Production of Cyanotoxins by Microcystis aeruginosa Mediates Interactions with the Mixotrophic Flagellate Cryptomonas


Abstract
:1. Introduction
2. Results
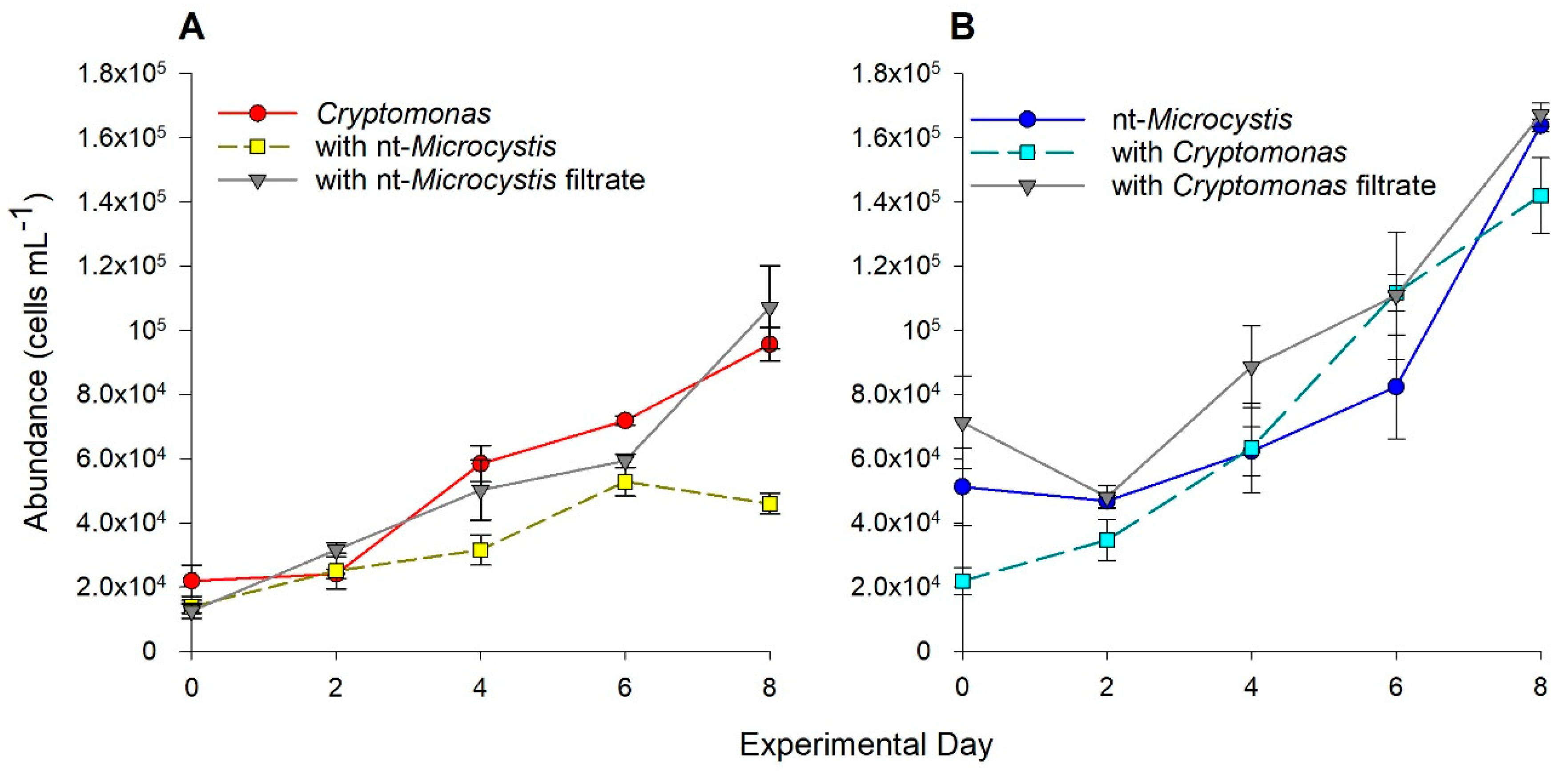
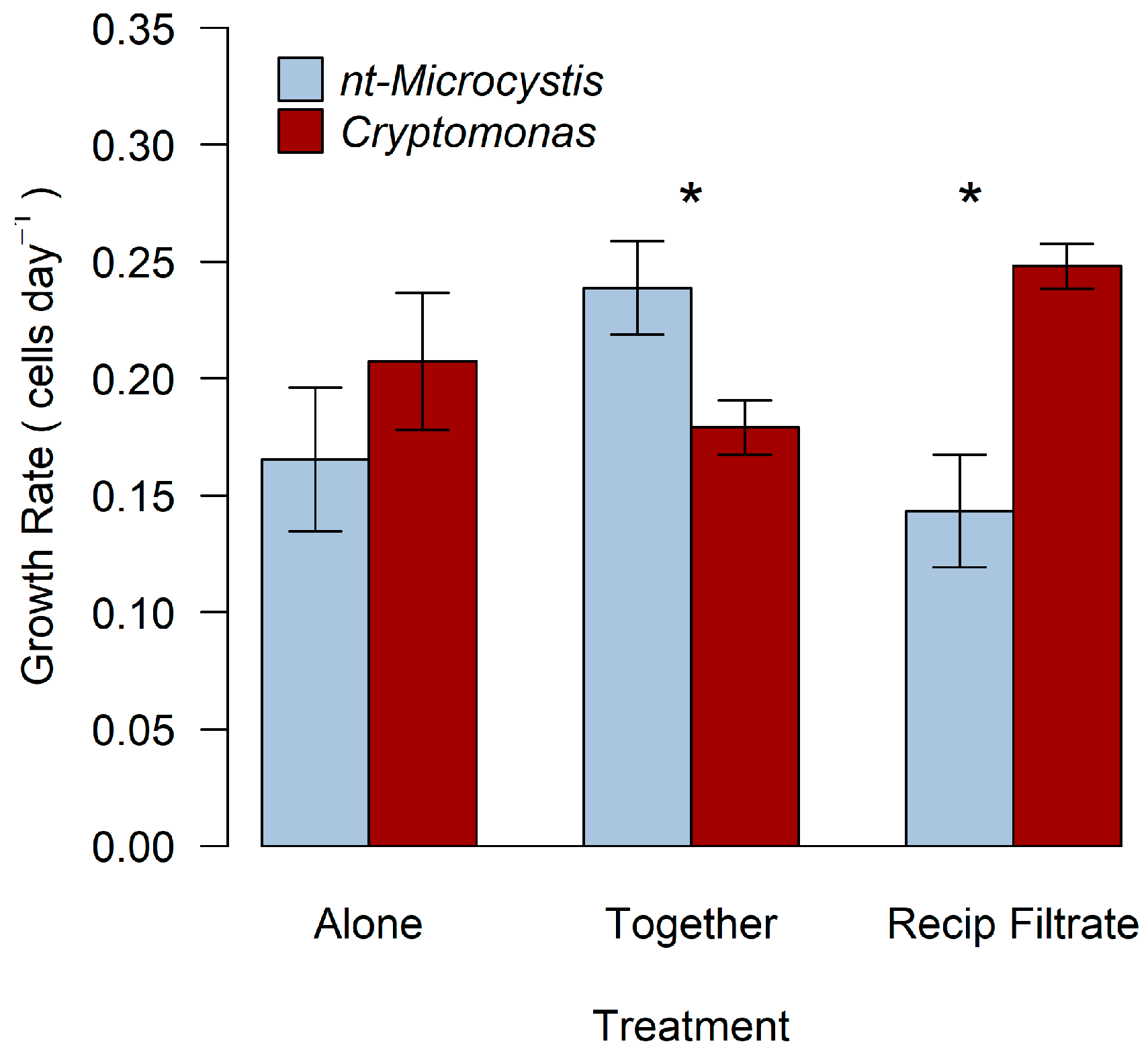
2.1. Biotic Interactions between Cryptomonas and nt-Microcystis
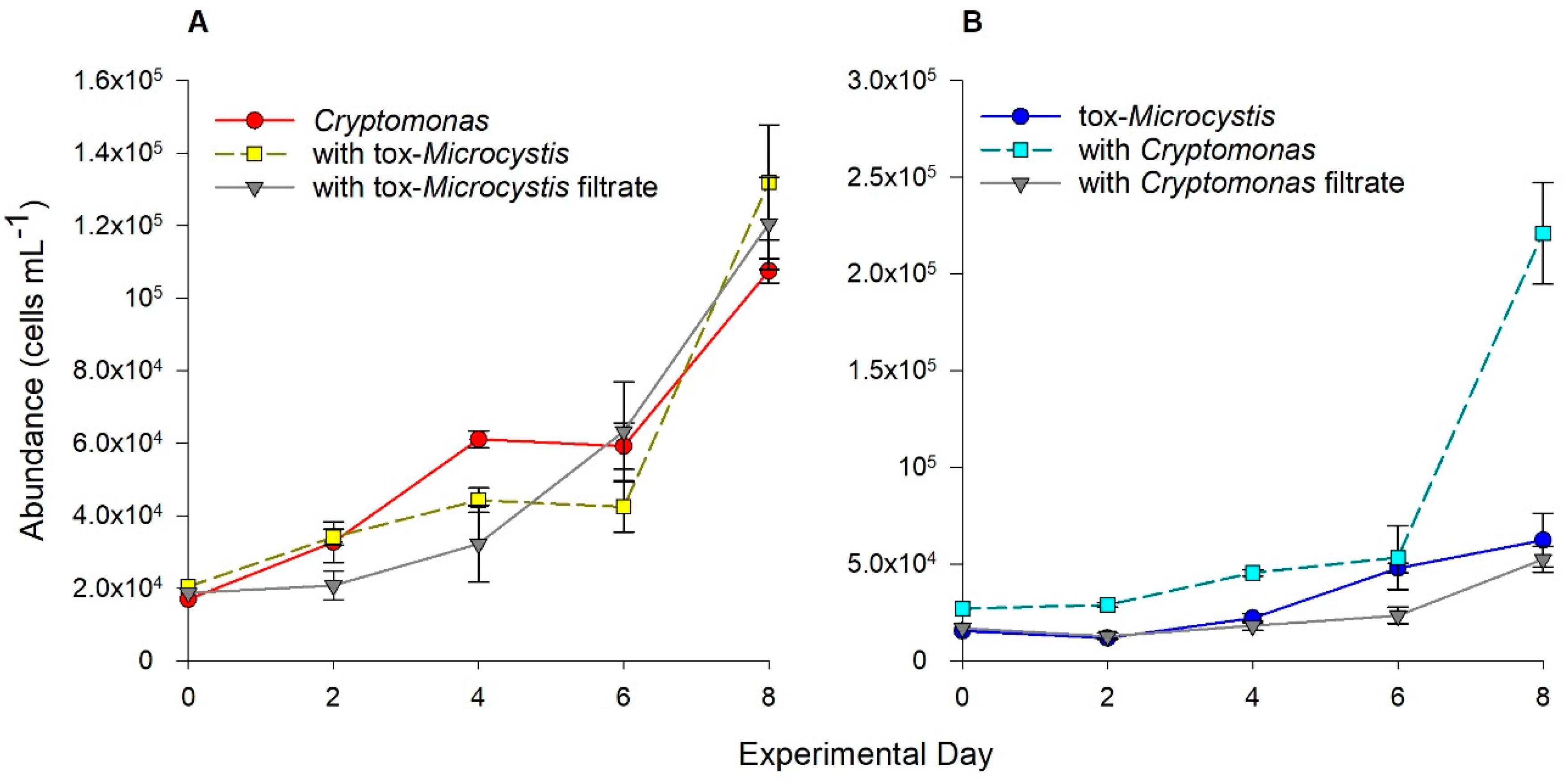
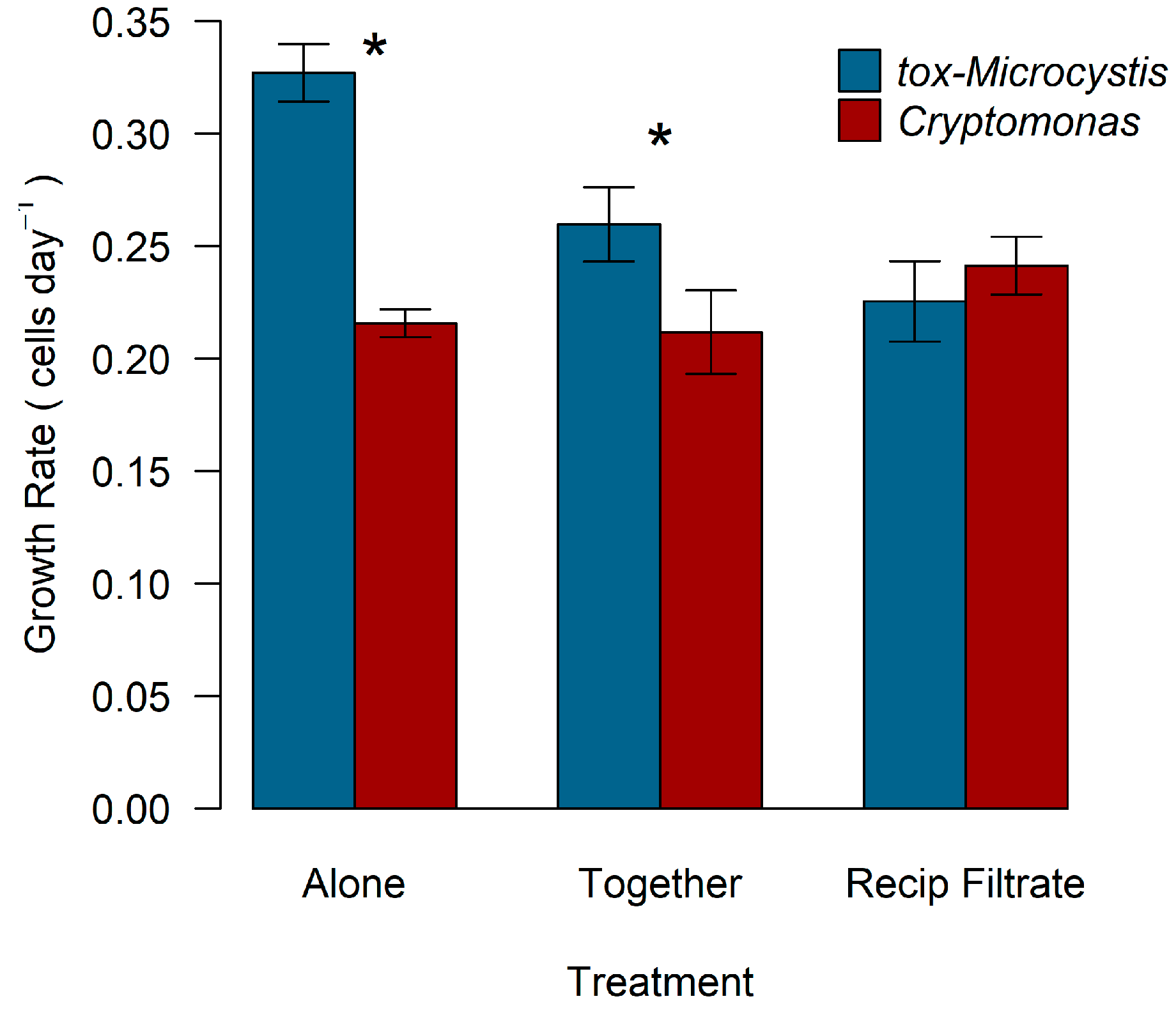
2.2. Biotic Interactions between Cryptomonas and tox-Microcystis
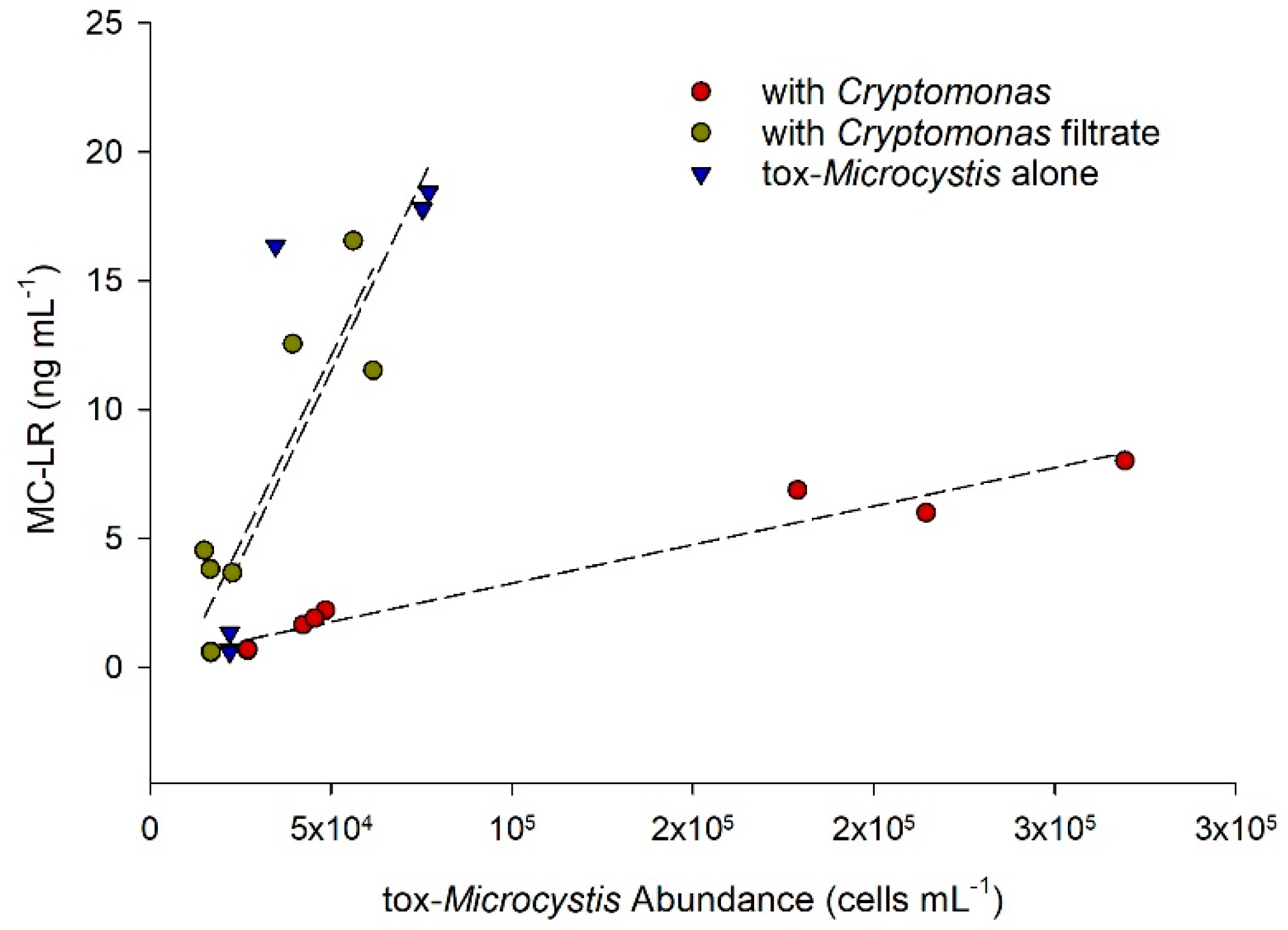
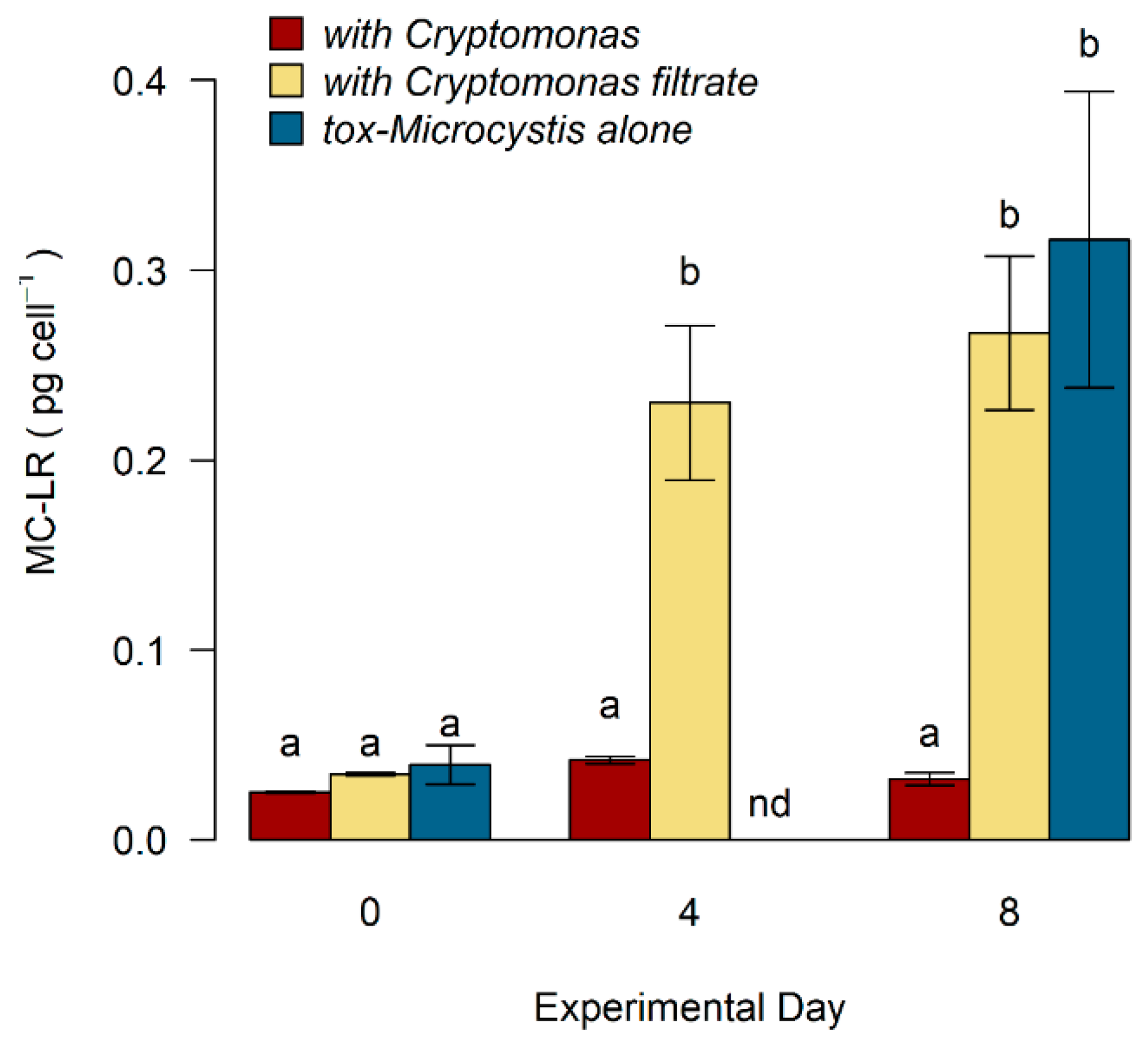
2.3. Toxicity of Microcystis in Response to Cryptomonas
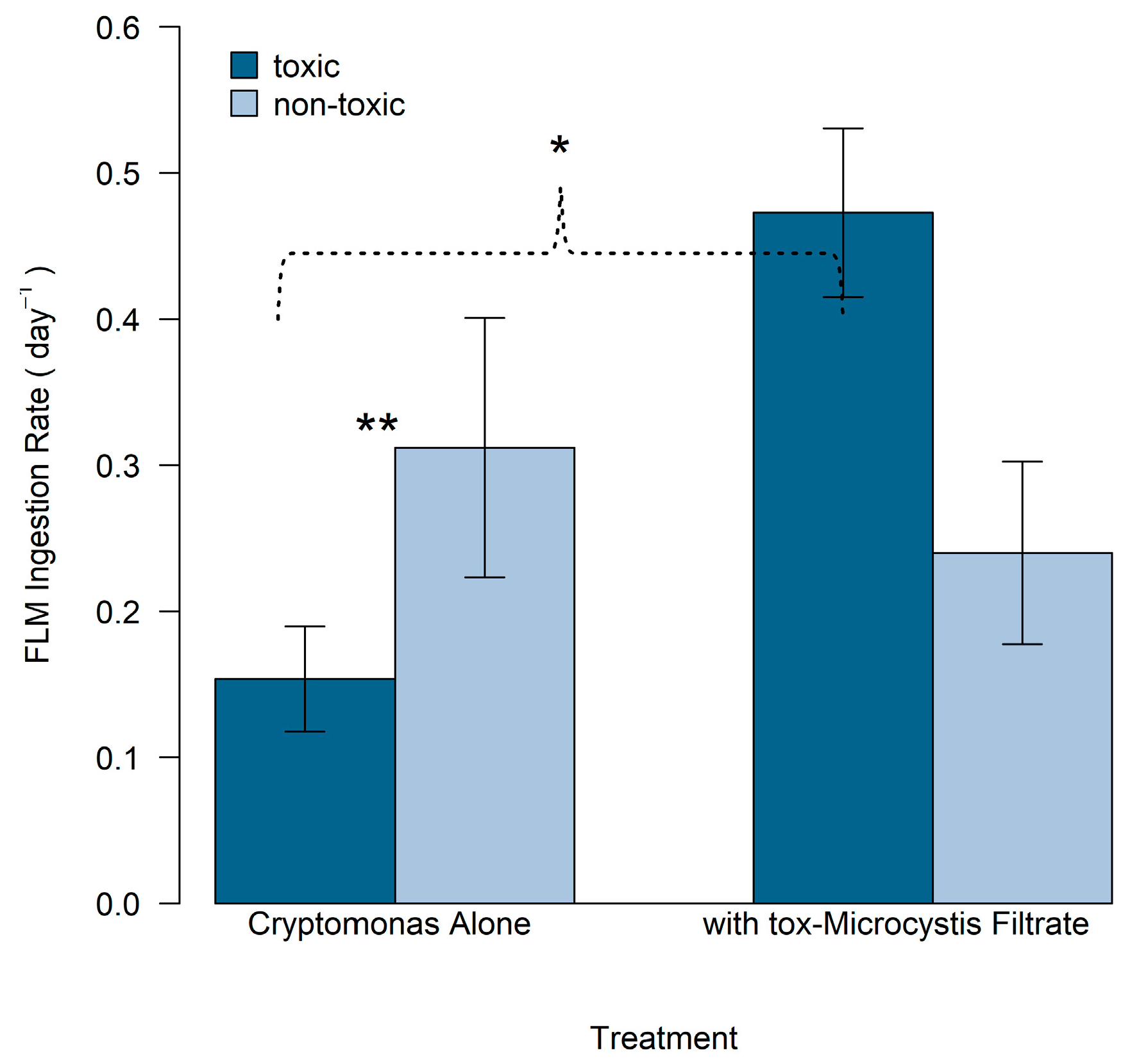
2.4. Ability of Mixotrophic Cryptomonas to Ingest tox- and nt-Microcystis
3. Discussion
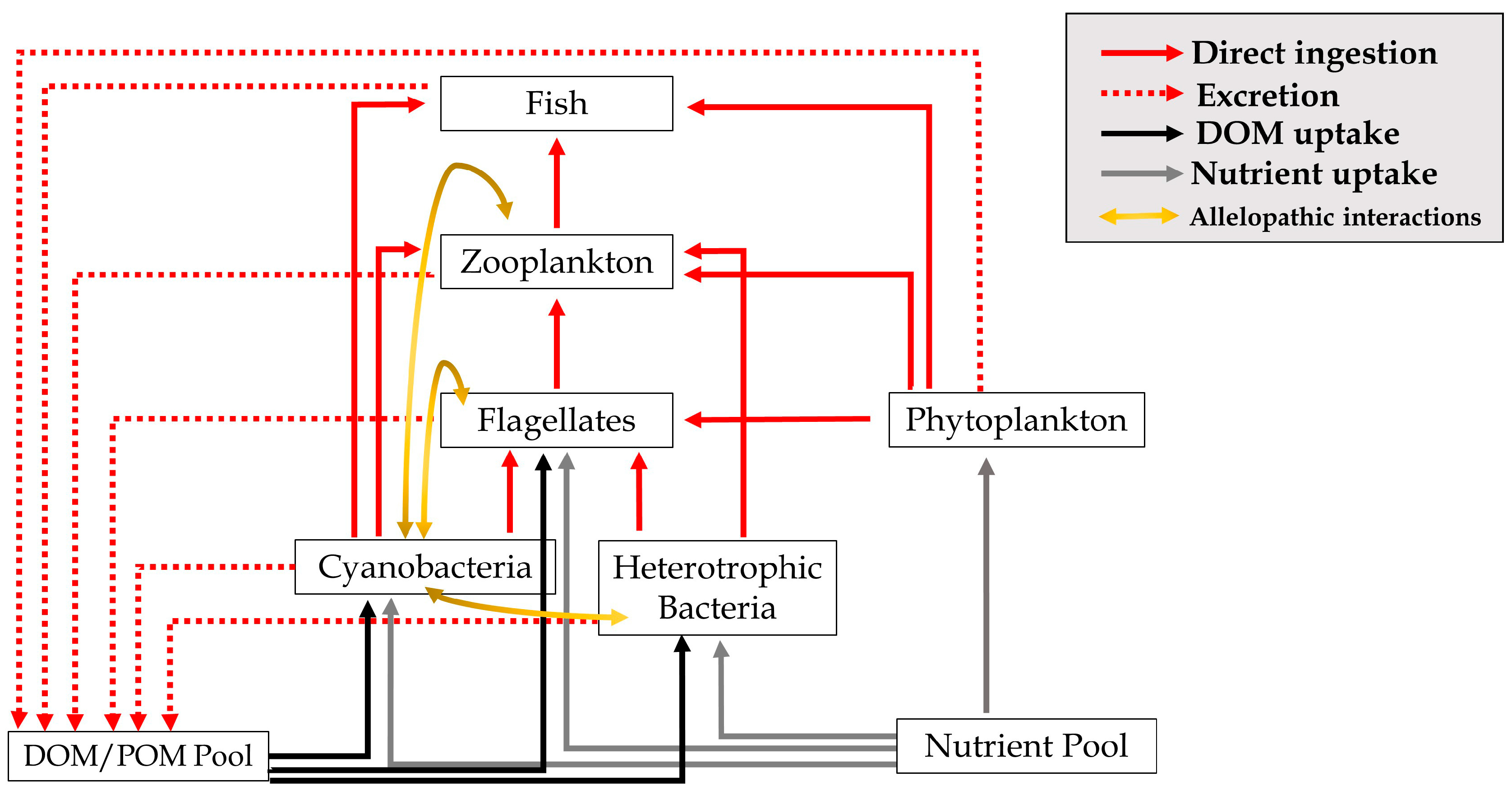
4. Materials and Methods
4.1. Preparation and Analysis of Algal Filtrate
4.2. Biotic Interactions between a Mixotrophic Flagellate, Cryptomonas, and Two Strains of Microcystis aeruginosa
4.3. Cyanobacterivory by Cryptomonas
4.4. Microcystin Analysis and Extraction
4.5. Cell Enumeration and Examination of Bacterivory by Fluorescence Microscopy
4.6. Determination of Population Growth Rate and Ingestion Rate
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beaulieu, M.; Pick, F.; Gregory-Eaves, I. Nutrients and water temperature are significant predictors of cyanobacterial biomass in a 1147 lakes data set. Limnol. Oceanogr. 2013, 58, 1736–1746. [Google Scholar] [CrossRef] [Green Version]
- Taranu, Z.E.; Gregory-Eaves, I.; Leavitt, P.R.; Bunting, L.; Buchaca, T.; Catalan, J.; Domaizon, I.; Guilizzoni, P.; Lami, A.; McGowan, S.; et al. Acceleration of cyanobacterial dominance in north temperate-subarctic lakes during the Anthropocene. Ecol. Lett. 2015, 18, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Beaver, J.R.; Manis, E.E.; Loftin, K.A.; Graham, J.L.; Pollard, A.I.; Mitchell, R.M. Land use patterns, ecoregion, and microcystin relationships in US lakes and reservoirs: A preliminary evaluation. Harmful Algae 2014, 36, 57–62. [Google Scholar] [CrossRef]
- Paerl, H.W. Mitigating Toxic Planktonic Cyanobacterial Blooms in Aquatic Ecosystems Facing Increasing Anthropogenic and Climatic Pressures. Toxins 2018, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Dolman, A.M.; Rucker, J.; Pick, F.R.; Fastner, J.; Rohrlack, T.; Mischke, U.; Wiedner, C. Cyanobacteria and cyanotoxins: The influence of nitrogen versus phosphorus. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Rigosi, A.; Carey, C.C.; Ibelings, B.W.; Brookes, J.D. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol. Oceanogr. 2014, 59, 99–114. [Google Scholar] [CrossRef] [Green Version]
- Marmen, S.; Aharonovich, D.; Grossowicz, M.; Blank, L.; Yacobi, Y.Z.; Sher, D.J. Distribution and Habitat Specificity of Potentially-Toxic Microcystis across Climate, Land, and Water Use Gradients. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Beaver, J.R.; Tausz, C.E.; Scotese, K.C.; Pollard, A.I.; Mitchell, R.M. Environmental factors influencing the quantitative distribution of microcystin and common potentially toxigenic cyanobacteria in US lakes and reservoirs. Harmful Algae 2018, 78, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Adrian, R. Cyanobacteria dominance: Quantifying the effects of climate change. Limnol. Oceanogr. 2009, 54, 2460–2468. [Google Scholar] [CrossRef] [Green Version]
- Carey, C.C.; Ibelings, B.W.; Hoffmann, E.P.; Hamilton, D.P.; Brookes, J.D. Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate. Water Res. 2012, 46, 1394–1407. [Google Scholar] [CrossRef]
- Wilkinson, A.A.; Hondzo, M.; Guala, M. Investigating abiotic drivers for vertical and temporal heterogeneities of cyanobacteria concentrations in lakes using a seasonal in-situ monitoring station. Water Resour. Res. 2018. [Google Scholar] [CrossRef]
- Graham, J.L.; Loftin, K.A.; Meyer, M.T.; Ziegler, A.C. Cyanotoxin Mixtures and Taste-and-Odor Compounds in Cyanobacterial Blooms from the Midwestern United States. Environ. Sci. Technol. 2010, 44, 7361–7368. [Google Scholar] [CrossRef] [PubMed]
- Loftin, K.A.; Graham, J.L.; Hilborn, E.D.; Lehmann, S.C.; Meyer, M.T.; Dietze, J.E.; Griffith, C.B. Cyanotoxins in inland lakes of the United States: Occurrence and potential recreational health risks in the EPA Natural Lakes Assessment 2007. Harmful Algae 2016, 56, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, K. Ecological implications of cyanobacterial toxins in aquatic food webs. Phycologia 1996, 35, 42–50. [Google Scholar] [CrossRef]
- Jacoby, J.M.; Collier, D.C.; Welch, E.B.; Hardy, F.J.; Crayton, M. Environmental factors associated with a toxic bloom of Microcystis aeruginosa. Can. J. Fish. Aquat. Sci. 2000, 57, 231–240. [Google Scholar] [CrossRef]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8. [Google Scholar] [CrossRef]
- Hayes, N.M.; Vanni, M.J. Microcystin concentrations can be predicted with phytoplankton biomass and watershed morphology. Inland Waters 2018. [Google Scholar] [CrossRef]
- Wilson, A.E.; Sarnelle, O.; Tillmanns, A.R. Effects of cyanobacterial toxicity and morphology on the population growth of freshwater zooplankton: Meta-analyses of laboratory experiments. Limnol. Oceanogr. 2006, 51, 1915–1924. [Google Scholar] [CrossRef] [Green Version]
- Urrutia-Cordero, P.; Ekvall, M.K.; Hansson, L.A. Responses of cyanobacteria to herbivorous zooplankton across predator regimes: Who mows the bloom? Freshw. Biol. 2015, 60, 960–972. [Google Scholar] [CrossRef]
- Ger, K.A.; Otten, T.G.; DuMais, R.; Ignoffo, T.; Kimmerer, W. In situ ingestion of Microcystis is negatively related to copepod abundance in the upper San Francisco Estuary. Limnol. Oceanogr. 2018, 63, 2394–2410. [Google Scholar] [CrossRef]
- Triest, L.; Stiers, I.; Van Onsem, S. Biomanipulation as a nature-based solution to reduce cyanobacterial blooms. Aquat. Ecol. 2016, 50, 461–483. [Google Scholar] [CrossRef]
- Bierman, V.J.; Kaur, J.; DePinto, J.V.; Feist, T.J.; Dilks, D.W. Modeling the role of zebra mussels in the proliferation of blue-green algae in Saginaw Bay, Lake Huron. J. Great Lakes Res. 2005, 31, 32–55. [Google Scholar] [CrossRef]
- Fulton, R.S.; Paerl, H.W. effects of colonial morphology on zooplankton utilization of algal resources during blue-green-algal (microcystis-aeruginosa) blooms. Limnol. Oceanogr. 1987, 32, 634–644. [Google Scholar] [CrossRef]
- Demott, W.R.; Moxter, F. Foraging on cyanobacteria by copepods—Responses to chemical defenses and resource abundance. Ecology 1991, 72, 1820–1834. [Google Scholar] [CrossRef]
- Yang, Z.; Kong, F.X.; Shi, X.L.; Zhang, M.; Xing, P.; Cao, H.S. Changes in the morphology and polysaccharide content of Microcystis aeruginosa (Cyanobacteria) during flagellate grazing. J. Phycol. 2008, 44, 716–720. [Google Scholar] [CrossRef]
- Yang, Z.; Kong, F.X. Formation of large colonies: A defense mechanism of Microcystis aeruginosa under continuous grazing pressure by flagellate Ochromonas sp. J. Limnol. 2012, 71, 61–66. [Google Scholar] [CrossRef]
- Kim, B.R.; Nakano, S.; Kim, B.H.; Han, M.S. Grazing and growth of the heterotrophic flagellate Diphylleia rotans on the cyanobacterium Microcystis aeruginosa. Aquat. Microb. Ecol. 2006, 45, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, Z.; Alshehri, A. Grazing on Microcystis aeruginosa and degradation of microcystins by the heterotrophic flagellate Diphylleia rotans. Ecotox. Environ. Saf. 2013, 96, 48–52. [Google Scholar] [CrossRef]
- Van Wichelen, J.; van Gremberghe, I.; Vanormelingen, P.; Debeer, A.E.; Leporcq, B.; Menzel, D.; Codd, G.A.; Descy, J.P.; Vyverman, W. Strong effects of amoebae grazing on the biomass and genetic structure of a Microcystis bloom (Cyanobacteria). Environ. Microbiol. 2010, 12, 2797–2813. [Google Scholar] [CrossRef]
- Caron, D.A.; Countway, P.D.; Jones, A.C.; Kim, D.Y.; Schnetzer, A. Marine protistan diversity. Ann. Rev. Mar. Sci. 2012, 4, 467–493. [Google Scholar] [CrossRef]
- Domaizon, I.; Viboud, S.; Fontvieille, D. Taxon-specific and seasonal variation in flagellates grazing on heterotrophic bacteria in the oligotrophic Lake Annecy – importance of mixotrophy. FEMS Microbiol. Ecol. 2003, 46, 317–329. [Google Scholar] [CrossRef]
- Oikonomou, A.; Filker, S.; Breiner, H.; Stoeck, T. Protistan diversity in a permanently stratified meromictic lake (Lake Alatsee, SW Germany). Environ. Microbial. 2015, 17, 2144–2157. [Google Scholar] [CrossRef]
- Princiotta, S.D.; Sanders, R.W. Heterotrophic and mixotrophic nanoflagellates in a mesotrophic lake: Abundance and grazing impacts across season and depth. Limnol Oceanogr. 2017, 62, 632–644. [Google Scholar] [CrossRef]
- Wilken, S.; Huisman, J.; Naus-Wiezer, S.; Van Donk, E. Mixotrophic organisms become more heterotrophic with rising temperature. Ecol. Lett. 2013, 16, 225–233. [Google Scholar] [CrossRef]
- Princiotta, S.D.; Smith, B.T.; Sanders, R.W. Temperature-dependent phagotrophy and phototrophy in a mixotrophic chrysophyte. J. Phycol. 2016, 52, 432–440. [Google Scholar] [CrossRef]
- Jones, H.L.J. A classification of mixotrophic protists based on their behavior. Freshw. Biol. 1997, 37, 35–43. [Google Scholar] [CrossRef]
- Mitra, A.; Flynn, K.J.; Tillmann, U.; Raven, J.A.; Caron, D.; Stoecker, D.K.; Not, F.; Hansen, P.J.; Hallegraeff, G.; Sanders, R.W.; et al. Defining planktonic protist functional groups on mechanisms for energy and nutrient acquisition: Incorporation of diverse mixotrophic strategies. Protist 2016, 167, 106–120. [Google Scholar] [CrossRef]
- Caron, D.A.; Sanders, R.W.; Lim, E.L.; Marrase, C.; Amaral, L.A.; Whitney, S.; Aoiki, R.B.; Porter, K.G. Light-dependent phagotrophy in the freshwater mixotrophic chrysophyte Dinobryon cylindricum. Microb. Ecol. 1993, 25, 93–111. [Google Scholar] [CrossRef]
- Wilken, S.; Verspagen, J.M.H.; Naus-Wiezer, S.; Van Donk, E.; Huisman, J. Biological control of toxic cyanobacteria by mixotrophic predators: An experimental test of intraguild predation theory. Ecol. Appl. 2014, 24, 1235–1249. [Google Scholar] [CrossRef]
- Holen, D.A. Effects of prey abundance and light intensity on the mixotrophy chrysophyte Poterioochromonas malhamensis from a mesotrophic lake. Freshw. Biol. 1999, 42, 445–455. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, H.Y.; Hong, Y.; Yang, J. Isolation of a Poterioochromonas capable of feeding on Microcystis aeruginosa and degrading microcystin-LR. FEMS Microbiol. Lett. 2008, 288, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.H.; Hong, S.S.; Song, S.Y.; Lee, H.O.; Nakano, S.; Han, M.S. Grazing Effects on Toxic and Non-Toxic Microcystis aeruginosa by the Mixotrophic Flagellate Ochromonas sp. J. Freshw. Ecol. 2009, 24, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Van Donk, E.; Cerbin, S.; Wilken, S.; Helmsing, N.R.; Ptacnik, R.; Verschoor, A.M. The effect of a mixotrophic chrysophyte on toxic and colony-forming cyanobacteria. Freshw. Biol. 2009, 54, 1843–1855. [Google Scholar] [CrossRef]
- Wilken, S.; Wiezer, S.; Huisman, J.; Van Donk, E. Microcystins do not provide anti-herbivore defense against mixotrophic flagellates. Aquat. Microb. Ecol. 2010, 59, 207–216. [Google Scholar] [CrossRef]
- Sanders, R.W.; Porter, K.G.; Caron, D.A. Relationship between phototrophy and phagotrophy in the mixotrophic chrysophyte Poterioochromonas malhamensis. Microb. Ecol. 1990, 19, 97–109. [Google Scholar] [CrossRef]
- Holen, D. The effects of heterotrophy on chlorophyll a and photosynthesis in a mixotrophic chrysophyte. Nova Hedwigia 2001, 122, 107–118. [Google Scholar]
- Brett, M.T.; Kainz, M.J.; Taipale, S.J.; Seshan, H. Phytoplankton, not allochthonous carbon, sustains herbivorous zooplankton production. Proc. Natl. Acad. Sci. USA 2009, 106, 21197–21201. [Google Scholar] [CrossRef] [Green Version]
- Hiltunen, M.; Honkanen, M.; Taipale, S.; Strandberg, U.; Kankaala, P. Trophic upgrading via the microbial food web may link terrestrial dissolved organic matter to Daphnia. J. Plankton Res. 2017, 39, 861–869. [Google Scholar] [CrossRef] [Green Version]
- Tranvik, L.; Porter, K.G.; Sieburth, J.M. Occurance of bacterivory in Cryptomonas, a common freshwater phytoplankter. Oecologia 1989, 78, 473–476. [Google Scholar] [CrossRef]
- Urabe, J.; Gurung, T.B.; Yoshida, T.; Sekino, T.; Nakanishi, M.; Maruo, M.; Nakayama, E. Diel changes in phagotrophy by Cryptomonas in Lake Biwa. Limnol. Oceanogr. 2000, 45, 1558–1563. [Google Scholar] [CrossRef] [Green Version]
- B-Beres, V.; Grigorszky, I.; Vasas, G.; Borics, G.; Varbiro, G.; Nagy, S.A.; Borbely, G.; Bacsi, I. The effects of Microcystis aeruginosa (cyanobacterium) on Cryptomonas ovata (Cryptophyta) in laboratory cultures: Why these organisms do not coexist in steady-state assemblages? Hydrobiologia 2012, 691, 97–107. [Google Scholar] [CrossRef]
- Grujcic, V.; Nuy, J.K.; Salcher, M.M.; Shabarova, T.; Kasalicky, V.; Boenigk, J.; Jensen, M.; Simek, K. Cryptophyta as major bacterivores in freshwater summer plankton. ISME J. 2018, 12, 1668–1681. [Google Scholar] [CrossRef]
- Worm, J.; Sondergaard, M. Dynamics of heterotrophic bacteria attached to Microcystis spp. (Cyanobacteria). Aquat. Microb. Ecol. 1998, 14, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Seymour, J.R.; Amin, S.A.; Raina, J.B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton-bacteria relationships. Nat. Microbiol. 2017, 2. [Google Scholar] [CrossRef]
- Kamjunke, N.; Boing, W.; Voigt, H. Bacterial and primary production under hypertrophic conditions. Aquat. Microb. Ecol. 1997, 13, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Kirkwood, A.E.; Nalewajko, C.; Fulthorpe, R.R. The effects of cyanobacterial exudates on bacterial growth and biodegradation of organic contaminants. Microb. Ecol. 2006, 51, 4–12. [Google Scholar] [CrossRef]
- Shen, H.; Niu, Y.; Xie, P.; Tao, M.; Yang, Xi. Morphological and physiological changes in Microcystis aeruginosa as a result of interactions with heterotrophic bacteria. Freshw. Biol. 2011, 56, 1065–1080. [Google Scholar] [CrossRef] [Green Version]
- Monchamp, M.E.; Pick, F.R.; Beisner, B.E.; Maranger, R. Nitrogen Forms Influence Microcystin Concentration and Composition via Changes in Cyanobacterial Community Structure. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Fu, Y.; O’Kelly, C.O.; Sieracki, M.; Distel, D.L. Protistan grazing analysis by flow cytometry using prey labeled by in vivo expression of fluorescent proteins. Appl. Environ. Microbiol. 2003, 69, 6848–6855. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Sherr, E.B.; Sherr, B.F. Differential feeding by marine flagellates on growing versus starving, and on motile versus nonmotile, bacterial prey. Mar. Ecol. Prog. Ser. 1993, 102, 257–267. [Google Scholar] [CrossRef]
- Ou, D.; Song, L.; Gan, N.; Chen, W. Effects of microcystins on and toxin degradation by Poterioochromonas sp. Environ. Toxicol. 2005, 20, 373–380. [Google Scholar] [CrossRef]
- Reiter, M.A. A simple fluorescent labeling technique for the marking of algae in mixed assemblages. J. Freshw. Ecol. 1997, 12, 163–165. [Google Scholar] [CrossRef]
- Sherr, B.F.; Sherr, E.B.; Fallon, R.D. Use of monodispersed, fluorescently labeled bacteria to estimate in-situ protozoan bacterivory. Appl. Environ. Microbiol. 1987, 53, 958–965. [Google Scholar]
- Shoemaker, J.; Tettenhorst, D.; Delacruz, A. Method 544: Determination of Microcystins and Nodularin in Drinking Water by Solid Phase Extraction and Liquid Chromatography/Tandem Mass Spectrometry (LC/MS/MS); U.S. Environmental Protection Agency: Washington, DC, USA, 2015.
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous interference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Variable | df | F | p |
| Microcystis alone | day | 1 | 12.309 | 0.025 |
| Microcystis + Cryptomonas | day | 2 | 14.888 | 0.005 |
| Microcystis + Cryptomonasfilt | day | 2 | 14.222 | 0.005 |
| Day of Experiment | df | F | p | |
| 0 | 2 | 5.600 * | 0.061 | |
| 4 | 1 | 21.400 | 0.010 | |
| 8 | 2 | 8.919 | 0.016 |
| Variable | df | F | p |
| Treatment | 3 | 19.350 | <0.001 |
| Time | 1 | 62.539 | <0.001 |
| Residual | 31 | - | - |
| Treatment | FLM Type | p | |
| Cryptomonas alone | tox-nt | <0.001 | |
| Cryptomonas + filtrate | tox-nt | 0.240 | |
| alone-filtrate | Tox | <0.001 | |
| alone-filtrate | nt | 0.190 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Princiotta, S.D.; Hendricks, S.P.; White, D.S. Production of Cyanotoxins by Microcystis aeruginosa Mediates Interactions with the Mixotrophic Flagellate Cryptomonas. Toxins 2019, 11, 223. https://doi.org/10.3390/toxins11040223
Princiotta SD, Hendricks SP, White DS. Production of Cyanotoxins by Microcystis aeruginosa Mediates Interactions with the Mixotrophic Flagellate Cryptomonas. Toxins. 2019; 11(4):223. https://doi.org/10.3390/toxins11040223
Chicago/Turabian StylePrinciotta, Sarah DeVaul, Susan P. Hendricks, and David S. White. 2019. "Production of Cyanotoxins by Microcystis aeruginosa Mediates Interactions with the Mixotrophic Flagellate Cryptomonas" Toxins 11, no. 4: 223. https://doi.org/10.3390/toxins11040223
APA StylePrinciotta, S. D., Hendricks, S. P., & White, D. S. (2019). Production of Cyanotoxins by Microcystis aeruginosa Mediates Interactions with the Mixotrophic Flagellate Cryptomonas. Toxins, 11(4), 223. https://doi.org/10.3390/toxins11040223