |
| Available phosphorus | 0.49 |
| Target Gene | Primer | Primer Sequence (5′→3′) | Accession No. |
|---|---|---|---|
| β-Actin | Forward Reverse | CCCGCAAATGCTCTAAACC CCAATCCTGTCTTGTTTTATGC | L08165 |
| Bax | Forward Reverse | TCCTCATCGCCATGCTCAT CCTTGGTCTGGAAGCAGAAGA | XM_422067 |
| Bcl-2 | Forward Reverse | CGCCGCTACCAGAGGGACTT CCGGACCCAGTTGACCCCAT | Z_11961.1 |
| Caspase-9 | Forward Reverse | CCAACCTGAGAGTGAGCGATT GTACACCAGTCTGTGGGTCGG | AY057940 |
| Caspase-3 | Forward Reverse | GGCTCCTGGTTTATTCAGTCTC ATTCTGCCACTCTGCGATTT | NM_204725.1 |
| p53 | Forward Reverse | GCCGTGGCCGTCTATAAGAA GGTCTCGTCGTCGTGGTAAC | NM_205264.1 |
| SOD | Forward Reverse | CGTCATTCACTTCGAGCAGAAGG GTCTGAGACTCAGACCACATA | NM_205064 |
| GPx1 | Forward Reverse | GACCAACCCGCAGTACATCA GAGGTGCGGGCTTTCCTTTA | NM_001277853.1 |
| CAT | Forward Reverse | CCACGTGGACCTCTTCTTGT AAACACTTTCGCCTTGCAGT | NM_001031215.1 |
| GST | Forward Reverse | AGTCGAAGCCTGATGCACTT TCTAGGCGTGGTTTCCTTTG | L15386.1 |
| Cytochrome-C | Forward Reverse | CGCAGGCTCCATACTACTCG TTAGGGCACCTCATAGGGCT | NC_001323.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajput, S.A.; Zhang, C.; Feng, Y.; Wei, X.T.; Khalil, M.M.; Rajput, I.R.; Baloch, D.M.; Shaukat, A.; Rajput, N.; Qamar, H.; et al. Proanthocyanidins Alleviates AflatoxinB1-Induced Oxidative Stress and Apoptosis through Mitochondrial Pathway in the Bursa of Fabricius of Broilers. Toxins 2019, 11, 157. https://doi.org/10.3390/toxins11030157
Rajput SA, Zhang C, Feng Y, Wei XT, Khalil MM, Rajput IR, Baloch DM, Shaukat A, Rajput N, Qamar H, et al. Proanthocyanidins Alleviates AflatoxinB1-Induced Oxidative Stress and Apoptosis through Mitochondrial Pathway in the Bursa of Fabricius of Broilers. Toxins. 2019; 11(3):157. https://doi.org/10.3390/toxins11030157
Chicago/Turabian StyleRajput, Shahid Ali, Cong Zhang, Yue Feng, Xiao Tian Wei, Mahmoud Mohamed Khalil, Imran Rashid Rajput, Dost Muhammad Baloch, Aftab Shaukat, Nasir Rajput, Hammad Qamar, and et al. 2019. "Proanthocyanidins Alleviates AflatoxinB1-Induced Oxidative Stress and Apoptosis through Mitochondrial Pathway in the Bursa of Fabricius of Broilers" Toxins 11, no. 3: 157. https://doi.org/10.3390/toxins11030157
APA StyleRajput, S. A., Zhang, C., Feng, Y., Wei, X. T., Khalil, M. M., Rajput, I. R., Baloch, D. M., Shaukat, A., Rajput, N., Qamar, H., Hassan, M., & Qi, D. (2019). Proanthocyanidins Alleviates AflatoxinB1-Induced Oxidative Stress and Apoptosis through Mitochondrial Pathway in the Bursa of Fabricius of Broilers. Toxins, 11(3), 157. https://doi.org/10.3390/toxins11030157