Occurrence of Mycotoxigenic Fusarium Species and Competitive Fungi on Preharvest Maize Ear Rot in Poland


Abstract
:1. Introduction
2. Results and Discussion
2.1. Occurrence of Fusarium, Trichoderma spp., S. zeae, and L. lecanii in Two Maize Producing Localities in Poland
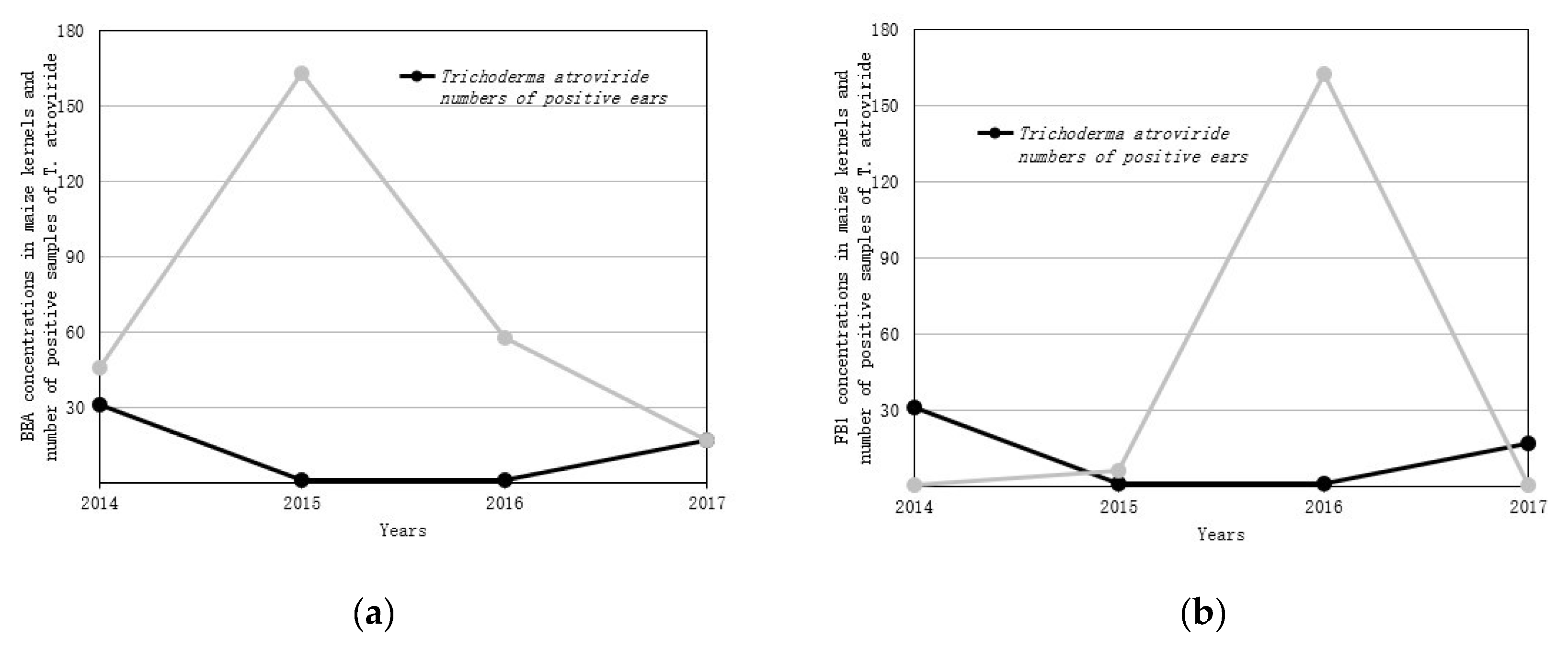
2.2. Mycotoxin Accumulation
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Fungal Isolation and Identification
3.3. Molecular Identification
3.4. Sample Preparation, Extraction and HPLC Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ksiezak, J.; Bojarczuk, J. The economic assessment of maize cultivation on pre-sowing tillage system. Acta Sci. Pol. Agric. 2010, 9, 55–67. [Google Scholar]
- White, D.G. Compendium of Corn Diseases, 3rd ed.; American Phytopathological Society Press: St Paul, MN, USA, 1999. [Google Scholar]
- Wit, M.; Warzecha, R.; Mirzwa-Mroz, E.; Jabłonska, E.; Ochodzki, P.; Waskiewicz, A.; Wakulinski, W. Susceptibility of flint and dent maize ears to Fusarium species. Phytopathologia 2011, 60, 35–45. [Google Scholar]
- Bottalico, A. Fusarium diseases of cereals: Species complex and related mycotoxins profiles in Europe. J. Plant Pathol. 1998, 80, 85–103. [Google Scholar]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 998–1003. [Google Scholar] [CrossRef]
- Munkvold, G.P. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 2003, 109, 705–713. [Google Scholar] [CrossRef]
- Pascale, M.; Visconti, A.; Chełkowski, J. Ear rot susceptibility and mycotoxin contamination of maize hybrids inoculated with Fusarium species under field conditions. Eur. J. Plant Pathol. 2002, 108, 645–651. [Google Scholar] [CrossRef]
- Bacon, C.W.; Bennett, R.M.; Hinton, D.M.; Voss, K.A. Scanning electron microscopy of Fusarium moniliforme within asymptomatic corn kernels and kernels associated with equine leukoencephalomalacia. Plant Dis. 1992, 76, 144–148. [Google Scholar] [CrossRef]
- Sobek, E.A. Associations between corn Insects and Symptomatic and Asymptomatic Corn Kernel Infection by Fusarium Moniliforme. Master’s Thesis, Iowa State University, Ames, Ames, IA, USA, 1996. [Google Scholar]
- Chełkowski, J. Distribution of Fusarium species and their mycotoxins in cereal grains. In Mycotoxins in Food and Agricultural Safety; Sinha, K., Bathnagar, D., Eds.; Marcel Dekker: New York, NY, USA, 1998; pp. 45–64. [Google Scholar]
- Gromadzka, K.; Górna, K.; Chełkowski, J.; Waśkiewicz, A. Mycotoxins and related Fusarium species in preharvest maize ear rot in Poland. Plant Soil Environ. 2016, 62, 348–354. [Google Scholar] [CrossRef]
- Gromadzka, K.; Wit, M.; Górna, K.; Chełkowski, J.; Waśkiewicz, A.; Ochodzki, P.; Warzecha, R. Fumonisins and related Fusarium species in pre-harvest maize ear rot in Poland. Cereal Res. Commun. 2017, 45, 93–103. [Google Scholar] [CrossRef]
- Logrieco, A.; Mule, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Eur. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Chełkowski, J. Mycotoxins associated with corn cob fusariosis. In Fusarium–Mycotoxins, Taxonomy and Pathogenicity; Chełkowski, J., Ed.; Elsevier: Amsterdam, The Netherlands, 1989; pp. 53–62. [Google Scholar]
- Tomczak, M.; Wiśniewska, H.; Stępień, Ł.; Kostecki, M.; Chełkowski, J.; Goliński, P. Deoxynivalenol, nivalenol and moniliformin occurrence in wheat samples with scab symptoms in Poland (1998–2000). Eur. J. Plant Pathol. 2002, 108, 625–630. [Google Scholar] [CrossRef]
- Goliński, P.; Waśkiewicz, A.; Wiśniewska, H.; Kiecana, I.; Mielniczuk, E.; Gromadzka, K.; Kostecki, M.; Bocianowski, J.; Rymaniak, E. Reaction of winter wheat (Triticum aestivum L.) cultivars to infection with Fusarium spp.: Mycotoxin contamination in grain and chaff. Food Addit. Contam. 2010, 27, 1015–1024. [Google Scholar]
- NUTRIAD Mycotoxin Survey. 2017. Available online: https://nutriad.com/wp content/uploads/2018/01/Article_Nutriad_Poland_2017_Mycotoxin_Survey_Maize.pdf (accessed on 10 January 2018).
- Ferrigo, D.; Raiola, A.; Rasera, R.; Causin, R. Trichoderma harzianum seed treatment controls Fusarium verticillioides colonization and fumonisin contamination in maize under field conditions. Crop Prot. 2014, 65, 51–56. [Google Scholar] [CrossRef]
- Vega, F.E.; Posada, F.; Aime, M.C.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic fungal endophytes. Biol. Control 2008, 46, 72–92. [Google Scholar] [CrossRef]
- Vidal, S.; Jaber, L.R. Entomopathogenic fungi as endophytes: Plant-endophyte-herbivore interactions and prospects for use in biological control. Curr. Sci. 2015, 109, 46–53. [Google Scholar]
- Shinde, S.V.; Patel, K.G.; Purohit, M.S.; Pandya, J.R.; Sabalpara, A.N. Lecanicillium lecanii (Zimm) Zare and Gams an important biocontrol agent for the management of insect pests—A Review. Agric. Rev. 2010, 31, 235–252. [Google Scholar]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational research on Trichoderma: From ‘omics to the field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Horwitz, B.A.; Kenerley, C.M. Secondary metabolism in Trichoderma—A genomic perspective. Microbiology 2012, 158, 35–45. [Google Scholar] [CrossRef]
- Chełkowski, J.; Ritieni, A.; Wisniewska, H.; Mule, G.; Logrieco, A. Occurrence of toxic hexadepsipeptides in preharvest maize ear rot infected by Fusariun poae in Poland. J. Phytopathol. 2007, 155, 8–12. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Komon, M.; Kubicek, C.P.; Druzhinina, I.S. Hypocrea voglmayrii sp. nov. from the Austrian Alps represents a new phylogenetic clade in Hypocrea/Trichoderma. Mycologia 2005, 97, 1365–1378. [Google Scholar] [CrossRef]
- Błaszczyk, L.; Popiel, D.; Chełkowski, J.; Koczyk, G.; Samuels, G.J.; Sobieralski, K.; Siwulski, M. Species diversity of Trichoderma in Poland. J. Appl. Genet. 2011, 52, 233–243. [Google Scholar] [CrossRef]
- Błaszczyk, L.; Basińska, A.; Ćwiek, H.; Gromadzka, K.; Popiel, D.; Stępień, Ł. Suppressive effect of Trichoderma on toxigenic Fusarium species. Pol. J. Microbiol. 2017, 1, 85–100. [Google Scholar] [CrossRef]
- Buśko, M.; Chelkowski, J.; Popiel, D.; Perkowski, J. Solid substrate bioassay to evaluate impact of Trichoderma on trichothecene mycotoxin production by Fusarium species. J. Sci. Food Agric. 2008, 88, 536–541. [Google Scholar] [CrossRef]
- Gromadzka, K.; Chełkowski, J.; Popiel, D.; Kachlicki, P.; Kostecki, M.; Goliński, P. Solid substrate bioassay to evaluate the effect of Trichoderma and Clonostachys on the production of zearalenone by Fusarium species. World Mycotoxin J. 2009, 2, 45–52. [Google Scholar] [CrossRef]
- Popiel, D.; Koczyk, G.; Dawidziuk, A.; Gromadzka, K.; Błaszczyk, L.; Chełkowski, J. Zearalenone lactonohydrolase activity in Hypocreales and its evolutionary relationships within the epoxide hydrolase subset of a/b-hydrolases. BMC Microbiol. 2014, 14, 82. [Google Scholar] [CrossRef]
- Jeleń, H.; Błaszczyk, L.; Chełkowski, J.; Rogowicz, K.; Strakowska, J. Formation of 6-n-pentyl-2H-pyran-2-one (6-PAP) and other volatiles by different Trichoderma species. Mycol. Prog. 2013, 13, 589–600. [Google Scholar] [CrossRef]
- Lumi Abe, C.A.; Bertechini Faria, C.; Castro, F.F.; Souza, S.R.; Santos, F.C.; Silva, C.N.; Tessmann, D.J.; Barbosa-Tessmann, I.P. Fungi isolated from maize (Zea mays L.) grains and production of associated enzyme activities. Ed. Ritva Tikkanen. Int. J. Mol. Sci. 2015, 16, 15328–15346. [Google Scholar] [CrossRef]
- Bhattacharya, K.; Raha, S. Deteriorative changes of maize, groundnut and soybean seeds by fungi in storage. Mycopathologia 2002, 155, 135–141. [Google Scholar] [CrossRef]
- Niaz, I.; Dawar, S. Detection of seed borne mycoflora in maize (Zea mays L.). Pak. J. Bot. 2009, 41, 443–451. [Google Scholar]
- Summerbell, R.C.; Gueidan, C.; Schreors, H.J.; de Hoog, G.S.; Staring, M.; Arocha Rosete, Y.; Guarro, J.; Scott, J.A. Acremonium phylogenetic overview and revision of Gliomastix, Sarocladium, and Trichothecium. Stud. Mycol. 2011, 68, 139–162. [Google Scholar] [CrossRef]
- Nirenberg, H.I. A simplified method for identifying Fusarium spp. occurring on wheat. Can. J. Bot. 1981, 59, 1599–1609. [Google Scholar] [CrossRef]
- Soman, A.G.; Gloer, J.B.; Angawi, R.F.; Wicklow, D.T.; Dowd, P.F. Vertilecanins: new phenopicolinic acid analogues from Verticillium lecanii. J. Nat. Prod. 2001, 64, 189–192. [Google Scholar] [CrossRef]
- Poling, S.M.; Wicklow, D.T.; Rogers, K.D.; Gloer, J.B. Acremonium zeae, a protective endophyte of maize, produces dihydroresorcylide and 7-hydroxydihydroresorcylides. J. Agric. Food Chem. 2008, 56, 3006–3009. [Google Scholar] [CrossRef]
- Pamphile, J.A.; Azwedo, J.L. Molecular characterization of endophytic strains of Fusarium verticillioides (=Fusarium moniliforme) from maize (Zea mays. L). World J. Microbiol. Biotechnol. 2002, 18, 391–396. [Google Scholar] [CrossRef]
- Wicklow, D.T.; Roth, S.; Deyrup, S.T.; Gloer, J.B. A protective endophyte of maize: Acremonium zeae antibiotics inhibitory to Aspergillus flavus and Fusarium verticillioides. Mycol. Res. 2005, 109, 610–618. [Google Scholar] [CrossRef]
- Giraldo, A.; Gené, J.; Cano, J.; Hoog, S.; Guarro, J. Two new species of Acremonium from Spanish soils. Mycologia 2012, 104, 1456–1465. [Google Scholar] [CrossRef]
- Yeh, Y.H.; Kirschner, R. Sarocladium spinificis, a new endophytic species from the coastal grass Spinifex littoreus in Taiwan. Bot. Stud. 2014, 55, 25. [Google Scholar] [CrossRef]
- Han, X.; Xu, W.; Zhang, J.; Xu, J.; Li, F. Natural occurrence of beauvericin and enniatins in corn- and wheat-based samples harvested in 2017 collected from Shandong Province, China. Toxins 2019, 11, 9. [Google Scholar] [CrossRef]
- Jurjevic, Z.; Solfrizzo, M.; Cvjetkovic, B.; De Girolamo, A.; Visconti, A. Occurrence of beauvericin in corn from Croatia. Food Technol. Biotechnol. 2002, 40, 91–94. [Google Scholar]
- Berardo, N.; Lanzanova, C.; Locatelli, S.; Laganà, P.; Verderio, A.; Motto, M. Levels of total fumonisins in maize samples from Italy during 2006–2008. Food Addit. Contam. Part B 2011, 4, 116–124. [Google Scholar] [CrossRef]
- Waalwijk, C.; Koch, S.; Ncube, E.; Allwood, J.; Flett, B.C.; de Vries, I.; Khema, G.H.J. Quantitativedetection of Fusarium spp. and its correlation with fumonisin content in maize from South African subsistence farmers. World Mycotoxins J. 2008, 1, 37–45. [Google Scholar] [CrossRef]
- Munkvold, G.P.; Hellmich, R.L.; Showers, W.B. Reduced Fusarium ear rot and symptomless infection in kernels of maize genetically engineered for European corn borer resistance. Phytopathology 1997, 87, 1071–1077. [Google Scholar] [CrossRef]
- Avantaggiato, G.; Quaranta, F.; Desiderio, E.; Visconti, A. Fumonisin contamination of maize hybrids visibly damaged by Sesamia. J. Sci. Food Agric. 2003, 83, 13–18. [Google Scholar] [CrossRef]
- Hermos, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef]
- Kostecki, M.; Grabarkiewicz-Szczęsna, J.; Chełkowski, J.; Wiśniewska, H. Beauvericin and moniliformin production by Polish isolates of Fusarium subglutinans and natural co-occurrence of both mycotoxins in maize samples. Microbiol.-Aliments-Nutr. 1995, 13, 67–70. [Google Scholar]
- Kwaśna, H.; Chełkowski, J.; Zajkowski, P. Fusarium (Sierpik) I; Institute of Botany, Polish Academy of Sciences: Warszawa, Poland, 1991. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blaackwell Publishing: Hoboken, NJ, USA, 2006; p. 388. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Jestoi, M.; Rokka, M.; Yli-Mattila, T.; Parikka, P.; Rizzo, A.; Peltonen, K. Presence and concentrations of the Fusarium-related mycotoxins beauvericin, enniatins and moniliformin in finish grain samples. Food Addit. Contam. 2004, 21, 794–802. [Google Scholar] [CrossRef]


{kind=link}
| Year | Locality 1 (Greater Poland Region) | Locality 2 (Silesia Region) | ||
|---|---|---|---|---|
| F. subglutinans | F. verticillioides | F. subglutinans | F. verticillioides | |
| 2014 | 54.5 | 45.4 | 9.3 | 47.7 |
| 2015 | 60.0 | 13.3 | 4.0 | 56.0 |
| 2016 | 55.8 | 11.8 | 0.0 | 57.1 |
| 2017 | 77.2 | 18.2 | 21.8 | 59.4 |
| Year | Numbers of Examined Ears | Trichoderma atroviride* Numbers of Positive Ears | Sarocladium zeae and Lecanicillium lecanii* Numbers of Positive Ears |
|---|---|---|---|
| 2014 | 100 | 31 | 1 |
| 2015 | 83 | 1 | 35 |
| 2016 | 58 | 1 | 8 |
| 2017 | 48 | 17 | 16 |
| total | 289 | 50 | 60 |
| Year | BEA [μg g−1] | Fumonisins [μg g−1] | |||||||
|---|---|---|---|---|---|---|---|---|---|
| FB1 | FB2 | FB3 | FB1 | FB2 | FB3 | ||||
| Locality 1 | Locality 2 | Locality 1 | Locality 2 | ||||||
| 2014 | average | 61.02 | 31.20 | 0.89 | 0.20 | 0.02 | 0.13 | 0.08 | 0.005 |
| maximum | 445.43 | 119.52 | 31.84 | 7.24 | 0.19 | 1.73 | 1.19 | 0.12 | |
| %positive | 84.62 | 50.00 | 50.00 | 15.00 | 25.00 | 58.70 | 21.74 | 19.57 | |
| 2015 | average | 201.33 | 124.10 | 6.54 | 0.33 | 0.08 | 5.52 | 0.93 | 0.13 |
| maximum | 1731.55 | 258.88 | 214.39 | 10.19 | 2.85 | 92.34 | 16.67 | 2.35 | |
| %positive | 94.59 | 100.00 | 83.78 | 56.76 | 54.05 | 95.23 | 95.23 | 42.86 | |
| 2016 | average | 44.21 | 70.79 | 75.41 | 26.91 | 4.33 | 248.85 | 71.93 | 19.01 |
| maximum | 474.56 | 130.99 | 465.00 | 282.97 | 72.89 | 1418.34 | 660.75 | 219.88 | |
| %positive | 89.29 | 96.15 | 35.71 | 35.71 | 35.71 | 69.23 | 69.23 | 69.23 | |
| 2017 | average | 32.25 | 1.84 | 0.22 | 0.021 | 0.005 | 1.39 | 0.13 | 0.01 |
| maximum | 140.69 | 24.14 | 0.81 | 0.094 | 0.022 | 10.21 | 0.92 | 0.11 | |
| %positive | 95.24 | 50.00 | 57.89 | 57.89 | 47.37 | 53.85 | 53.85 | 42.31 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gromadzka, K.; Błaszczyk, L.; Chełkowski, J.; Waśkiewicz, A. Occurrence of Mycotoxigenic Fusarium Species and Competitive Fungi on Preharvest Maize Ear Rot in Poland. Toxins 2019, 11, 224. https://doi.org/10.3390/toxins11040224
Gromadzka K, Błaszczyk L, Chełkowski J, Waśkiewicz A. Occurrence of Mycotoxigenic Fusarium Species and Competitive Fungi on Preharvest Maize Ear Rot in Poland. Toxins. 2019; 11(4):224. https://doi.org/10.3390/toxins11040224
Chicago/Turabian StyleGromadzka, Karolina, Lidia Błaszczyk, Jerzy Chełkowski, and Agnieszka Waśkiewicz. 2019. "Occurrence of Mycotoxigenic Fusarium Species and Competitive Fungi on Preharvest Maize Ear Rot in Poland" Toxins 11, no. 4: 224. https://doi.org/10.3390/toxins11040224
APA StyleGromadzka, K., Błaszczyk, L., Chełkowski, J., & Waśkiewicz, A. (2019). Occurrence of Mycotoxigenic Fusarium Species and Competitive Fungi on Preharvest Maize Ear Rot in Poland. Toxins, 11(4), 224. https://doi.org/10.3390/toxins11040224