Impact of Naturally Contaminated Substrates on Alphitobius diaperinus and Hermetia illucens: Uptake and Excretion of Mycotoxins


 ,
,
Abstract
:1. Introduction
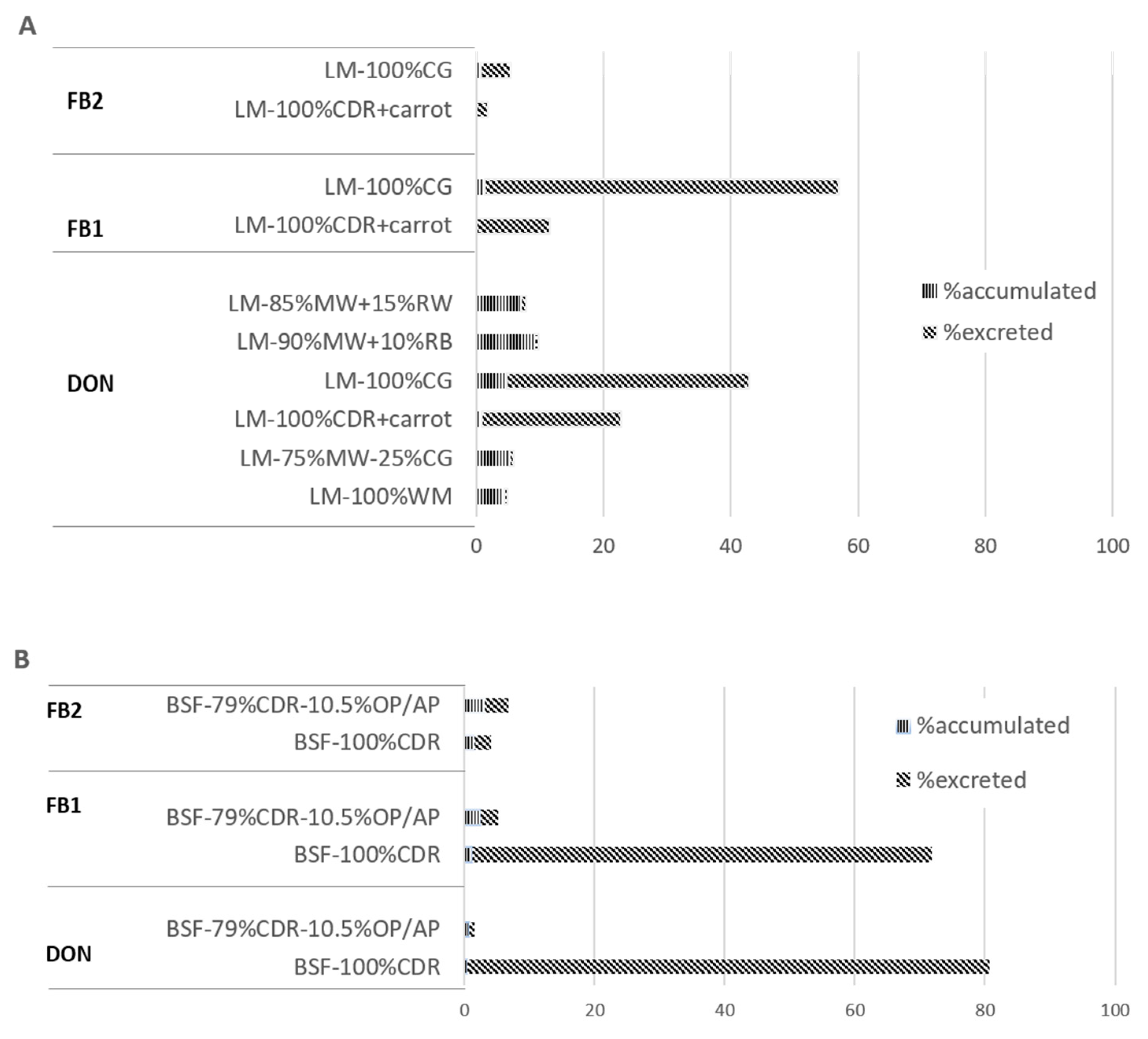
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Insect Treatments and Sampling
5.3. Mycotoxins Extraction and Purification
5.4. Mycotoxins Analysis
5.5. Mass Balance Calculation
Author Contributions
Funding
Conflicts of Interest
References
- Food Wastage Footprint: Impacts on Natural Resources (Summary Report). Available online: http://www.fao.org/3/i3347e/i3347e.pdf (accessed on 18 August 2019).
- Veldkamp, T.; Van Duinkerken, G.; Van Huis, A.; Lakemond, C.M.M.; Ottevanger, E.; Bosch, G.; Van Boekel, M.A.J.S. Insects as a Sustainable Feed Ingredient in Pig and Poultry Diets—A Feasibility Study; Wageningen University Livestock Research: Wageningen, The Netherlands, 2012. [Google Scholar]
- Van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef] [Green Version]
- Charlton, A.J.; Dickinson, M.; Wakefield, M.E.; Fitches, E.; Kenis, M.; Han, R.; Zhu, F.; Kone, N.; Grant, M.; Devic, E.; et al. Exploring the chemical safety of fly larvae as a source of protein for animal feed. J. Insects Food Feed 2015, 1, 7–16. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Potential and challenges of insects as an innovative source for food and feed production. Innov. Food Sci. Emerg. Technol. 2013, 17, 1–11. [Google Scholar] [CrossRef]
- REGULATION (EU) 2015/2283 OF THE EUROPEAN PARLIAMENT AND OF THE COUNCIL of 25 November 2015 on Novel Foods, Amending Regulation (EU) No. 1169/2011 of the European Parliament and of the Council and Repealing Regulation (EC) No. 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No. 1852/2001. Available online: www.eur-lex.europa.eu/legal-content/en/TXT/?uri=CELEX%3A32015R2283 (accessed on 10 June 2019).
- Van Raamsdonk, L.W.D.; van der Fels-Klerx, H.J.; de Jong, J. New feed ingredients: the insect opportunity. Food Addit. Contam. Part A 2017, 34, 1384–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- COMMISSION REGULATION (EU) 2017/893 of 24 May 2017 Amending Annexes I and IV to Regulation (EC) No. 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No. 142/2011 as Regards the Provisions on Processed Animal Protein. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32017R0893 (accessed on 10 June 2019).
- REGULATION (EC) No. 999/2001 OF THE EUROPEAN PARLIAMENT AND OF THE COUNCIL of 22 May 2001 Laying down Rules for the Prevention, Control and Eradication of Certain Transmissible Spongiform Encephalopathies. Available online: www.eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32001R0999 (accessed on 10 June 2019).
- EFSA Scientific Committee. Scientific Opinion on a Risk Profile Related to Production and Consumption of Insects as Food and Feed. Available online: www.efsa.europa.eu/it/efsajournal/pub/4257 (accessed on 10 June 2019).
- European Commission (EC). Council Directive (EC) 2002/32/ EC of 7 May 2002 on Undesirable Substances in Animal Feed. Off. J. Eur. Union 2002, L140, 10–21. Available online: www.eur-lex.europa.eu/legal-content/en/ALL/?uri=CELEX:32002L0032 (accessed on 10 June 2019).
- COMMISSION RECOMMENDATION 2006/576/EC of 17 August 2006 on the Presence of Deoxynivalenol, Zearalenone, Ochratoxin A, T-2 and HT-2 and Fumonisins in Products Intended for Animal Feeding. Off. J. Eur. Union 2019, L229, 7–9. Available online: www.eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2006:229:0007:0009:EN:PDF (accessed on 10 June 2019).
- Hussein, H.S.; Brasel, J.M. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef]
- Steinkellner, H.; Binaglia, M.; Dall’Asta, C.; Gutleb, A.C.; Metzler, M.; Oswald, I.P.; Parent-Massin, D.; Alexander, J. Combined hazard assessment of mycotoxins and their modified forms applying relative potency factors: Zearalenone and T2/HT2 toxin. Food Chem. Toxicol. 2019, 131, 110599. [Google Scholar] [CrossRef]
- Binder, E.M.; Tan, L.M.; Chin, L.J.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Tech. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Berthiller, F.; Krska, R.; Domig, K.J.; Kneifel, W.; Juge, N.; Schuhmacher, R.; Adam, G. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 2011, 206, 264–267. [Google Scholar] [CrossRef] [Green Version]
- De Zutter, N.; Audenaert, K.; Arroyo-Manzanares, N.; De Boevre, M.; Van Poucke, C.; De Saeger, S.; Haesaert, G.; Smagghe, G. Aphids transform and detoxify the mycotoxin deoxynivalenol via a type II biotransformation mechanism yet unknown in animals. Sci. Rep. 2016, 6, 38640. [Google Scholar] [CrossRef] [Green Version]
- Van Broekhoven, S.; Gutierrez, J.M.; De Rijk, T.C.; De Nijs, W.C.M.; Van Loon, J.J.A. Degradation and excretion of the Fusarium toxin deoxynivalenol by an edible insect, the Yellow mealworm (Tenebrio molitor L.). World Mycotoxin J. 2017, 10, 163–169. [Google Scholar] [CrossRef]
- Niermans, K.; Woyzichovski, J.; Kröncke, N.; Benning, R.; Maul, R. Feeding study for the mycotoxin zearalenone in yellow mealworm (Tenebrio molitor) larvae—Investigation of biological impact and metabolic conversion. Mycotoxin Res. 2019, 35, 231–242. [Google Scholar] [CrossRef]
- Camenzuli, L.; Van Dam, R.; de Rijk, T.; Andriessen, R.; Van Schelt, J.; Van der Fels-Klerx, H.J. Tolerance and Excretion of the Mycotoxins Aflatoxin B1, Zearalenone, Deoxynivalenol, and Ochratoxin A by Alphitobius diaperinus and Hermetia illucens from contaminated substrates. Toxins 2018, 10, 91. [Google Scholar] [CrossRef]
- Purschke, B.; Scheibelberger, R.; Axmann, S.; Adler, A.; Jäger, H. Impact of substrate contamination with mycotoxins, heavy metals and pesticides on the growth performance and composition of black soldier fly larvae (Hermetia illucens) for use in the feed and food value chain. Food Addit. Contam. Part A. 2017, 34, 1410–1420. [Google Scholar] [CrossRef]
- Sanabria, C.O.; Hogan, N.; Madder, K.; Gillott, C.; Blakley, B.; Reaney, M.; Beattie, A.; Buchanan, F. Yellow Mealworm Larvae (Tenebrio molitor) Fed Mycotoxin-Contaminated Wheat—A Possible Safe, Sustainable Protein Source for Animal Feed? Toxins 2019, 11, 282. [Google Scholar] [CrossRef]
- Dall’Erta, A.; Cirlini, M.; Dall’Asta, M.; Del Rio, D.; Galaverna, G.; Dall’Asta, G. Masked Mycotoxins Are Efficiently Hydrolyzed by Human Colonic Microbiota Releasing Their Aglycones. Chem. Res. Toxicol. 2013, 26, 305–312. [Google Scholar] [CrossRef]
- Gratz, S.W.; Dinesh, R.; Yoshinari, T.; Holtrop, G.; Richardson, A.J.; Duncan, G.; MacDonald, G.; Lloyd, A.; Tarbin, J. Masked trichothecene and zearalenone mycotoxins withstand digestion and absorption in the upper GI tract but are efficiently hydrolyzed by human gut microbiota in vitro. Mol. Nutr. Food Res. 2017, 61, 1600680. [Google Scholar] [CrossRef]
- Rychlik, M.; Humpf, H.U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef]
- Damiani, T.; Righetti, L.; Suman, M.; Galaverna, G.; Dall’Asta, C. Analytical issue related to fumonisins: A matter of sample comminution? Food Control. 2019, 95, 1–5. [Google Scholar] [CrossRef]
- Leni, G.; Soetemans, L.; Jacobs, J.; Depraetere, S.; Gianotten, N.; Bastiaens, L.; Caligiani, A.; Sforza, S. Protein hydrolysates from Alphitobius diaperinus and Hermetia illucens larvae treated with commercial proteases. Food Res. Int. 2019. under review. [Google Scholar]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef]
- Dall’Asta, C.; Galaverna, G.; Aureli, G.; Dossena, A.; Marchelli, R. LC/MS/MS method for the simultaneous quantification of free and masked fumonisins in maize and maize-based products. World Mycotoxin J. 2008, 1, 237–246. [Google Scholar] [CrossRef]


{kind=link}
| Substrate Samples | Mycotoxin Amount (µg/kg) | ||||
|---|---|---|---|---|---|
| Sample Code | Description | DON | FB1 | FB2 | ZEN |
| WM | Wheat middlings | 938 ± 100 | <LOD | <LOD | <LOD |
| CDR | Corn distillation residues | 779 ± 5 | 573 ± 3 | 441 ± 3 | <LOD |
| CG | Corn gluten feed | 1207 ± 43 | 727 ± 6 | 294 ± 5 | 173 ± 4 |
| RB | Rice Bran | <LOD | <LOD | <LOD | <LOD |
| RW | Rapeseed wastes | <LOD | <LOD | <LOD | <LOD |
| OP | Olive pomace | LOD | <LOD | <LOD | <LOD |
| AP | Apple pomace | <LOD | <LOD | <LOD | <LOD |
| CC | Chopped carrots | <LOD | <LOD | <LOD | <LOD |
| Larvae Samples | Mycotoxin Amount (µg/kg) | ||||
|---|---|---|---|---|---|
| Sample Code | Description | DON | FB1 | FB2 | ZEN |
| BSF-100% CDR | BSF larvae grown on: 100% CDR | <LOD | <LOD | <LOD | <LOD |
| BSF-79% CDR-10.5% OP/AP | BSF larvae grown on: 79% CDR, 10.5% OP, 10.5% AP | <LOD | <LOD | <LOD | <LOD |
| LM-100% WM | LM larvae grown on: 100% WM, 0% CG | 416 ± 28 | <LOD | <LOD | <LOD |
| LM-75% WM-25% CG | LM larvae grown on: 75% WM, 25% CG | 608 ± 59 | <LOD | <LOD | <LOD |
| LM-50% WM-50% CG | LM larvae grown on: 50% WM, 50% CG | <LOD | <LOD | <LOD | <LOD |
| LM-100% CG | LM larvae grown on: 100% CG | 726 ± 164 | 127 ± 6 | <LOD | <LOD |
| LM-100% CDR * | LM larvae grown on: 100% CDR * | 468 ± 181 | <LOD | <LOD | <LOD |
| LM-95% WM-5% RB | LM larvae grown on: 95% WM, 5% RB | <LOD | <LOD | <LOD | <LOD |
| LM-90% WM-10% RB | LM larvae grown on: 90% WM, 10% RB | 755 ± 134 | <LOD | <LOD | <LOD |
| LM-85% WM-15% RB | LM larvae grown on: 85% WM, 15% RB | <LOD | <LOD | <LOD | <LOD |
| LM-80% WM-20% RB | LM larvae grown on: 80% WM, 20% RB | <LOD | <LOD | <LOD | <LOD |
| LM-95% WM-5% RW | LM larvae grown on: 95% WM, 5% RW | <LOD | <LOD | <LOD | <LOD |
| LM-90% WM-10% RW | LM larvae grown on: 90% WM, 10% RW | <LOD | <LOD | <LOD | <LOD |
| LM-85% WM-15% RW | LM larvae grown on: 85% WM, 15% RW | 557 ± 237 | <LOD | <LOD | <LOD |
| LM-80% WM-20% RW | LM larvae grown on: 80% WM, 20% RW | <LOD | <LOD | <LOD | <LOD |
| Residual Fraction Samples | Mycotoxin Amount (µg/kg) | ||||
|---|---|---|---|---|---|
| Sample Code | Description | DON | FB1 | FB2 | ZEN |
| REST-BSF-100% CDR | Rests of BSF larvae grown on: 100% CDR | 1473 ± 197 | 951 ± 152 | 344 ± 64 | 334 ± 44 |
| REST-BSF-79% CDR-10.5% OP/AP | Rests of BSF larvae grown on: 79% CDR, 10.5% OP, 10.5% AP | <LOD | <LOD | <LOD | <LOD |
| REST-LM-100% WM | Rests of LM larvae grown on: 100% WM, 0% CG | <LOD | <LOD | <LOD | <LOD |
| REST-LM-75% WM-25% CG | LM larvae grown on: 75% WM, 25% CG | <LOD | <LOD | <LOD | <LOD |
| REST-LM-100% CG | Rests of LM larvae grown on: 100% CG | 827 ± 61 | 728 ± 7 | <LOD | <LOD |
| REST-LM-100% CDR * | Rests of LM larvae grown on: 100% CDR * | 587 ± 73 | 224 ± 8 | <LOD | <LOD |
| REST-LM-90% WM-10% RB | Rests of LM larvae grown on: 90% WM, 10% RB | <LOD | <LOD | <LOD | <LOD |
| REST-LM-85% WM-15% RW | Rests of LM larvae grown on: 85% WM, 15% RW | <LOD | <LOD | <LOD | <LOD |
| Compound | Ionization Mode | Precursor Ion (m/z) | Product Ions (m/z) | Collision Energy (V) | LOD (µg/kg) |
|---|---|---|---|---|---|
| PAT | Negative | 152.9 [M − H]− | 109/81 | −12/−12 | 100 |
| NIV | Negative | 371.1 [M + CH3COO]− | 311.1/281.1/59.1 | −10/−32/−48 | 10 |
| DON | Negative | 355.1 [M + CH3COO]− | 295.1/265.1 | −13/−16 | 10 |
| 3ADON | Negative | 397.1 [M + CH3COO]− | 307.1/59 | −18/−20 | 20 |
| FUSX | Negative | 413.3 [M + CH3COO]− | 353.6/262.9/59.1 | −14/−22/−10 | 20 |
| OTA | Positive | 404.5 [M + H]+ | 238.7/220.7/101.7 | 21/31/68 | 20 |
| FB1 | Positive | 722.3 [M + H]+ | 704.7/352.1/334.1 | 26/35/38 | 25 |
| FB2 | Positive | 706.5 [M + H]+ | 688.4/336.3 | 51/51 | 25 |
| DAS | Positive | 384.2 [M + NH4]+ | 307.2/105.1 | 17/61 | 10 |
| T2 | Positive | 484.3 [M + NH4]+ | 215.0/185.0 | 19/22 | 10 |
| HT2 | Positive | 442.0 [M + NH4]+ | 263.1 | 11 | 10 |
| ZEN | Negative | 317.0 [M − H]− | 175.0/131.0 | −26/−32 | 10 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leni, G.; Cirlini, M.; Jacobs, J.; Depraetere, S.; Gianotten, N.; Sforza, S.; Dall’Asta, C. Impact of Naturally Contaminated Substrates on Alphitobius diaperinus and Hermetia illucens: Uptake and Excretion of Mycotoxins. Toxins 2019, 11, 476. https://doi.org/10.3390/toxins11080476
Leni G, Cirlini M, Jacobs J, Depraetere S, Gianotten N, Sforza S, Dall’Asta C. Impact of Naturally Contaminated Substrates on Alphitobius diaperinus and Hermetia illucens: Uptake and Excretion of Mycotoxins. Toxins. 2019; 11(8):476. https://doi.org/10.3390/toxins11080476
Chicago/Turabian StyleLeni, Giulia, Martina Cirlini, Johan Jacobs, Stefaan Depraetere, Natasja Gianotten, Stefano Sforza, and Chiara Dall’Asta. 2019. "Impact of Naturally Contaminated Substrates on Alphitobius diaperinus and Hermetia illucens: Uptake and Excretion of Mycotoxins" Toxins 11, no. 8: 476. https://doi.org/10.3390/toxins11080476
APA StyleLeni, G., Cirlini, M., Jacobs, J., Depraetere, S., Gianotten, N., Sforza, S., & Dall’Asta, C. (2019). Impact of Naturally Contaminated Substrates on Alphitobius diaperinus and Hermetia illucens: Uptake and Excretion of Mycotoxins. Toxins, 11(8), 476. https://doi.org/10.3390/toxins11080476