Abnormal Development of Hyalomma Marginatum Ticks (Acari: Ixodidae) Induced by Plant Cytotoxic Substances

 ,
,  , ,
, ,
Abstract
:1. Introduction
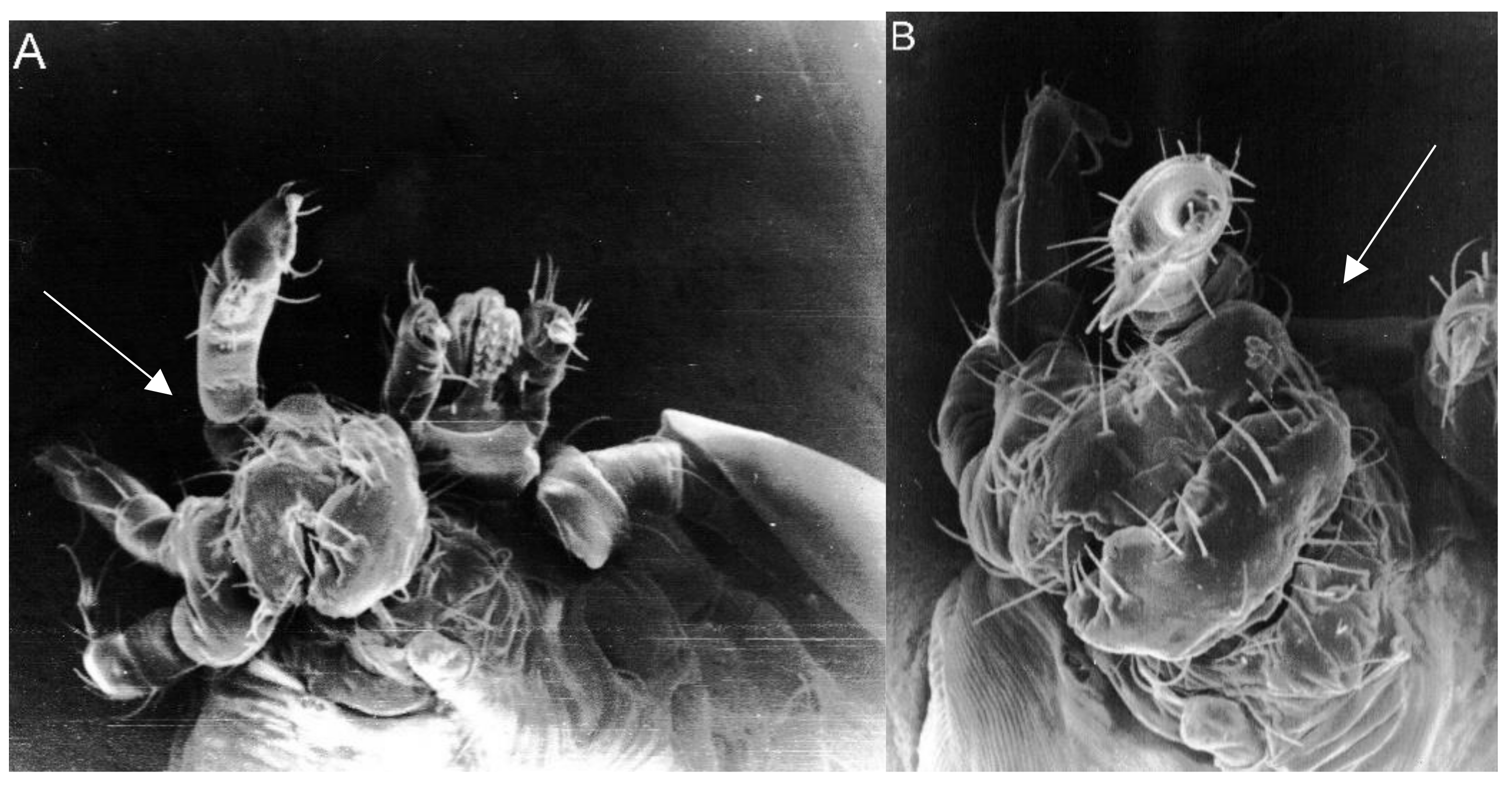
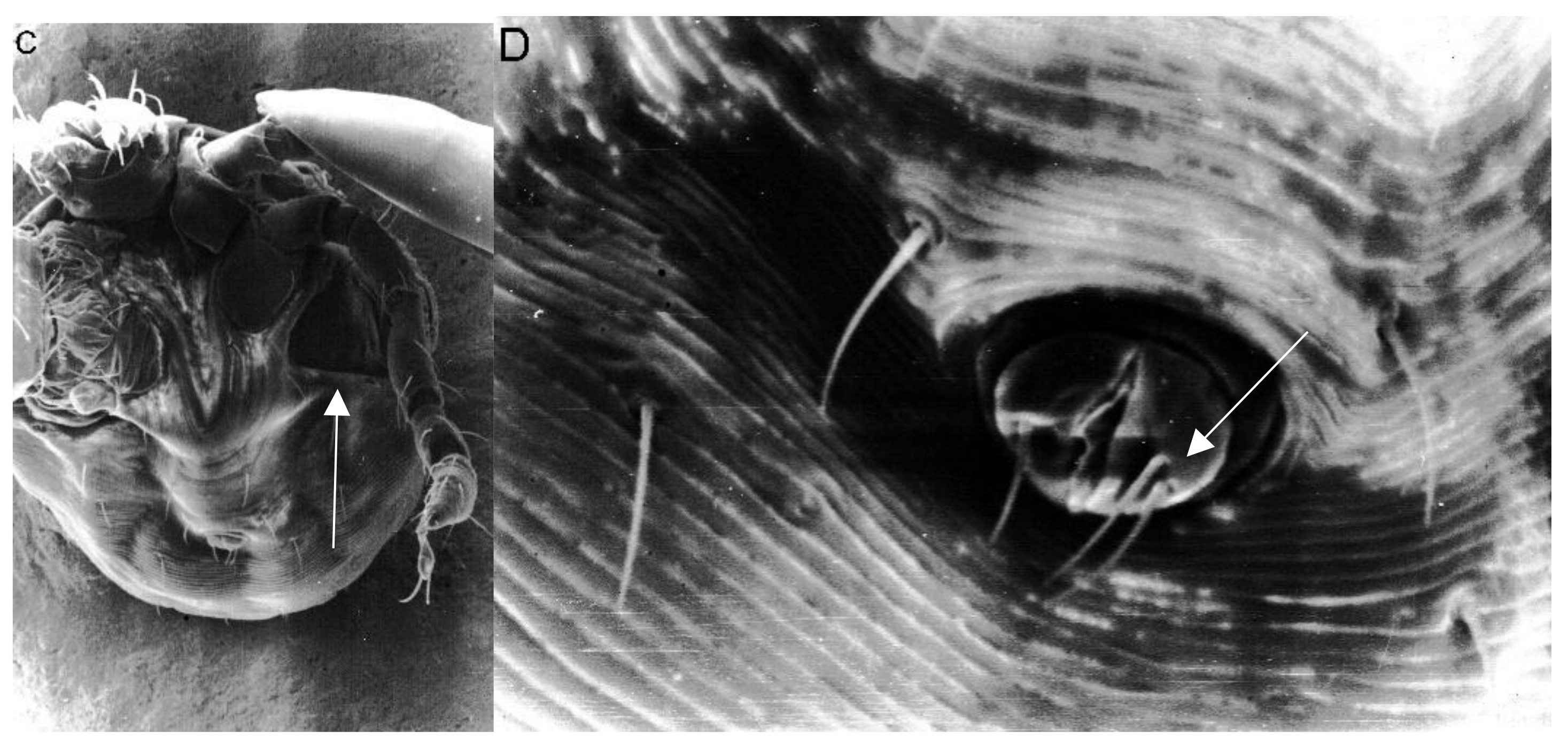
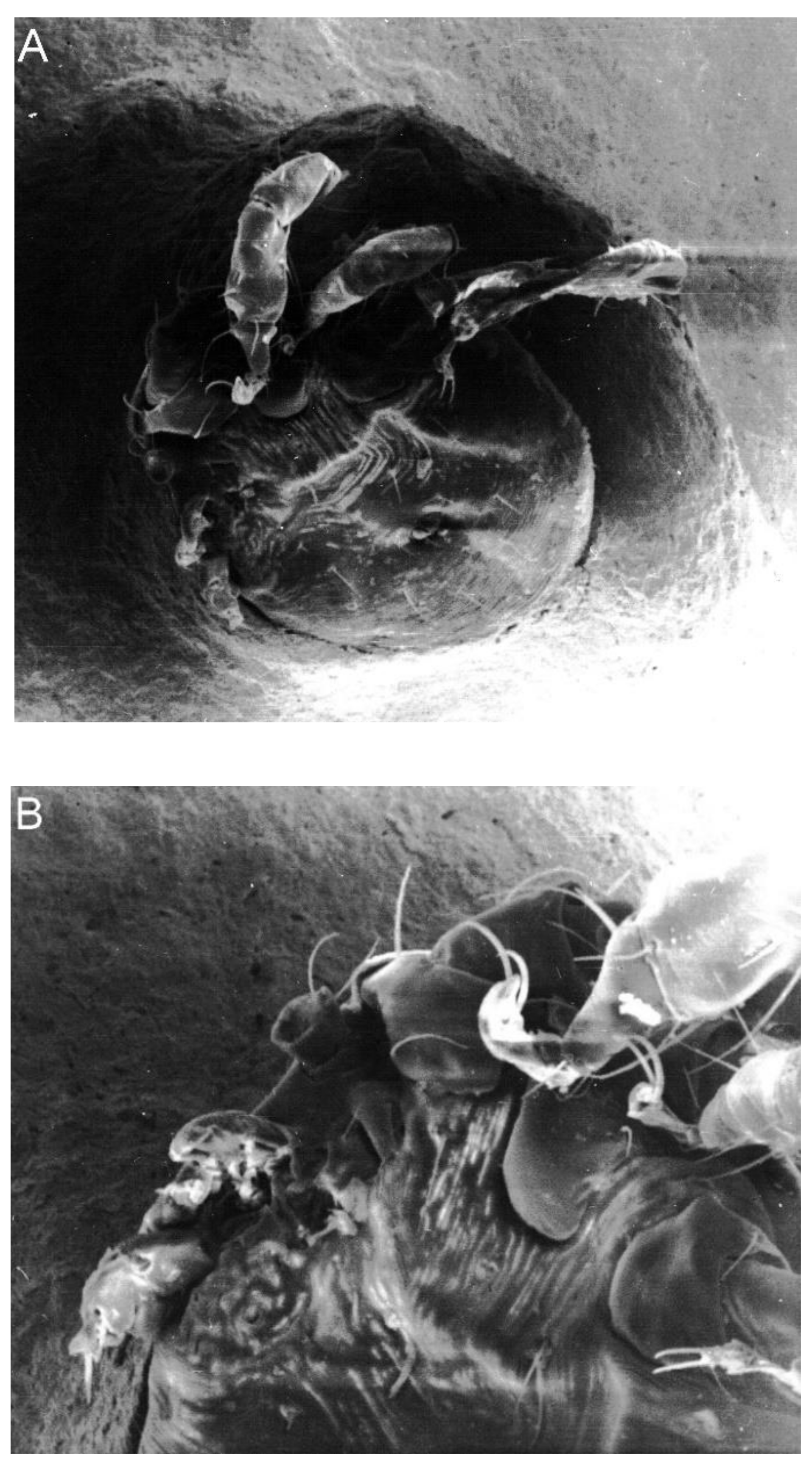
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
Ethical Statement
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hoogstraal, H. Ticks of the Sudan, with Special Reference to Equatoria Province and with Preliminary Reviews to the Genera Boophilus, Margaropus and Hyalomma; Naval Medical Research Unit-3: Washington, DC, USA, 1956. [Google Scholar]
- Kolonin, G.V. Fauna of Ixodid Ticks of the World (Acari, Ixodidae); Nauka: Moscow, Russia, 2009. [Google Scholar]
- Nosek, J.; Folk, C. Relationships of birds to arboviruses and their vectors. Acta Sci. Nat. Acad. Sci. Bohemoslov. Brno 1977, 11, 1–61. [Google Scholar]
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Durrani, A.Z.; Shakoori, A.R.; Kamal, N. Bionomics of Hyalomma ticks in three districts of Punjab, Pakistan. J. Anim. Plant Sci. 2008, 18, 17–23. [Google Scholar]
- Movila, A.; Alekseev, A.N.; Dubinina, H.V.; Toderas, I. Detection of tick-borne pathogens in ticks from migratory birds in the Baltic region of Russia. Med. Vet. Entomol. 2013, 27, 113–117. [Google Scholar] [CrossRef]
- Orkun, O.; Karaer, Z.; Çakmak, A.; Nalbantoglu, S. Identification of Tick-Borne Pathogens in Ticks Feeding on Humans in Turkey. PLoS Negl. Trop. Dis. 2014, 8, e3067. [Google Scholar] [CrossRef] [PubMed]
- Wallménius, K.; Barboutis, C.; Fransson, T.; Jaenson, T.G.T.; Lindgren, P.-E.; Nyström, F.; Olsen, B.; Salaneck, E.; Nilsson, K. Spotted fever Rickettsia species in Hyalomma and Ixodes ticks infesting migratory birds in the European Mediterranean area. Parasit. Vectors 2014, 7, 318. [Google Scholar] [CrossRef] [PubMed]
- Ehounoud, C.B.; Yao, K.P.; Dahmani, M.; Achi, Y.L.; Amanzougaghene, N.; N’Douba, A.K.; N’Guessan, J.D.; Raoult, D.; Fenollar, F.; Mediannikov, O. Multiple pathogens including potential new species in tick vectors in Cote d’Ivoire. PLoS Negl. Trop. Dis. 2016, 10, e0004367. [Google Scholar] [CrossRef] [PubMed]
- Gargili, A.; Estrada-Peña, A.; Spengler, J.R.; Lukashev, A.; Nuttall, P.A.; Bente, D.A. The role of ticks in the maintenance and transmission of Crimean-Congo hemorrhagic fever virus: A review of published field and laboratory studies. Antivir. Res. 2017, 144, 93–119. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Martínez Avilés, M.; Muñoz Reoyo, M.J. A population model to describe the distribution and seasonal dynamics of the tick Hyalomma marginatum in the Mediterranean Basin. Transbound. Emerg. Dis. 2011, 58, 213–223. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Sańchez, N.; Estrada-Sańchez, A. An assessment of the distribution and spread of the tick Hyalomma marginatum in the Western Palearctic under different climate scenarios. Vector Borne Zoonotic Dis. 2012, 12, 758–768. [Google Scholar] [CrossRef]
- Geevarghese, G.; Dhanda, V. The feeding pattern of Hyalomma ticks on different host, and its evolutionary significance. Curr. Sci. 1982, 51, 424–425. [Google Scholar]
- Magano, S.R.; Els, D.A.; Chown, S.L. Feeding patterns of immature stages of Hyalomma truncatum and Hyalomma marginatum rufipes on different hosts. Exp. Appl. Acarol. 2000, 24, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.M.; Norval, R.A.I.; Rechav, Y. The life cycle tick Hyalomma marginatum rufipes Koch under laboratory conditions. J. Parasitol. 1978, 64, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Ouhelli, H. Comparative development of Hyalomma marginatum (Koch, 1844), H. detritum (Schulze, 1919), H. anatolicum excavatum (Koch, 1844), H. lusitanicum (Koch, 1844) and H. dromedarii (Koch, 1844) under laboratory conditions. Acta Parasitol. 1994, 39, 153–157. [Google Scholar]
- Gharbi, M.; Darghouth, M.A. A review of Hyalomma scupense (Acari, Ixodidae) in the Maghreb region: From biology to control. Parasite 2014, 21, 2. [Google Scholar] [CrossRef] [PubMed]
- Buczek, A. Wirkungen von Temperatur und Luftfeuchtigkeit auf Embryonalentwicklung und das Eischlupfen bei der Schildzecke Hyalomma marginatum Koch (Acari, Ixodidae). Anzeiger für Schädlingskunde Pflanzenschutz Umweltschutz 1993, 66, 6–9. [Google Scholar] [CrossRef]
- Buczek, A. Experimental teratogeny in the tick Hyalomma marginatum marginatum (Acari: Ixodida: Ixodidae): Effect of high humidity on embryonic development. J. Med. Entomol. 2000, 37, 807–814. [Google Scholar] [CrossRef]
- Duke, S.O.; Cantrell, C.L.; Meepagala, K.M.; Wedge, D.E.; Tabanca, N.; Schrader, K.T. Natural toxins for use in pest management. Toxins 2010, 2, 1943–1962. [Google Scholar] [CrossRef]
- Borges, L.M.; Sousa, L.A.; Barbosa Cda, S. Perspectives for the use of plant extracts to control the cattle tick Rhipicephalus (Boophilus) microplus. Rev. Bras. Parasitol. Vet. 2011, 20, 89–96. [Google Scholar] [CrossRef]
- Ghosh, S.; Tiwari, S.S.; Srivastava, S.; Sharma, A.K.; Kumar, S.; Ray, D.D.; Rawat, A.K. Acaricidal properties of Ricinus communis leaf extracts against organophosphate and pyrethroids resistant Rhipicephalus (Boophilus) microplus. Vet. Parasitol. 2013, 192, 259–267. [Google Scholar] [CrossRef]
- Ghosh, S.; Tiwari, S.S.; Kumar, B.; Srivastava, S.; Sharma, A.K.; Kumar, S.; Bandyopadhyay, A.; Julliet, S.; Kumar, R.; Rawat, A.K. Identification of potential plant extracts for anti-tick activity against acaricide resistant cattle ticks, Rhipicephalus (Boophilus) microplus (Acari: Ixodidae). Exp. Appl. Acarol. 2015, 66, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Adenubi, O.T.; Fasina, F.O.; McGaw, L.J.; Eloff, J.N.; Naidoo, V. Plant extracts to control ticks of veterinary and medical importance: A review. S. Afr. J. Bot. 2016, 105, 178–193. [Google Scholar] [CrossRef]
- Adenubi, O.T.; Ahmed, A.S.; Fasina, F.O.; McGaw, L.J.; Eloff, J.N.; Naidoo, V. Pesticidal plants as a possible alternative to synthetic acaricides in tick control: A systematic review and meta-analysis. Ind. Crops Prod. 2018, 123, 779–806. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Canale, A.; Mehlhorn, H. Tick repellents and acaricides of botanical origin: A green roadmap to control tick-borne diseases? Parasitol. Res. 2016, 115, 2545–2560. [Google Scholar] [CrossRef] [PubMed]
- Rosado-Aguilar, J.A.; Ariona-Cambranes, K.; Torres-Acosta, J.F.J.; Rodrigez-Vivas, R.I.; Bolio-González, M.E.; Ortega-Pacheco, A.; Alzina-López, A.; Gutiérrez-Ruiz, E.J.; Gutiérrez-Blanco, E.; Aguilar-Caballero, A.J. Plant products and secondary metabolites with acaricide activity against ticks. Vet. Parasitol. 2017, 238, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Pavela, R. Repellence of essential oils and selected compounds against ticks-A systematic review. Acta Trop. 2018, 179, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Madzimure, J.; Nyahangare, E.T.; Hamudikuwanda, H.; Hove, T.; Stevenson, P.C.; Belmain, S.R.; Mvumi, B.M. Acaricidal efficacy against cattle ticks and acute oral toxicity of Lippia javanica (Burm F.) Spreng. Trop. Anim. Health Prod. 2011, 43, 481–489. [Google Scholar] [CrossRef]
- Wanzala, W. Potential of Traditional Knowledge of Plants in the Management of Arthropods in Livestock Industry with Focus on (Acari) Ticks. Evid. Based Complement. Alternat. Med. 2017, 2017, 8647919. [Google Scholar] [CrossRef]
- Alekseev, A.N.; Dubinina, H.V. Abnormalities in Ixodes ticks (Ixodoidea, Ixodinae), Acarina. Russ. J. Acarol. 1993, 1, 73–85. [Google Scholar]
- Alekseev, A.N.; Dubinina, H.V. Some aspects of mite (Oppiidae) and tick (Ixodidae) pathology as a result of anthropogenic pressure. In Acarology IX; Mitchell, R., Horn, D.J., Needham, G.R., Welbourn, W.C., Eds.; The Ohio Biological Survey: Columbus, OH, USA, 1996; Volume 1, pp. 117–120. [Google Scholar]
- Buczek, A. Reduction in egg hatch of Argas (Argas) reflexus (Fabr.) (Acari, Argasidae) due to colchicine. Anzeiger für Schädlingskunde Pflanzenschutz Umweltschutz 1993, 66, 131–134. [Google Scholar] [CrossRef]
- Zharkov, S.D.; Dubinina, H.V.; Alekseev, A.N.; Jensen, P.M. Anthropogenic pressure and changes in Ixodes tick populations in the Baltic region of Russia and Denmark. Acarina 2000, 2, 137–141. [Google Scholar]
- Alekseev, A.N.; Dubinina, H.V.; Jääskeläinen, A.E.; Vapalahti, O.; Vaheri, A. First report on tick-borne pathogens and exoskeletal anomalies in Ixodes persulcatus Schulze (Acari: Ixodidae) collected in Kokkola coastal region, Finland. Int. J. Acarol. 2007, 33, 253–258. [Google Scholar] [CrossRef]
- Sshuchinova, L.D. Occurrence of ixodic ticks with exoskeleton anomalies and their infection with tick-borne encephalitis in Altai Republic. Russ. J. Parasitol. 2014, 2, 18–21. (In Russian) [Google Scholar]
- Černý, V. Abnormity u některȳch zăstupcú středoevropské fauny klīšťat. Cas. Csl. Spol. Entomol. 1957, 54, 162–171. [Google Scholar]
- Tovornik, D. Teratological forms of ixodid ticks. Biol. Vestn. 1987, 35, 91–100. [Google Scholar]
- Guglielmone, A.A.; Castella, J.; Mangold, A.J.; Estrada-Peña, A.; Vinabal, A.E. Phenotypic anomalies in a collection of neotropical ticks (Ixodidae). Acarologia 1999, 40, 127–132. [Google Scholar]
- Dergousoff, S.J.; Chilton, N.B. Abnormal morphology of an adult Rocky Mountain wood tick, Dermacentor andersoni (Acari: Ixodidae). J. Parasitol. 2007, 93, 708–709. [Google Scholar] [CrossRef] [PubMed]
- Aeschlimann, A. Dévelopment embryonnaire d’Ornithodorus moubata (Murray) et transmission transovarienne de Borrelia duttoni. Acta Trop. 1958, 15, 15–64. [Google Scholar]
- Aeschlimann, A. Complement a l’etude de l’embryologie d’Ornithodoros moubata (Murray). Acta Trop. 1961, 18, 58–60. [Google Scholar]
- El Kammah, K.M.; Adham, F.K.; Tadross, N.R.; Osman, M. Embryonic development of the camel tick Hyalomma dromedarii (Ixodoidea: Ixodidae). Int. J. Acarol. 1982, 8, 47–54. [Google Scholar] [CrossRef]
- Aeschlimann, A.; Hess, E. What is our current knowledge of acarine embryology. In Acarology VI; Griffiths, D.A., Bowman, C.E., Eds.; Ellis Horwood: Chichester, UK, 1984; pp. 90–99. [Google Scholar]
- Balashov, Y.S. Embryonic development. In Taiga Ticks Ixodes Persulcatus Schulze (Acarina, Ixodidae); Filippova, N.A., Ed.; Nauka Publ.: Leningrad, Russia, 1985; pp. 93–95. (In Russian) [Google Scholar]
- El Gohary, M.; Kamel, M.Y.; Madbouly, M.H. On the morphology of developing eggs of the camel tick Hyalomma dromedarii Koch, 1844. Can. J. Zool. 1986, 64, 1994–1997. [Google Scholar] [CrossRef]
- Shiraishi, S.; Yano, Y.; Uchida, T.A. Embryogenesis in the cattle tick, Haemaphysalis longicornis. J. Fac. Agric. Kyushu Univ. 1990, 34, 265–272. [Google Scholar]
- Buczek, A. Influence of iodine compounds on embryogenesis of Argas (A.) reflexus (Fabricius, 1794) (Acari: Ixodida: Argasidae). Acta Parasitol. 1993, 38, 41–43. [Google Scholar]
- Buczek, A. Inhibitory factors of Argas (Argas) reflexus (Fab.) (Acari, Argasidae) larvae development. Anzeiger für Schädlingskunde Pflanzenschutz Umweltschutz 1996, 69, 23–25. [Google Scholar] [CrossRef]
- Campana, Y. Schizomelies spontanees et experimentales chez les Ixodoidea. Ann. Parasit. Hum. Comp. 1947, 22, 53–62. [Google Scholar] [CrossRef]
- Ioffe, I.D. Possible correlation between the reduction of legs IV in the larvae and the development of the nervous apparatus in the Parasitiformes (Acarina). In Acarology VI; Griffiths, D.A., Bowman, C.E., Eds.; Ellis Horwood Limited: Chichester, UK, 1984; Volume 1, pp. 286–294. [Google Scholar]
- Buczek, A.; Bartosik, K.; Kuczyński, P. Evaluation of the effect of various concentrations of selected pyrethroids on the development of Dermacentor reticulatus eggs and larvae. Ann. Agric. Environ. Med. 2013, 20, 447–451. [Google Scholar] [PubMed]
- Buczek, A.; Bartosik, K.; Kuczyński, P. Sensitivity to permethrin in a Dermacentor reticulatus population from eastern Poland in laboratory study. Parasit. Vectors 2014, 7, 18. [Google Scholar] [CrossRef]
- Buczek, A.; Bartosik, K.; Buczek, W.; Buczek, A.M.; Kuczyński, P. The effect of sublethal concentrations of deltamethrin and alphacypermethrin on the fecundity and development of Ixodes ricinus (Acari: Ixodidae) eggs and larvae. Exp. Appl. Acarol. 2019, 78, 203–221. [Google Scholar] [CrossRef]
- Oliver, J.H., Jr.; Delfin, E.D. Gynandromorphism in Dermacentor occidentalis (Acari: Ixodidae). Ann. Entomol. Soc. Am. 1967, 60, 1119–1121. [Google Scholar] [CrossRef]
- Buczek, A. Influence of high relative humidity on course of embryonic development and egg hatch of Argas (A.) reflexus (Fabricius, 1794)(Acari: Ixodida: Argasidae). Z. Angew. Zool. 1991, 4, 339–443. [Google Scholar]
- Buczek, A. Studies on the biology of Argas reflexus (Fabricius, 1794) (Acari: Ixodida: Argasidae). 2. Effect of altering temperatures on embryonic development and egg hatch. Folia Biol. 1992, 40, 151–153. [Google Scholar]
- Buczek, A. Disturbances of embryonic development and egg hatch of Argas (A.) reflexus (Fabricius, 1794) (Ixodida: Argasidae) caused by altering temperatures. In The Acari. Physiological Aspects of Acari-Host Relationships; Kropczyńska, D., Boczek, J., Tomczyk, A., Eds.; Publishing House Dabor: Warszawa, Poland, 1995; pp. 351–355. [Google Scholar]
- Buczek, A. Studies on the biology of Argas (A.) reflexus (Fabricius, 1794) (Acari: Ixodida: Argasidae). I. Effect temperature and relative humidity on embryonic development and egg hatch. Folia Biol. 1988, 36, 239–264. [Google Scholar]
- Juberthie, C. Monstrousités observés chez les Opilions. Bull. Mus. Nat. Hist. Nat. 1963, 35, 167–171. [Google Scholar]
- Juberthie, C. Recherchez sur la Biologie des Opilions. Ph.D. Thesis, Centre National de la Recherche Scientifique, Paris, France, 1964. [Google Scholar]
- Juberthie, C. Tératologie expérimentale chez un Opilion (Arachnidae). J. Embryol. Exp. Morph. 1968, 19, 49–82. [Google Scholar]
- Jacuński, L. Temperature induced developmental monstrosities in Tegenaria atrica C. L. Koch (Araneae, Agelenidae). Zool. Pol. 1971, 21, 285–316. (In Polish) [Google Scholar]
- Jacuński, L. Studies on Experimental Teratogeny in the Spider Tegenaria Atrica C. L. Koch; Academic Press: Toruń, Poland, 1984. [Google Scholar]
- Mikulska, I. Experimentally induced developmental monstrosities in the water spider Argyroneta aquatica (Clerk). Zool. Pol. 1973, 22, 127–134. [Google Scholar]
- Napiórkowska, T.; Kobak, J.; Napiórkowski, P.; Templin, J. The effect of temperature and light on embryogenesis and post-embryogenesis of the spider Eratigena atrica (Araneae, Agelenidae). J. Therm. Biol. 2018, 72, 26–32. [Google Scholar] [CrossRef]
- Sekigushi, S. The occurrence of double embryos in spider eggs after centrifugation. Arachnol. News 1952, 1, 1–4. [Google Scholar]
- Ehn, A. The development of spider embryos after centrifugation. Zool. Bidrag. Upps. 1962, 35, 339–368. [Google Scholar]
- Jacuński, L. Inducement of developmental monstrosities in the spider Tegenaria atrica C. L. Koch by centrifugation of mature females. Zool. Pol. 1969, 19, 589–600. (In Polish) [Google Scholar]
- Willadsen, P. Immunity to ticks. Adv. Parasitol. 1980, 18, 293–313. [Google Scholar]
- Latif, A.A.; Newson, R.M.; Dhadilla, T.S. Feeding performance of Amblyomma variegatum (Acarina: Ixodidae) fed repeatedly on rabbits. Exp. Appl. Acarol. 1988, 5, 83–92. [Google Scholar] [CrossRef]
- Bartosik, K.; Buczek, A.; Borzęcki, A.; Kulina, D. Study of the non-parasitic stage in Ixodes ricinus after co-feeding with Dermacentor reticulatus in three infestations. Ann. Agric. Environ. Med. 2017, 24, 90–95. [Google Scholar] [CrossRef]
- Latif, A.A.; Dhadialla, T.S.; Newson, R.M. Abnormal development of Amblyomma variegatum (Acarina: Ixodidae). J. Med. Entomol. 1988, 25, 142–143. [Google Scholar] [CrossRef]
- Estrada-Peña, A. Abnormal development of Rhipicephalus sanguineus (Ixodidae). Exp. Appl. Acarol. 2001, 25, 757–761. [Google Scholar] [CrossRef]
- Buczek, A.; Stanisławek, I.M.; Bartosik, K.; Kruk, T.; Olszewski, T. Local anomalies in larvae of ticks (Acari: Ixodida). Wiad. Parazytol. 2004, 50, 295–301. (In Polish) [Google Scholar]
- Buczek, A.; Bartosik, K.; Buczek, S. Four gynandromorphs of Hyalomma marginatum marginatum ticks (Acari: Ixodidae) from a laboratory colony. J. Nat. Hist. 2014, 48, 1947–1956. [Google Scholar] [CrossRef]
- Buczek, A.; Bartosik, K.; Buczek, S.; Zając, Z. Anomalies in Hyalomma marginatum larvae (Acari: Ixodidae) in relation to taxonomic studies. Syst. Appl. Acarol. 2017, 22, 423–430. [Google Scholar] [CrossRef]
- Slovák, M.; Vidlička, L. Hard tick (Acarina: Ixodidae) malformations. Entomofauna Carpathica 2013, 25, 13–24. [Google Scholar]
- Ren, Q.; Chen, Z.; Luo, J.; Liu, G.; Guan, G.; Yin, H.; Luo, J. Abnormal development of Haemaphysalis qinghaiensis (Acari: Ixodidae). J. Insect Sci. 2016, 16, 66. [Google Scholar] [CrossRef]
- Campana-Rouget, Y. La teratology des tiques (1). Ann. Parasit. Hum. Comp. 1959, 34, 209–260. [Google Scholar] [CrossRef]
- Buczek, A.; Siuda, K.; Alsied, S. Morphological anomalies in ticks (Acari: Ixodidae) collected from nature. Wiad. Parazytol. 1991, 37, 31–34. (In Polish) [Google Scholar]
- Labruna, M.B.; Riberio, A.F.; Cruz, M.V.; Camargo, L.M.; Camargo, E.P. Gynandromorphism in Amblyomma cajennense and Rhipicephalus sanguineus (Acari: Ixodidae). J. Parasitol. 2002, 88, 810–811. [Google Scholar] [CrossRef]
- Nowak-Chmura, M. Teratological changes in tick morphology in ticks feeding on exotic reptiles. J. Nat. Hist. 2012, 46, 911–921. [Google Scholar] [CrossRef]
- Kar, S.; Akyildiz, G.; Yilmazer, N.; Shaibi, T.; Gargili, A.; Vatansever, Z. External morphological anomalies in ixodid ticks from Thrace, Turkey. Exp. Appl. Acarol. 2015, 67, 457–466. [Google Scholar] [CrossRef]
- Prusinski, M.A.; Mertins, J.W.; Meehan, L.J. Two Gynandromorphs of Ixodes scapularis (Acari: Ixodidae) from New York State. J. Med. Entomol. 2015, 52, 278–282. [Google Scholar] [CrossRef]
- Keskin, A.; Simsek, E.; Bursali, A.; Keskin, A. Morphological abnormalities in ticks (Acari: Ixodidae) feeding on humans in Central Black Sea region, Turkey. Zoomorphology 2016, 135, 167–172. [Google Scholar] [CrossRef]
- Chitimia-Dobler, L.; Bestehorn, M.; Bröker, M.; Borde, J.; Molcanyi, T.; Skaarup Andersen, N.; Pfeffer, M.; Dobler, G. Morphological anomalies in Ixodes ricinus and Ixodes inopinatus collected from tick-borne encephalitis natural foci in Central Europe. Exp. Appl. Acarol. 2017, 72, 379–397. [Google Scholar] [CrossRef]
- Chitimia-Dobler, L.; Pfeffer, M. Gynandromorphism and local morphological abnormalities in Dermacentor reticulatus (Acari: Ixodidae). Syst. Appl. Acarol. 2017, 22, 449–455. [Google Scholar]
- Molaei, G.; Little, E.A.H. A nine-legged tick: Report of a morphological anomaly in the blacklegged tick, Ixodes scapularis (Acari: Ixodidae) from the northeastern United States. Ticks Tick Borne Dis. 2018, 9, 778–780. [Google Scholar] [CrossRef]
- Alekseev, A.N.; Jensen, P.M.; Dubinina, H.V.; Smirnova, L.A.; Makrouchina, N.A.; Zharkov, S.D. Peculiarities of behaviour of taiga (Ixodes persulcatus) and sheep (Ixodes ricinus) ticks (Acarina: Ixodida) determined by different method. Folia Parasitol. 2000, 47, 147–153. [Google Scholar] [CrossRef]
- Žygutienė, M.; Alekseev, A.; Dubinina, H.; Kazlauskienė, R. Evidence for a risk of tick-borne infection in the city parks of Vilnius, Lithuania. Ekologija 2008, 54, 40–43. [Google Scholar] [CrossRef]
- Alekseev, A.N.; Dubinina, H.V. Enhancement of risk of tick-borne infection: Environmental and parasitological aspects of the problem. J. Med. Entomol. 2008, 45, 812–815. [Google Scholar] [CrossRef]
- Gargili, A.; Thangamani, S.; Bente, D. Influence of laboratory animal hosts on the life cycle of Hyalomma marginatum and implications for an in vivo transmission model for Crimean-Congo hemorrhagic fever virus. Front. Cell. Infect. Microbiol. 2013, 3, 39. [Google Scholar] [CrossRef]
- Winston, P.H.; Bates, D.H. Saturated solutions for control of humidity in biological research. Ecology 1960, 41, 232–237. [Google Scholar] [CrossRef]
- Corwin, D.; Clifford, C.M.; Keirans, J.E. An improved method for cleaning and preparing ticks for examination with the scanning electron microscope. J. Med. Entomol. 1979, 16, 352–353. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Developmental Stages | Examined Group | Control a | p Values | Significance |
|---|---|---|---|---|
| No. of eggs examined | 41,512 | 4267 | ||
| Dead eggs | 16.3 | 11.0 | 0.4142 | no |
| Dead embryos | 9.8 | 0.6 | 0.5930 | no |
| Abnormally hatched larvae | 8.1 | 0.6 | 0.6698 | no |
| Larvae with anomalies | 0.2 | - | 0.7150 | no |
| Normal larvae | 65.6 | 87.8 | 0.7456 | no |
| Type of Anomalies | Anomalies | |
|---|---|---|
| Number | % | |
| Gnathosoma | 16 | 22.5 |
| Oligomely of the palp | 5 | 7.0 |
| Atrophy of the palp | 1 | 1.4 |
| Deformation of the palp | 3 | 4.2 |
| Abnormal position of the palp | 1 | 1.4 |
| Elongation of chelicerae | 5 | 7.0 |
| Deformation of chelicerae | 1 | 1.4 |
| Idiosoma | 49 | 67.6 |
| Changed shape of the idiosoma | 1 | 1.4 |
| Oligomely of legs | 11 | 15.5 |
| Atrophy of legs | 2 | 2.8 |
| Heterosymely of legs | 3 | 4.2 |
| Schistomely of legs | 1 | 1.4 |
| Deformation of segments | 25 | 35.2 |
| Atrophy of segments | 2 | 2.8 |
| Other leg anomalies | 4 | 5.6 |
| Composite anomalies | 6 | 8.5 |
| Total | 71 | 100 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buczek, A.; Bartosik, K.; Buczek, A.M.; Buczek, W.; Kulina, D. Abnormal Development of Hyalomma Marginatum Ticks (Acari: Ixodidae) Induced by Plant Cytotoxic Substances. Toxins 2019, 11, 445. https://doi.org/10.3390/toxins11080445
Buczek A, Bartosik K, Buczek AM, Buczek W, Kulina D. Abnormal Development of Hyalomma Marginatum Ticks (Acari: Ixodidae) Induced by Plant Cytotoxic Substances. Toxins. 2019; 11(8):445. https://doi.org/10.3390/toxins11080445
Chicago/Turabian StyleBuczek, Alicja, Katarzyna Bartosik, Alicja M. Buczek, Weronika Buczek, and Dorota Kulina. 2019. "Abnormal Development of Hyalomma Marginatum Ticks (Acari: Ixodidae) Induced by Plant Cytotoxic Substances" Toxins 11, no. 8: 445. https://doi.org/10.3390/toxins11080445
APA StyleBuczek, A., Bartosik, K., Buczek, A. M., Buczek, W., & Kulina, D. (2019). Abnormal Development of Hyalomma Marginatum Ticks (Acari: Ixodidae) Induced by Plant Cytotoxic Substances. Toxins, 11(8), 445. https://doi.org/10.3390/toxins11080445