Application of an Endothelial Cell Culture Assay for the Detection of Neutralizing Anti-Clostridium Perfringens Beta-Toxin Antibodies in a Porcine Vaccination Trial

 ,
, 
Abstract
:1. Introduction
2. Results
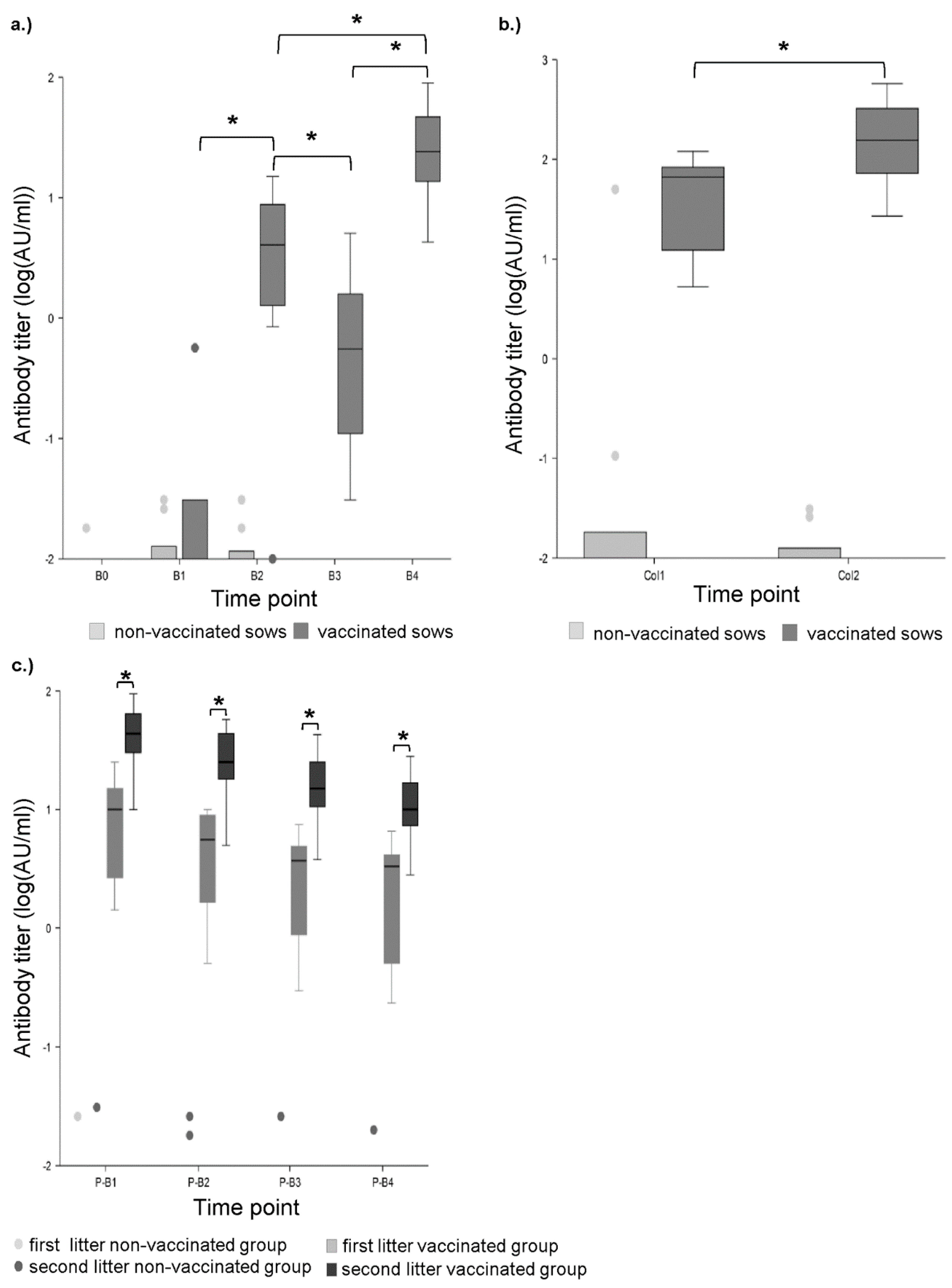
2.1. Total Anti-Beta-Toxin Antibodies Determined by ELISA
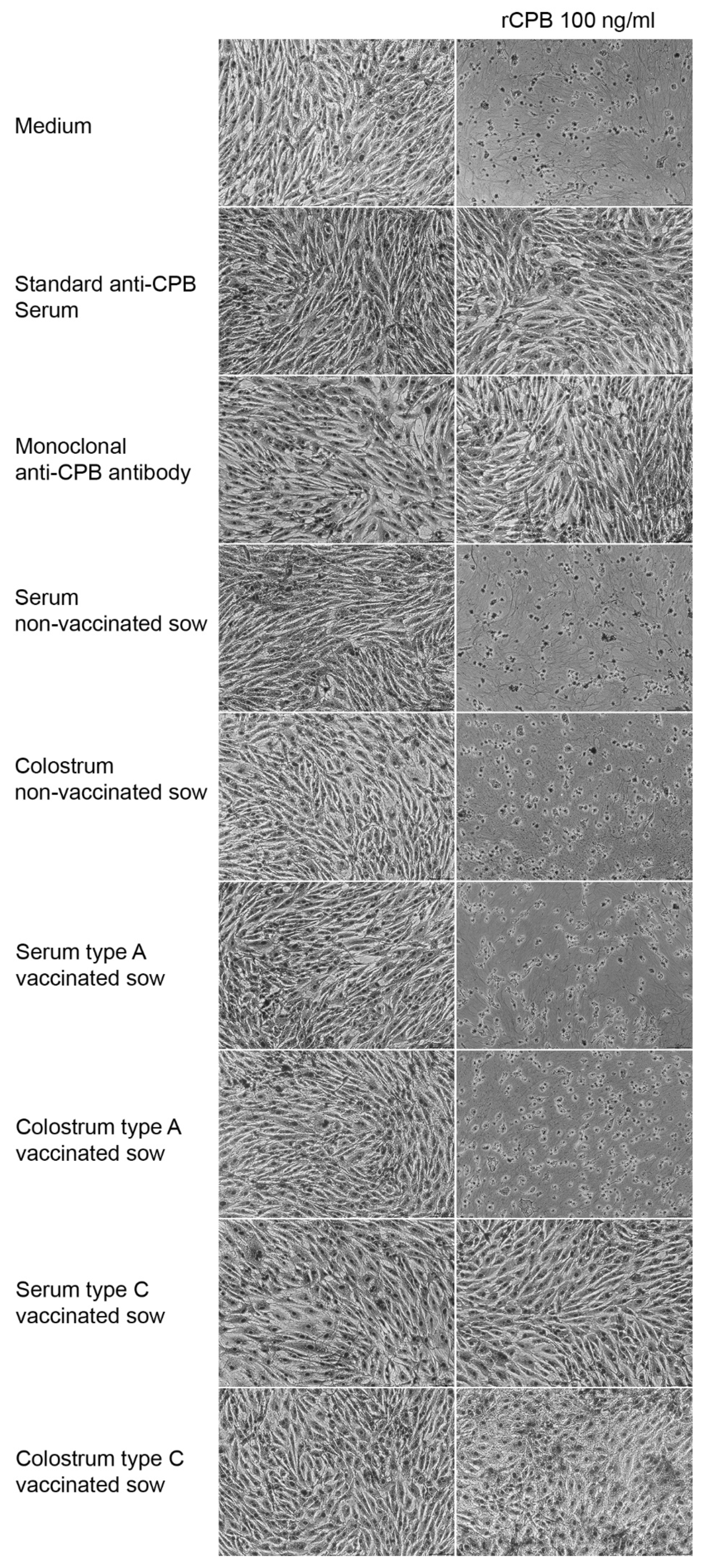
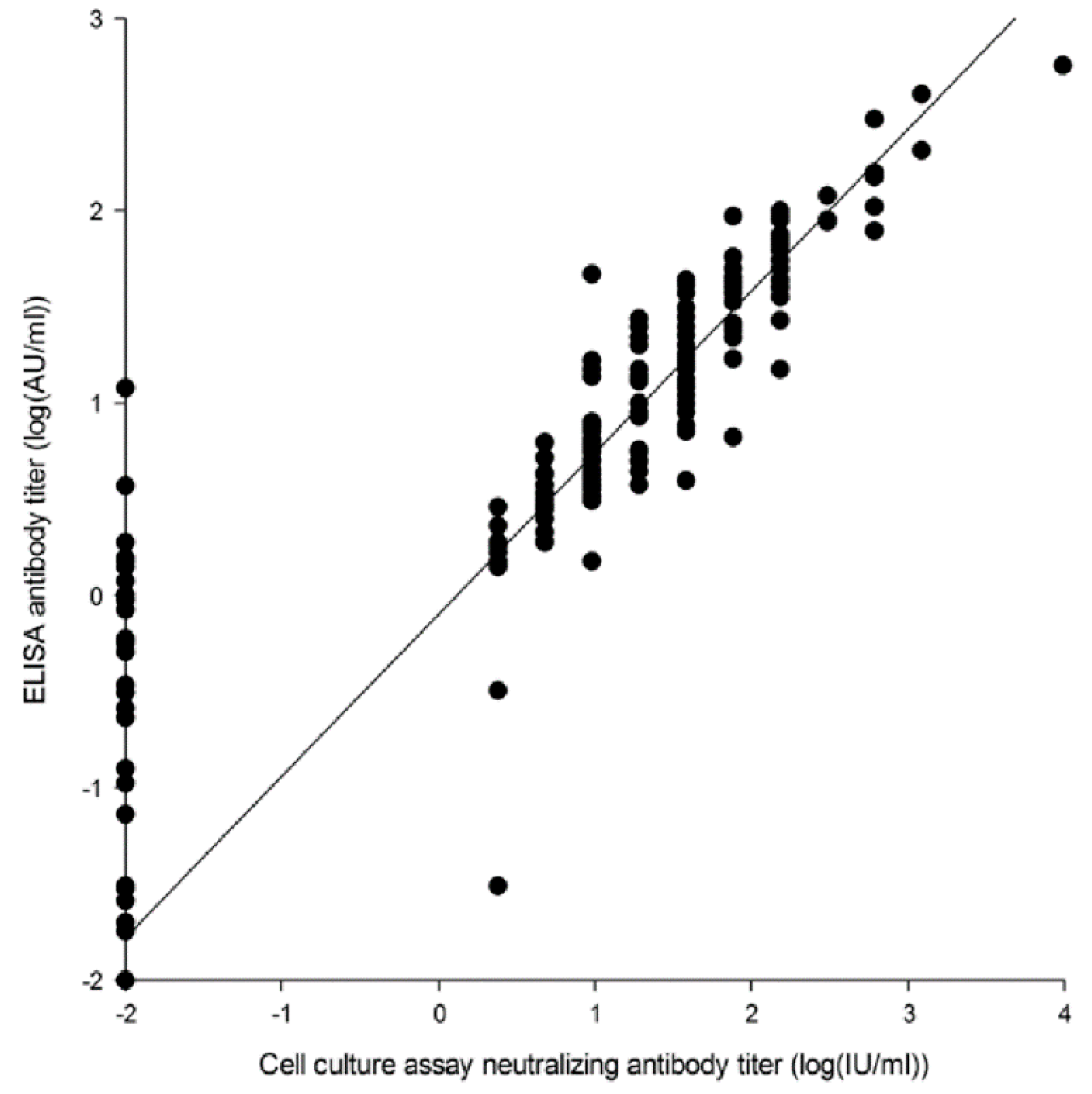
2.1.1. Cell Culture Assay
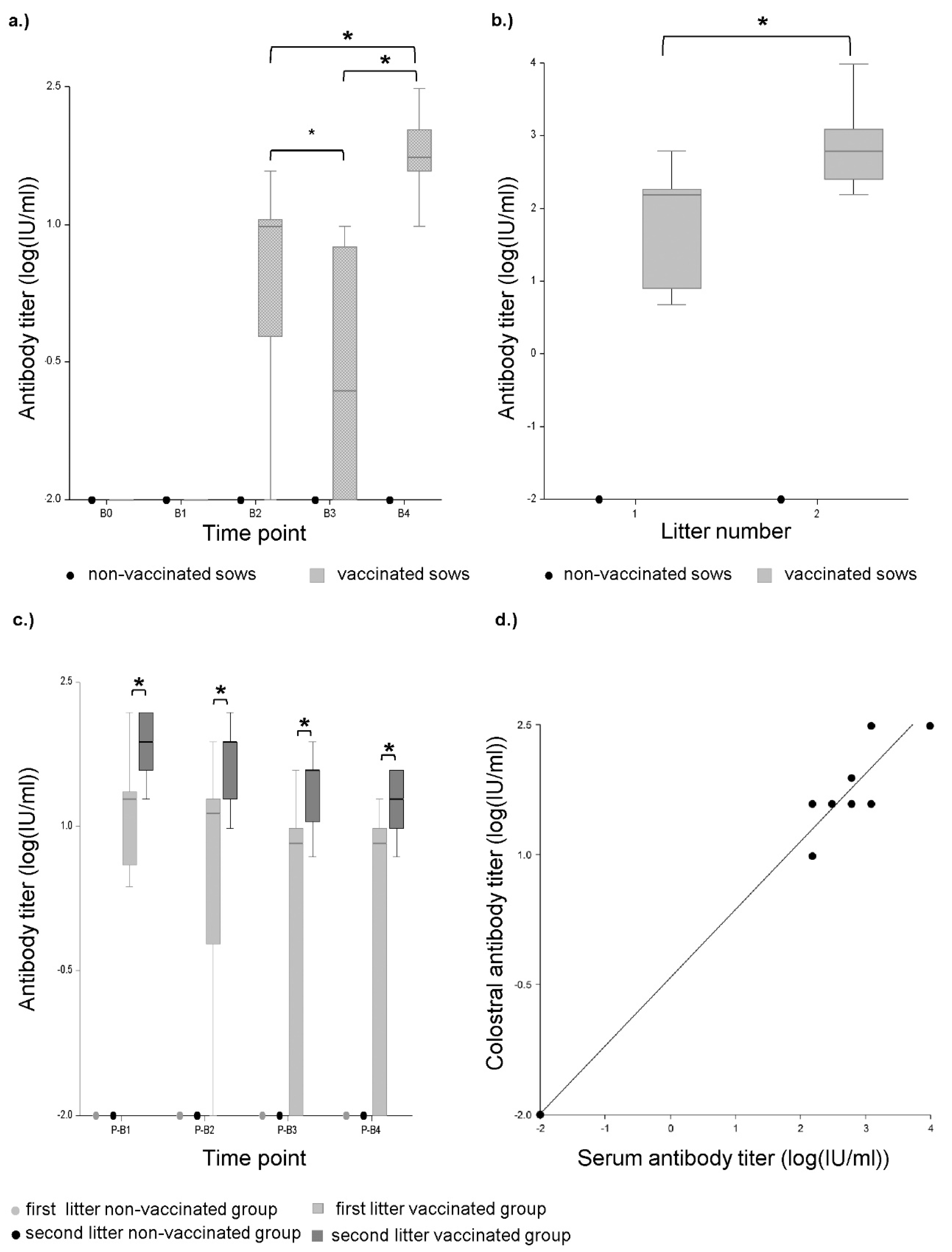
2.1.2. Challenge Experiment
3. Discussion
4. Materials and Methods
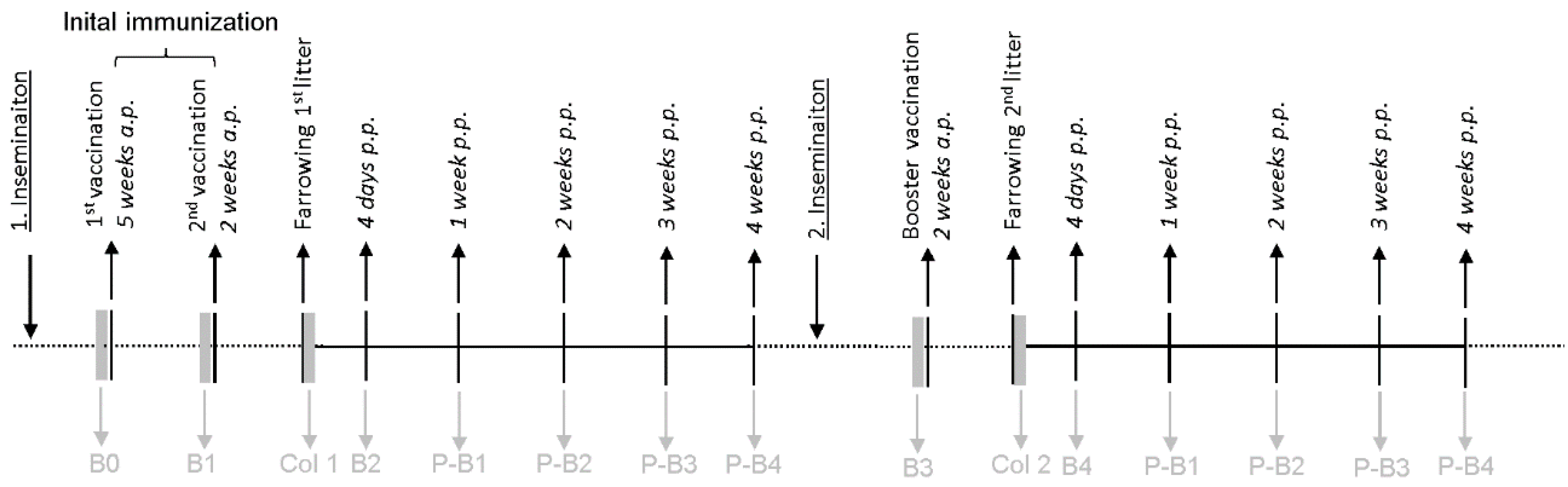
4.1. Vaccination Trial
4.2. Recombinant Beta-Toxin
4.3. ELISA
4.4. Cell Culture Assay
4.5. Challenge Experiments
4.6. Statistics
4.7. Ethics Approval
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Songer, J.G.; Uzal, F.A. Clostridial enteric infections in pigs. J. Vet. Diagn. Investig. 2005, 17, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, M.; Ochi, S.; Oda, M.; Miyamoto, K.; Takehara, M.; Kobayashi, K. Recent insights into Clostridium perfringens beta-toxin. Toxins 2015, 7, 396–406. [Google Scholar] [CrossRef]
- Gurtner, C.; Popescu, F.; Wyder, M.; Sutter, E.; Zeeh, F.; Frey, J.; von Schubert, C.; Posthaus, H. Rapid cytopathic effects of Clostridium perfringens beta-toxin on porcine endothelial cells. Infect. Immun. 2010, 78, 2966–2973. [Google Scholar] [CrossRef] [PubMed]
- Popescu, F.; Wyder, M.; Gurtner, C.; Frey, J.; Cooke, R.A.; Greenhill, A.R.; Posthaus, H. Susceptibility of primary human endothelial cells to C. perfringens beta-toxin suggesting similar pathogenesis in human and porcine necrotizing enteritis. Vet. Microbiol. 2011, 153, 173–177. [Google Scholar] [CrossRef]
- Schumacher, V.L.; Martel, A.; Pasmans, F.; Van Immerseel, F.; Posthaus, H. Endothelial binding of beta toxin to small intestinal mucosal endothelial cells in early stages of experimentally induced Clostridium perfringens type C enteritis in pigs. Vet. Pathol. 2013, 50, 626–629. [Google Scholar] [CrossRef]
- Lawrence, G.; Walker, P.D.; Freestone, D.S.; Shann, F. The prevention of pig-bel in Papua New Guinea. Papua New Guin. Med. J. 1979, 22, 30–34. [Google Scholar]
- Lawrence, G.W.; Lehmann, D.; Anian, G.; Coakley, C.A.; Saleu, G.; Barker, M.J.; Davis, M.W. Impact of active immunisation against enteritis necroticans in Papua New Guinea. Lancet 1990, 336, 1165–1167. [Google Scholar] [CrossRef]
- Springer, S.; Selbitz, H.J. The control of necrotic enteritis in sucking piglets by means of a Clostridium perfringens toxoid vaccine. FEMS Immunol. Med. Microbiol. 1999, 24, 333–336. [Google Scholar] [CrossRef]
- Djurickovic, S.M.; Dworak, J.E.; Wickham, K.L. Antitoxin titer in colostrum and milk after vaccination of sows with Clostridium perfringens type C toxoid vaccine. Vet. Med. Small Anim. Clin. 1975, 70, 283–285. [Google Scholar]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe 2018, 53, 5–10. [Google Scholar] [CrossRef]
- Vidal, J.E.; McClane, B.A.; Saputo, J.; Parker, J.; Uzal, F.A. Effects of Clostridium perfringens beta-toxin on the rabbit small intestine and colon. Infect. Immun. 2008, 76, 4396–4404. [Google Scholar] [CrossRef] [PubMed]
- Sayeed, S.; Uzal, F.A.; Fisher, D.J.; Saputo, J.; Vidal, J.E.; Chen, Y.; Gupta, P.; Rood, J.I.; McClane, B.A. Beta toxin is essential for the intestinal virulence of Clostridium perfringens type C disease isolate CN3685 in a rabbit ileal loop model. Mol. Microbiol. 2008, 67, 15–30. [Google Scholar] [CrossRef]
- Schafer, K.; Wyder, M.; Gobeli, S.; Candi, A.; Doherr, M.G.; Zehnder, B.; Zimmermann, W.; Posthaus, H. Detection of Clostridium perfringens type C in pig herds following disease outbreak and subsequent vaccination. Vet. Rec. 2012, 171, 503. [Google Scholar] [CrossRef]
- Schafer, K.; Zimmermann, W.; Posthaus, H. Possible influence of herd health management and hygiene on the in-herd prevalence of Clostridium perfringens type C in pig breeding farms. Schweiz. Arch. Tierheilkund. 2013, 155, 520–522. [Google Scholar] [CrossRef]
- Hogh, P. Experimental studies on serum treatment and vaccination against Cl. perfringens type C infection in piglets. Dev. Boil. Stand. 1976, 32, 69–76. [Google Scholar]
- Kennedy, K.K.; Norris, S.J.; Beckenhauer, W.H.; Hogg, A. Vaccination of pregnant sows with Clostridium perfringens type C toxoid. Vet. Med. Small Anim. Clin. 1977, 72, 1047–1049. [Google Scholar] [PubMed]
- Matisheck, P.H.; McGinley, M. Colostral transfer of Clostridium perfringens type C beta antitoxin in swine. Am. J. Vet. Res. 1986, 47, 1132–1133. [Google Scholar]
- Ripley, P.H.; Gush, A.F. Immunisation schedule for the prevention of infectious necrotic enteritis caused by Clostridium perfringens type C in piglets. Vet. Rec. 1983, 112, 201–202. [Google Scholar] [CrossRef] [PubMed]
- Ebert, E.; Öppling, V.; Werner, E.; Cubetaler, K. Development and prevalidation of alternative methods for the potency testing of C. perfringens vaccines. ALTEX 1997, 15, 59–61. [Google Scholar]
- Solanki, A.K.; Bhatia, B.; Kaushik, H.; Deshmukh, S.K.; Dixit, A.; Garg, L.C. Clostridium perfringens beta toxin DNA prime-protein boost elicits enhanced protective immune response in mice. Appl. Microbiol. Biotechnol. 2017, 101, 5699–5708. [Google Scholar] [CrossRef]
- Miclard, J.; Jaggi, M.; Sutter, E.; Wyder, M.; Grabscheid, B.; Posthaus, H. Clostridium perfringens beta-toxin targets endothelial cells in necrotizing enteritis in piglets. Vet. Microbiol. 2009, 137, 320–325. [Google Scholar] [CrossRef] [PubMed]
- European Pharmacopoeia Commission; European Directorate for the Quality of Medicines & Healthcare. European Pharmacopoeia; Council of Europe: Strasbourg, France, 2016. [Google Scholar]
- Steinthorsdottir, V.; Halldorsson, H.; Andresson, O.S. Clostridium perfringens beta-toxin forms multimeric transmembrane pores in human endothelial cells. Microb. Pathog. 2000, 28, 45–50. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morbidity Rate | Mortality Rate | ||||||
|---|---|---|---|---|---|---|---|
| Trial | Group | Sows (n) | Piglets (n) | n | In % | n | In % |
| 1 | 2 x vaccination | 10 | 20 | 0 * | 0 | 0 * | 0 |
| 1 | Control | 8 | 16 | 13 | 81.25 | 11 | 68.75 |
| 2 | 3 x vaccination | 9 | 18 | 0 * | 0 | 0 * | 0 |
| 2 | Control | 10 | 20 | 19 | 95.00 | 18 | 90.00 |
| Parameter | Clinical Findings and Score |
|---|---|
| General condition (general clinical impression) | 0 = undisturbed |
| 1 = disturbed | |
| Posture | 0 = normal, body supported equally on all 4 limbs, animal moves physiologically |
| 1 = unstable gait, kyphosis | |
| 2 = animal lies permanently | |
| Behaviour | 0 = normal, animal is alert |
| 1 = apathetic | |
| 2 = somnolent | |
| Milk up-take | 0 = yes |
| 1 = no | |
| Nutritional status * | 0 = good |
| 1 = medium (spine and pelvic bone easily recognisable) | |
| 2 = cachexia | |
| Respiration | 0 = normal |
| 1 = increased respiratory rate | |
| 2 = distinct abdominal breathing | |
| Ascites | 0 = no |
| 1 = slight | |
| 2 = severe | |
| Skin turgor | 0 = < 1 s (after pulling skin fold in neck region) |
| 1 = 1 – 3 s | |
| 2 = > 3 s | |
| Diarrhoea | 0 = no, formed faeces |
| 1 = yes, pasty | |
| 2 = yes, runny | |
| 3 = yes, pasty or runny, bloody, or black feces |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richard, O.K.; Springer, S.; Finzel, J.; Theuß, T.; Wyder, M.; Vidondo, B.; Posthaus, H. Application of an Endothelial Cell Culture Assay for the Detection of Neutralizing Anti-Clostridium Perfringens Beta-Toxin Antibodies in a Porcine Vaccination Trial. Toxins 2019, 11, 225. https://doi.org/10.3390/toxins11040225
Richard OK, Springer S, Finzel J, Theuß T, Wyder M, Vidondo B, Posthaus H. Application of an Endothelial Cell Culture Assay for the Detection of Neutralizing Anti-Clostridium Perfringens Beta-Toxin Antibodies in a Porcine Vaccination Trial. Toxins. 2019; 11(4):225. https://doi.org/10.3390/toxins11040225
Chicago/Turabian StyleRichard, Olivia K., Sven Springer, Jacqueline Finzel, Tobias Theuß, Marianne Wyder, Beatriz Vidondo, and Horst Posthaus. 2019. "Application of an Endothelial Cell Culture Assay for the Detection of Neutralizing Anti-Clostridium Perfringens Beta-Toxin Antibodies in a Porcine Vaccination Trial" Toxins 11, no. 4: 225. https://doi.org/10.3390/toxins11040225
APA StyleRichard, O. K., Springer, S., Finzel, J., Theuß, T., Wyder, M., Vidondo, B., & Posthaus, H. (2019). Application of an Endothelial Cell Culture Assay for the Detection of Neutralizing Anti-Clostridium Perfringens Beta-Toxin Antibodies in a Porcine Vaccination Trial. Toxins, 11(4), 225. https://doi.org/10.3390/toxins11040225