Venomic, Transcriptomic, and Bioactivity Analyses of Pamphobeteus verdolaga Venom Reveal Complex Disulfide-Rich Peptides That Modulate Calcium Channels

 ,
,  , , , ,
, , , ,
Abstract
:
1. Introduction
2. Results
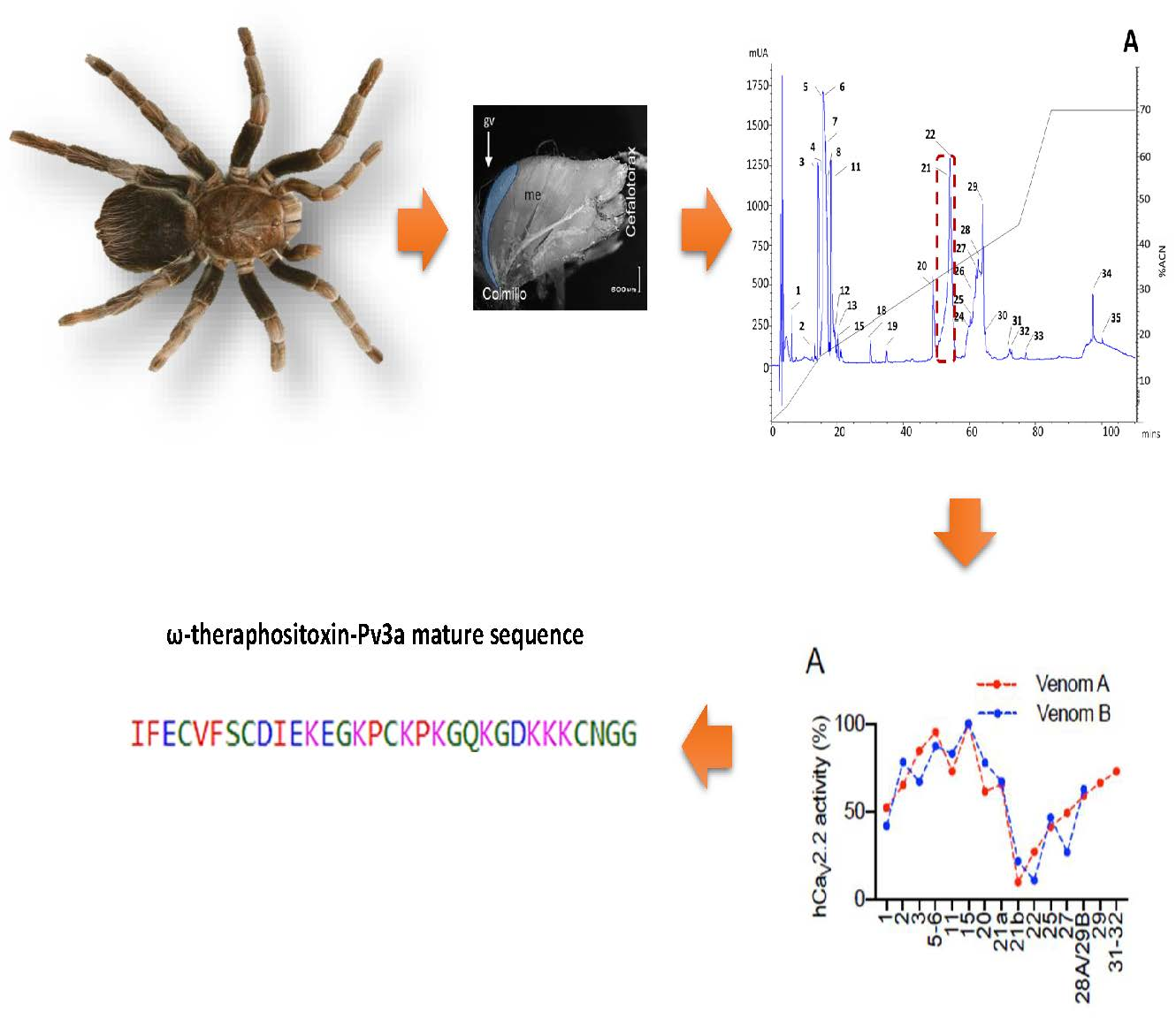
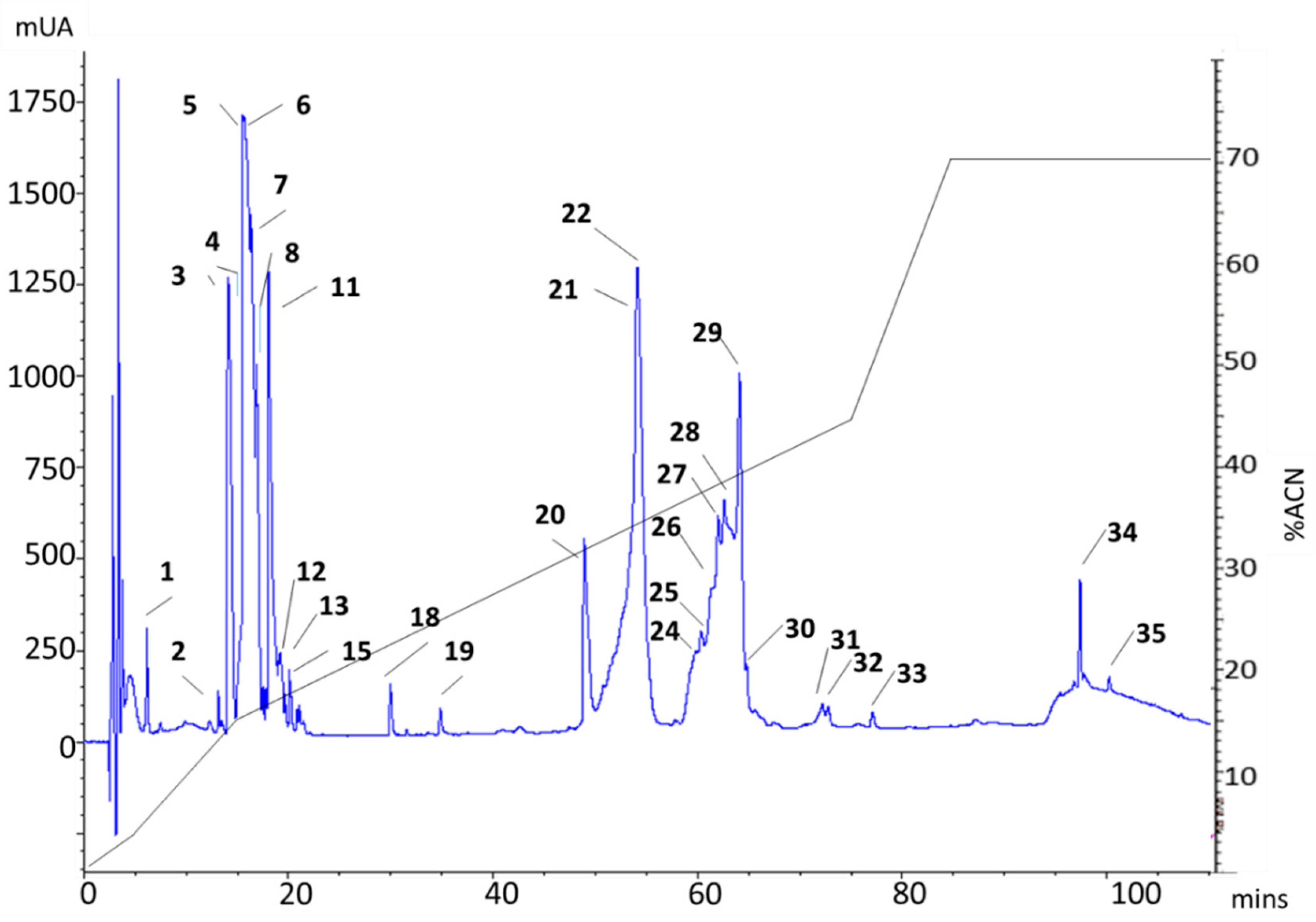
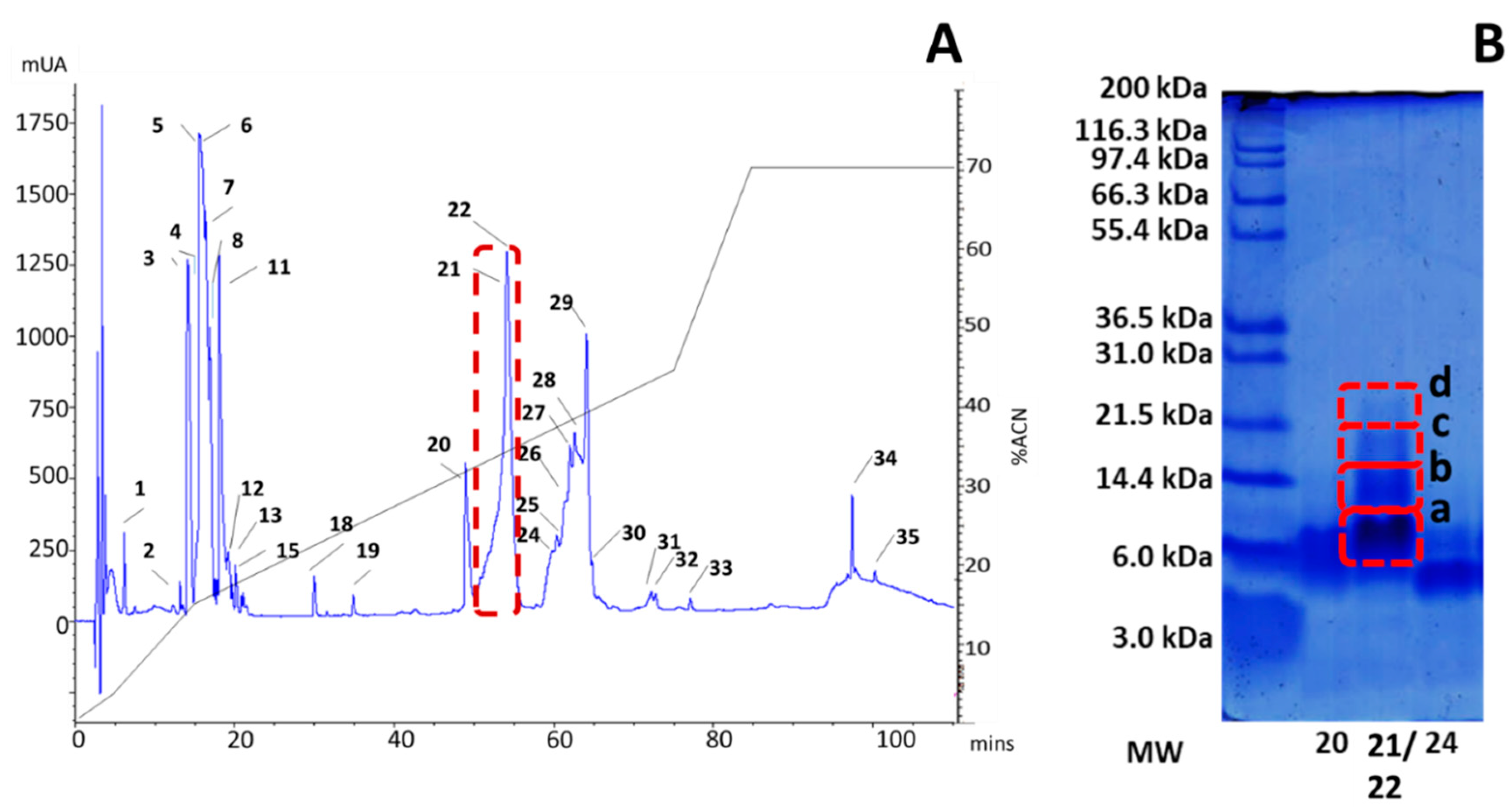
2.1. rp-HPLC Profile
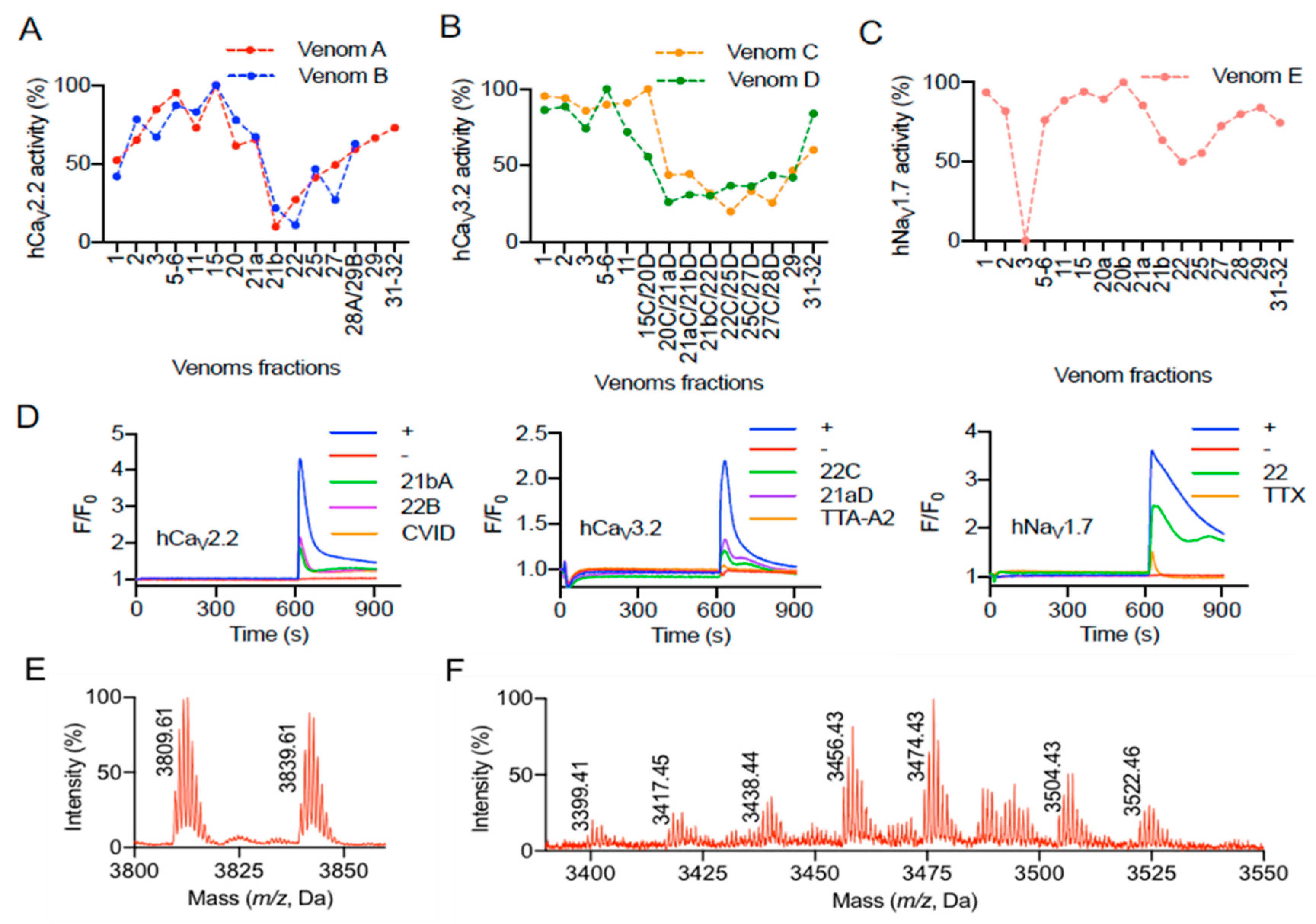
2.2. Bioactivity of Pamphobeteus verdolaga Venom
2.3. Tandem Mass Spectrometry (MS)/MS Analysis of the Active Fraction
2.4. Transcriptomic Results
2.4.1. General Transcriptomic Assembly Analysis
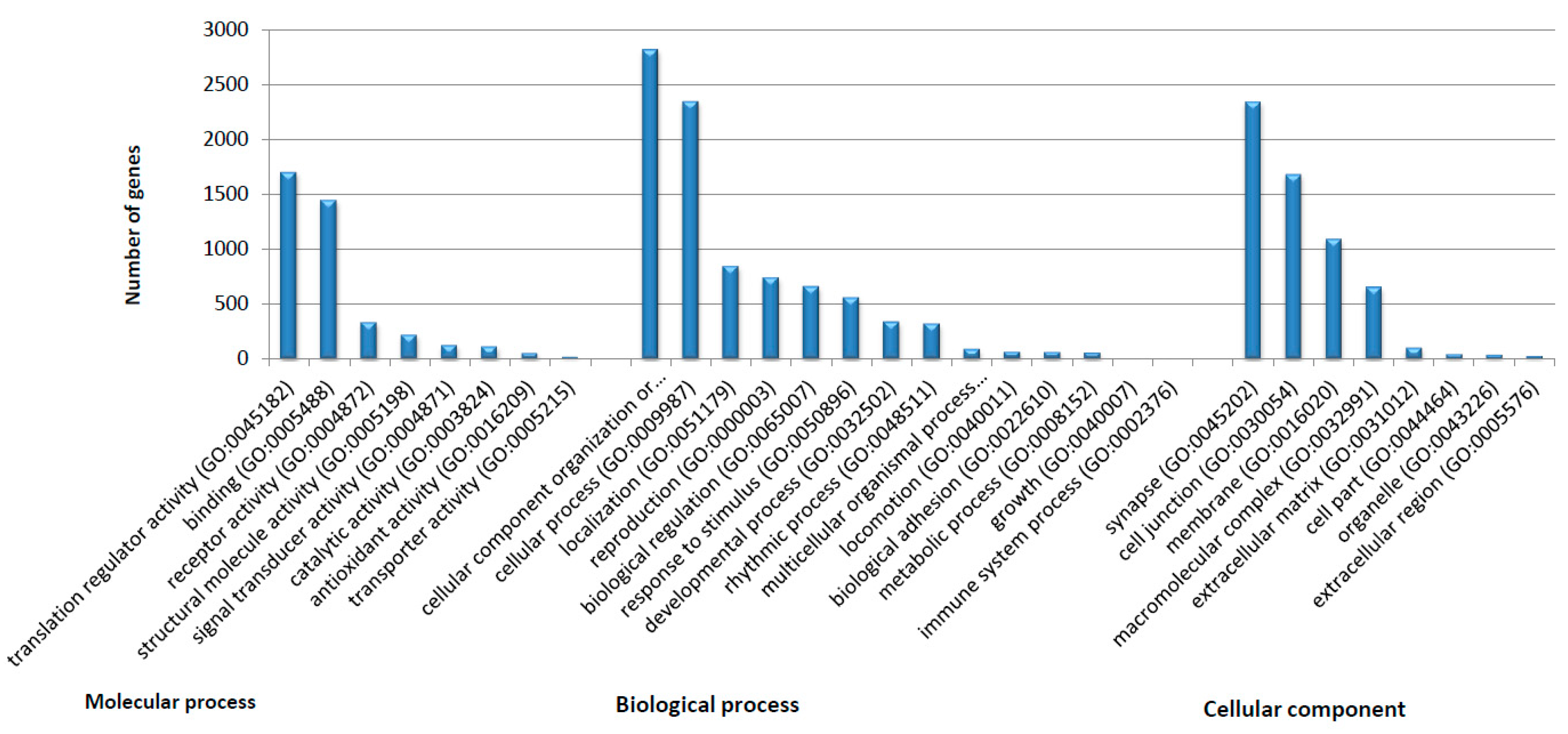
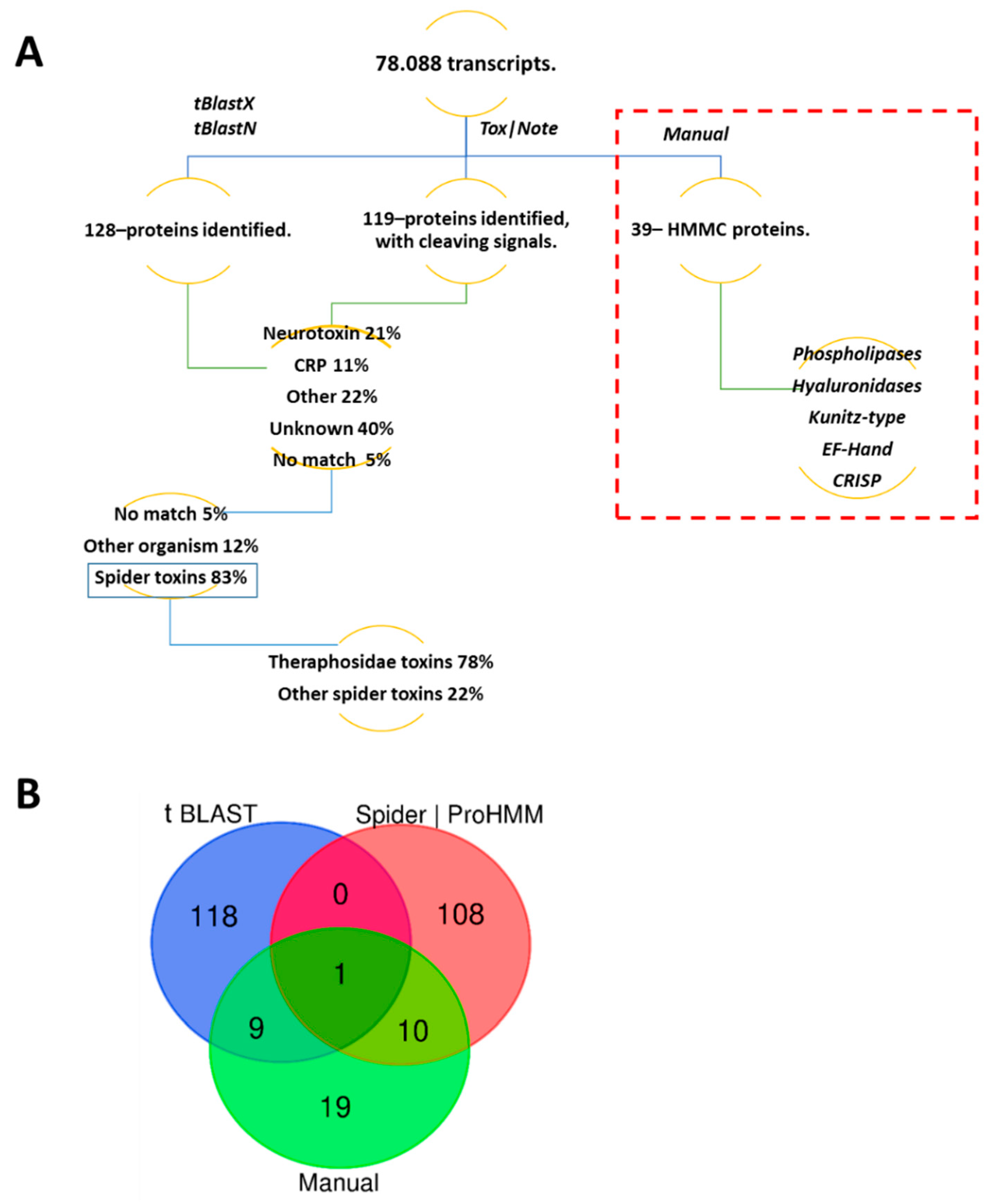
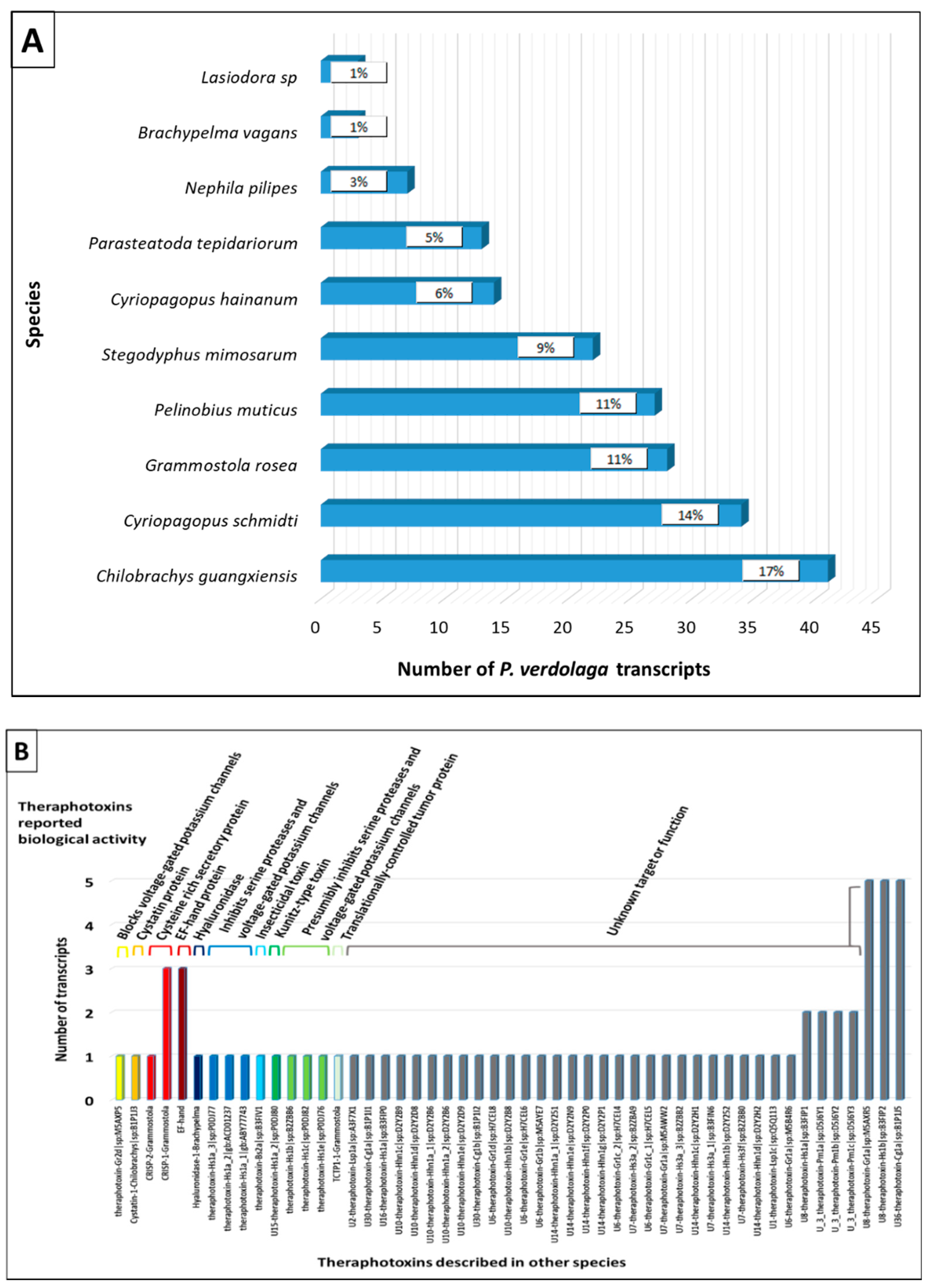
2.4.2. Transcriptomic Annotation
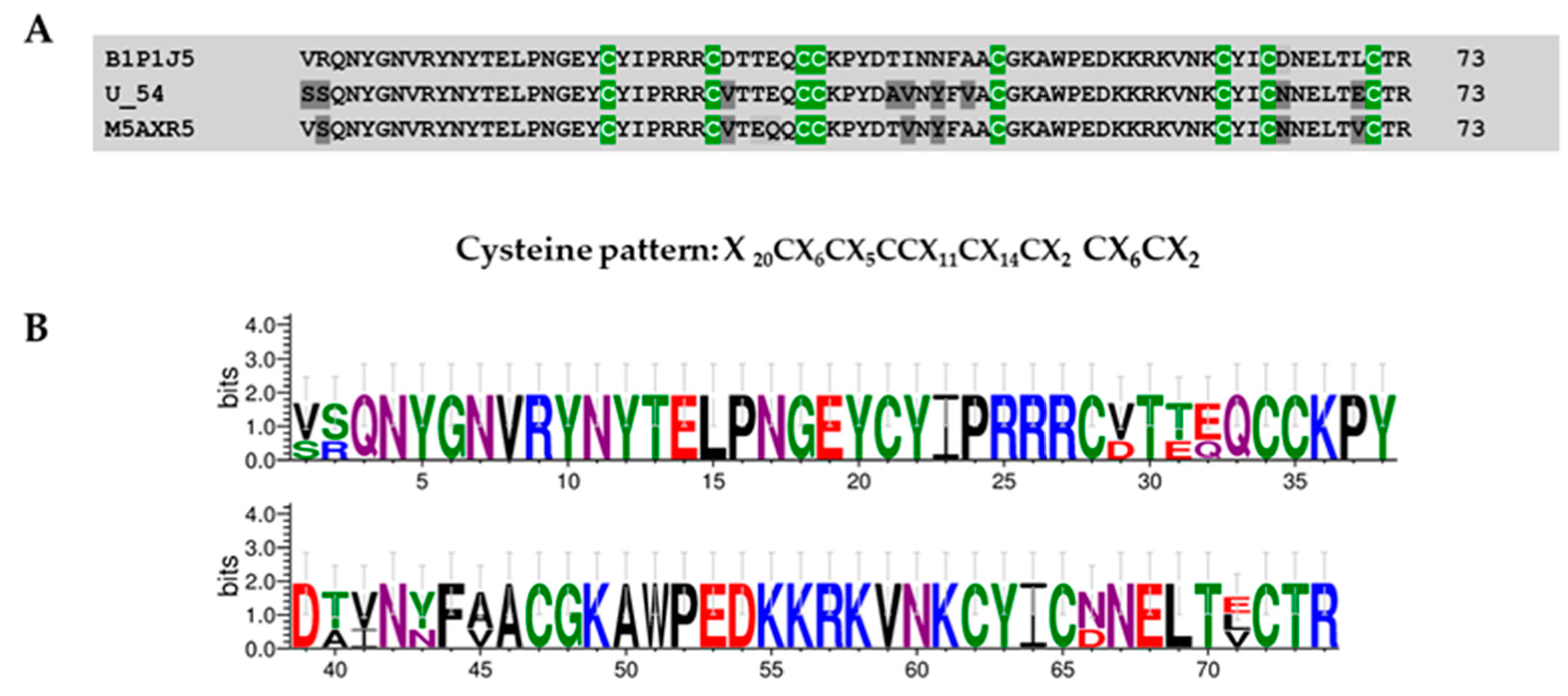
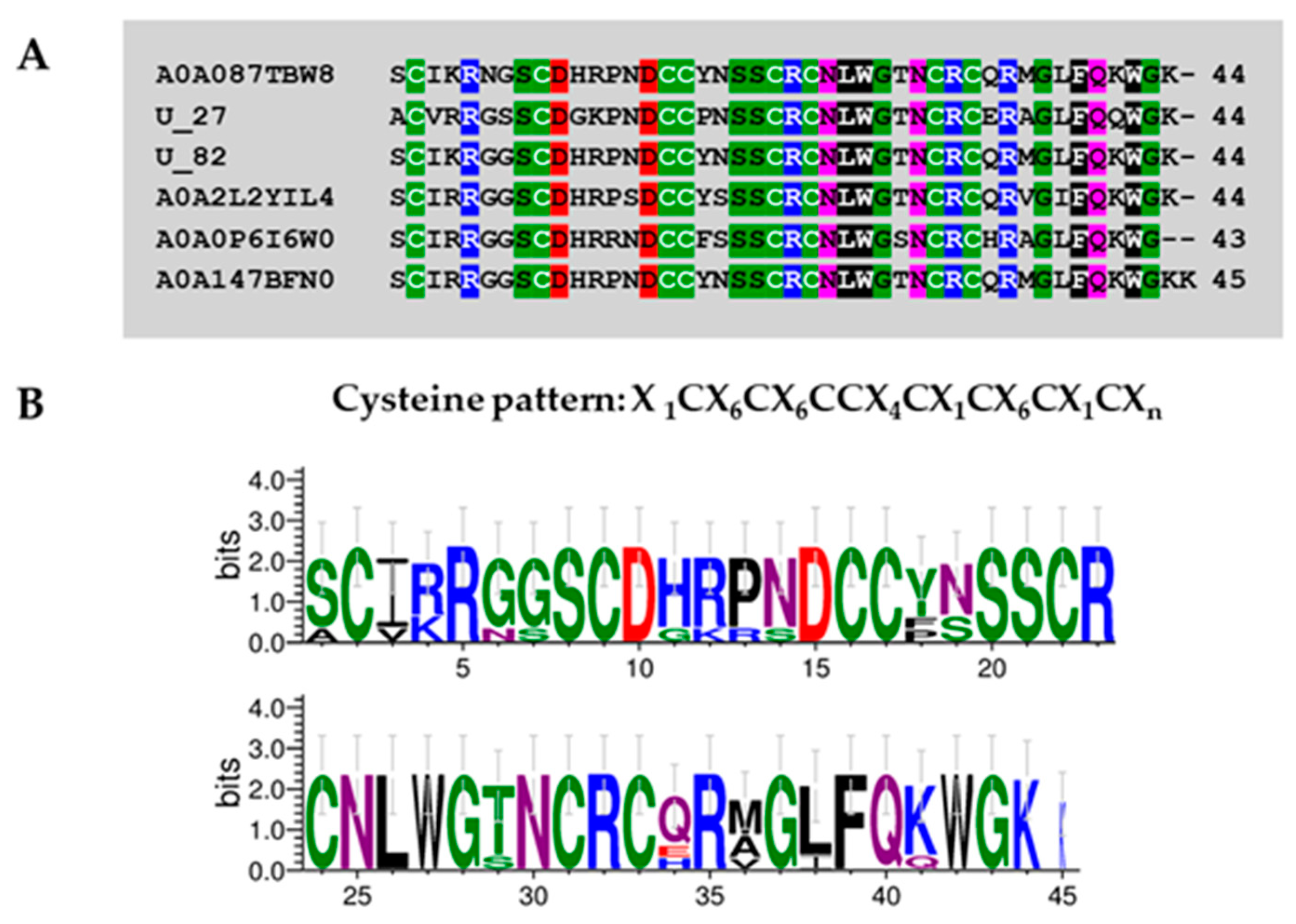
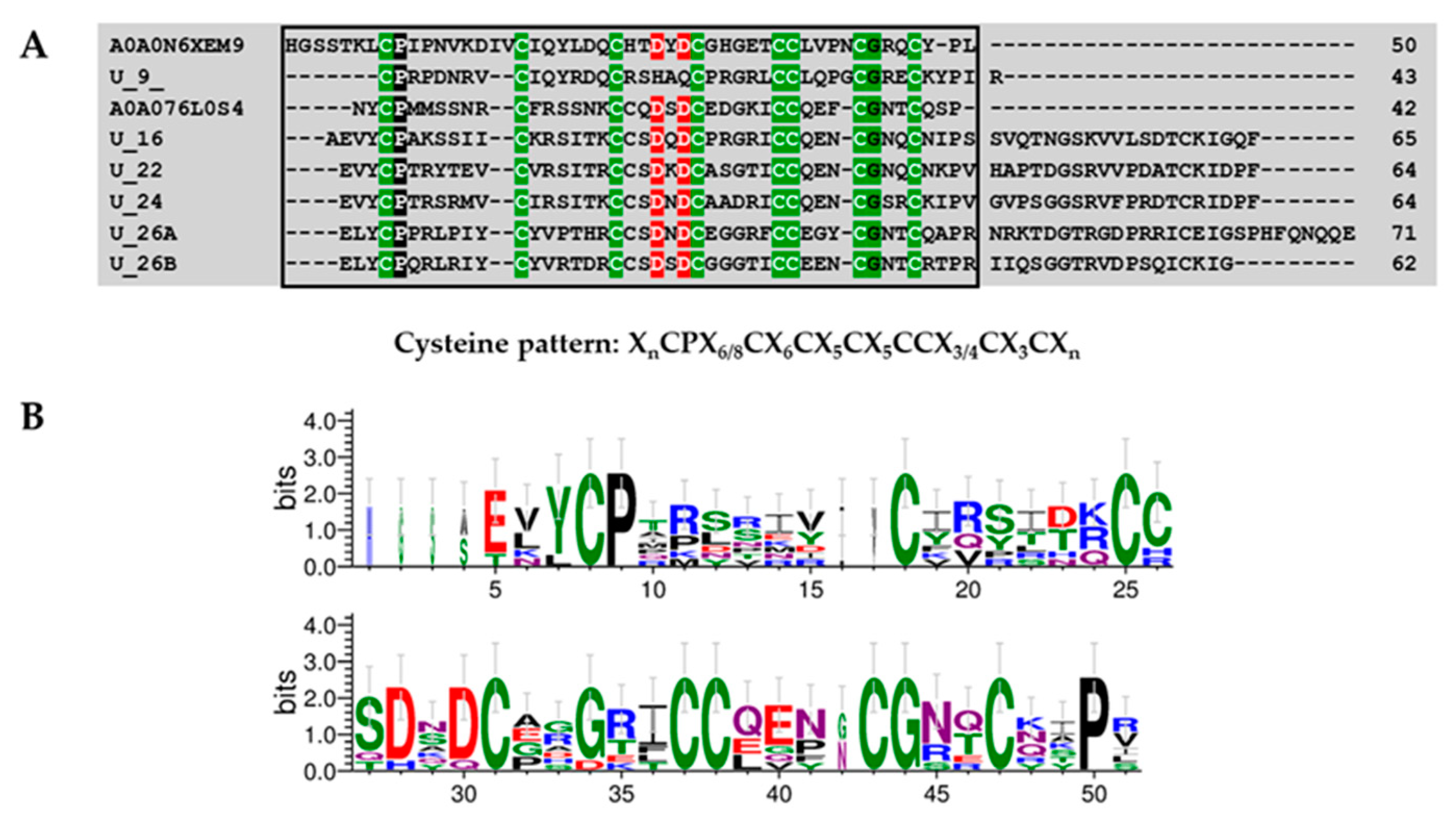
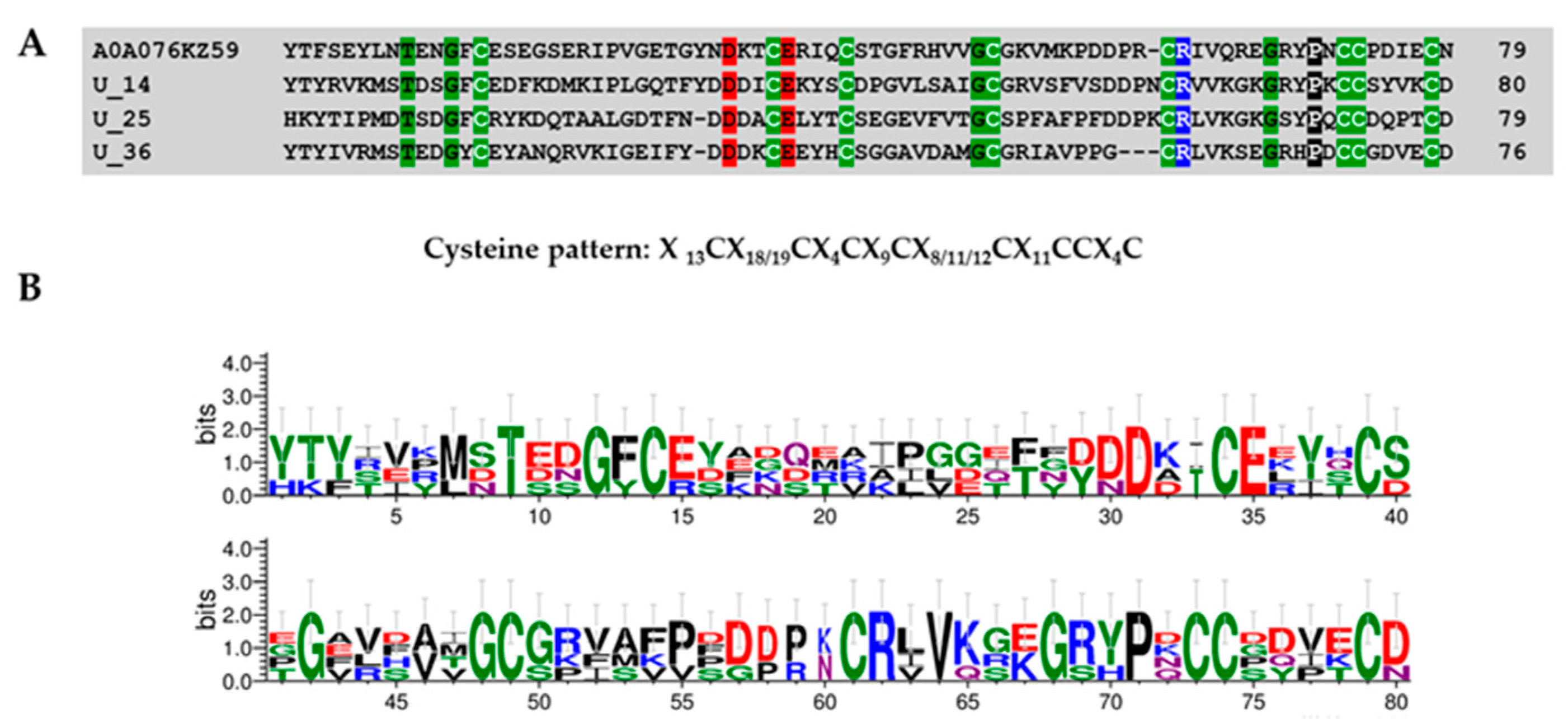
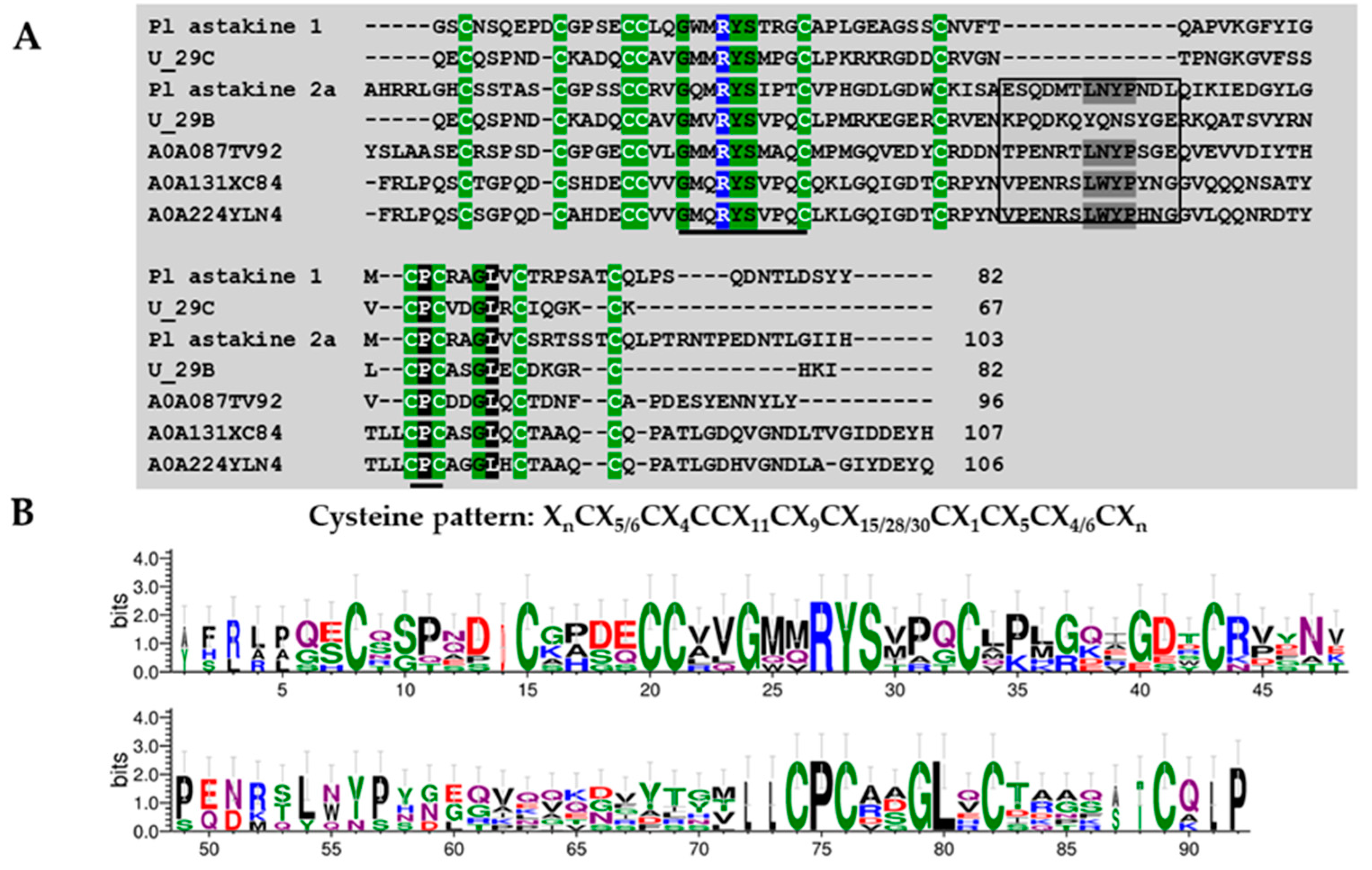
Superfamily 1
Superfamily 2
Superfamily 3
Superfamily 4
Superfamily 5
Superfamily 6
Superfamily 7
Superfamily 8
2.4.3. Transcriptomics Reveal Sequences of Potential Voltage-Gated Calcium Channel Modulators
3. Discussion
4. Materials and Methods
4.1. Spider Collection and Venom Extraction
4.2. Venom Fractionation
4.3. Sample Preparation for Proteomic Analysis
4.4. Data Analysis
4.5. LC-MS/MS
4.6. Venom Gland RNA Extraction and Library Construction
4.7. De Novo Transcriptome Assembly and Gene Annotation
4.8. Nomenclature
4.9. Protein Domain Searching and Sequence Logos
4.10. Disulfide Bond Prediction
4.11. Calcium Influx Assays
4.12. Data Deposition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Escoubas, P.; Rash, L. Tarantulas: Eight-legged pharmacists and combinatorial chemists. Toxicon 2004, 43, 555–574. [Google Scholar] [CrossRef] [PubMed]
- King, J.B.; Gross, J.; Lovly, C.M.; Piwnica-Worms, H.; Townsend, R.R. Identification of protein phosphorylation sites within ser/thr-rich cluster domains using site-directed mutagenesis and hybrid linear quadrupole ion trap fourier transform ion cyclotron resonance mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 3443–3451. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, G.M. Spider venom peptides. In Handbook of Biologically Active Peptides; Hastin, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Nicholson, G.M. Insect-selective spider toxins targeting voltage-gated sodium channels. Toxicon 2007, 49, 490–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassilevski, A.A.; Kozlov, S.A.; Grishin, E.V. Molecular diversity of spider venom. Biochemistry (Moscow) 2009, 74, 1505–1534. [Google Scholar] [CrossRef] [PubMed]
- Estrada, G.S.; Vargas, M.L.; Ramirez, A.; Quintana, C.J. Milking and partial characterization Pamphobeteus spp (aranae; theraphosidae) venom, from the colombian andean region. Toxicon 2012, 60, 231–232. [Google Scholar]
- Rash, L.D.; Hodgson, W.C. Pharmacology and biochemistry of spider venoms. Toxicon 2002, 40, 225–254. [Google Scholar] [CrossRef]
- King, G.F. Modulation of insect Ca(v) channels by peptidic spider toxins. Toxicon 2007, 49, 513–530. [Google Scholar] [CrossRef]
- King, G.F.; Hardy, M.C. Spider-venom peptides: Structure, pharmacology, and potential for control of insect pests. Ann. Rev. Entomol. 2013, 58, 475–496. [Google Scholar] [CrossRef]
- Pineda, S.S.; Undheim, E.A.; Rupasinghe, D.B.; Ikonomopoulou, M.P.; King, G.F. Spider venomics: Implications for drug discovery. Future Med. Chem. 2014, 6, 1699–1714. [Google Scholar] [CrossRef]
- Abreu, T.F.; Sumitomo, B.N.; Nishiyama, M.Y., Jr.; Oliveira, U.C.; Souza, G.H.; Kitano, E.S.; Zelanis, A.; Serrano, S.M.; Junqueira-de-Azevedo, I.; Silva, P.I., Jr.; et al. Peptidomics of Acanthoscurria gomesiana spider venom reveals new toxins with potential antimicrobial activity. J. Proteom. 2017, 151, 232–242. [Google Scholar] [CrossRef]
- Chen, J.; Deng, M.; He, Q.; Meng, E.; Jiang, L.; Liao, Z.; Rong, M.; Liang, S. Molecular diversity and evolution of cystine knot toxins of the tarantula Chilobrachys jingzhao. Cell. Mol. Life Sci. 2008, 65, 2431–2444. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhao, L.; Jiang, L.; Meng, E.; Zhang, Y.; Xiong, X.; Liang, S. Transcriptome analysis revealed novel possible venom components and cellular processes of the tarantula Chilobrachys jingzhao venom gland. Toxicon 2008, 52, 794–806. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.C.; Long, R.W.; Wu, Y.Q.; Guo, Y.B.; Liu, D.L.; Peng, L.; Li, D.Q.; Yang, D.W.; Xu, X.; Liu, F.X.; et al. Identification and characterization of toxins in the venom gland of the chinese bird spider, Haplopelma hainanum, by transcriptomic analysis. Insect Sci. 2016, 23, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.C.; Dekan, Z.; Rosengren, K.J.; Erickson, A.; Vetter, I.; Deuis, J.R.; Herzig, V.; Alewood, P.F.; King, G.F.; Lewis, R.J. Identification and characterization of protx-iii [u-trtx-tp1a], a new voltage-gated sodium channel inhibitor from venom of the tarantula Thrixopelma pruriens. Mol. Pharm. 2015, 88, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.C.; Dekan, Z.; Smith, J.J.; Deuis, J.R.; Vetter, I.; Herzig, V.; Alewood, P.F.; King, G.F.; Lewis, R.J. Modulatory features of the novel spider toxin μ-trtx-df1a isolated from the venom of the spider Davus fasciatus. Br. J. Pharm. 2017, 174, 2528–2544. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Gomez, S.; Gomez-Rave, L.; Vargas-Munoz, L.J.; van der Meijden, A. Characterizing the biological and biochemical profile of six different scorpion venoms from the buthidae and scorpionidae family. Toxicon 2017, 130, 104–115. [Google Scholar] [CrossRef]
- Estrada-Gomez, S.; Vargas Munoz, L.J.; Quintana Castillo, J.C. Extraction and partial characterization of venom from the colombian spider Pamphobeteus aff. Nigricolor (aranae:Theraphosidae). Toxicon 2013, 76, 301–309. [Google Scholar] [CrossRef]
- Cardoso, F.C.; Lewis, R.J. Sodium channels and pain: From toxins to therapies. Br. J. Pharmacol. 2018, 175, 2138–2157. [Google Scholar] [CrossRef]
- Xiong, X.F.; Poulsen, M.H.; Hussein, R.A.; Norager, N.G.; Stromgaard, K. Structure-activity relationship study of spider polyamine toxins as inhibitors of ionotropic glutamate receptors. ChemMedChem 2014, 9, 2661–2670. [Google Scholar] [CrossRef]
- WorldSpiderCatalog. World Spider Catalog. Bern, N.H.M., Ed.; 2019. Volume Version 20.0. Available online: http://wsc.nmbe.ch (accessed on 01 July 2019).
- Pineda, S.S.; Chaumeil, P.A.; Kunert, A.; Kaas, Q.; Thang, M.W.C.; Le, L.; Nuhn, M.; Herzig, V.; Saez, N.J.; Cristofori-Armstrong, B.; et al. Arachnoserver 3.0: An online resource for automated discovery, analysis and annotation of spider toxins. Bioinformatics 2018, 34, 1074–1076. [Google Scholar] [CrossRef]
- Savel-Niemann, A.; Roth, D. Biochemical analysis of tarantula venom (Eurypelma californicum). Naturwissenschaften 1989, 76, 212–213. [Google Scholar] [CrossRef] [PubMed]
- Nason, D.M.; Phillips, D.; Saccomano, N.A.; Volkmann, R.A. Calcium Channel Blocking Polypeptides from Therpahosidae Aphonopelma. U.S. Patent WO1994010196 A, 11 May 1994. [Google Scholar]
- Corzo, G.; Diego-Garcia, E.; Clement, H.; Peigneur, S.; Odell, G.; Tytgat, J.; Possani, L.D.; Alagon, A. An insecticidal peptide from the theraposid Brachypelma smithi spider venom reveals common molecular features among spider species from different genera. Peptides 2008, 29, 1901–1908. [Google Scholar] [CrossRef] [PubMed]
- Liang, S. An overview of peptide toxins from the venom of the chinese bird spider Selenocosmia huwena wang [=Ornithoctonus huwena (wang)]. Toxicon 2004, 43, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.; Daly, N.L. Venomics: A mini-review. High. Throughput 2018, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhang, Y.; Hu, W.; Xu, D.; Tao, H.; Yang, X.; Li, Y.; Jiang, L.; Liang, S. Molecular diversification of peptide toxins from the tarantula Haplopelma hainanum (Ornithoctonus hainana) venom based on transcriptomic, peptidomic, and genomic analyses. J. Proteome Res. 2010, 9, 2550–2564. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes, Y.; Estrada-Gomez, S.; Vargas Munoz, L.J.; Perafan, C. Description and molecular characterization of a new species of tarantula, Pamphobeteus verdolaga, from colombia (aranae: Mygalomorphae: Theraphosidae). Zoologia 2016, 33. [Google Scholar] [CrossRef]
- Escoubas, P.; Sollod, B.; King, G.F. Venom landscapes: Mining the complexity of spider venoms via a combined cdna and mass spectrometric approach. Toxicon 2006, 47, 650–663. [Google Scholar] [CrossRef]
- Leao, R.M.; Cruz, J.S.; Diniz, C.R.; Cordeiro, M.N.; Beirao, P.S. Inhibition of neuronal high-voltage activated calcium channels by the omega-phoneutria nigriventer tx3-3 peptide toxin. Neuropharmacology 2000, 39, 1756–1767. [Google Scholar] [CrossRef]
- Dos Santos, R.G.; Van Renterghem, C.; Martin-Moutot, N.; Mansuelle, P.; Cordeiro, M.N.; Diniz, C.R.; Mori, Y.; De Lima, M.E.; Seagar, M. Phoneutria nigriventer omega-phonetoxin iia blocks the cav2 family of calcium channels and interacts with omega-conotoxin-binding sites. J. Biol. Chem. 2002, 277, 13856–13862. [Google Scholar] [CrossRef]
- Vieira, L.B.; Kushmerick, C.; Hildebrand, M.E.; Garcia, E.; Stea, A.; Cordeiro, M.N.; Richardson, M.; Gomez, M.V.; Snutch, T.P. Inhibition of high voltage-activated calcium channels by spider toxin pntx3-6. J. Pharm. Exp. 2005, 314, 1370–1377. [Google Scholar] [CrossRef]
- Vieira, L.B.; Pimenta, A.M.; Richardson, M.; Bemquerer, M.P.; Reis, H.J.; Cruz, J.S.; Gomez, M.V.; Santoro, M.M.; Ferreira-de-Oliveira, R.; Figueiredo, S.G.; et al. Leftward shift in the voltage-dependence for Ca2+ currents activation induced by a new toxin from Phoneutria reidyi (aranae, Ctenidae) venom. Cell. Mol. Neurobiol. 2007, 27, 129–146. [Google Scholar] [CrossRef]
- Newcomb, R.; Palma, A.; Fox, J.; Gaur, S.; Lau, K.; Chung, D.; Cong, R.; Bell, J.R.; Horne, B.; Nadasdi, L.; et al. Snx-325, a novel calcium antagonist from the spider Segestria florentina. Biochemistry 1995, 34, 8341–8347. [Google Scholar] [CrossRef]
- Ikonomopoulou, M.P.; Smith, J.J.; Herzig, V.; Pineda, S.S.; Dziemborowicz, S.; Er, S.Y.; Durek, T.; Gilchrist, J.; Alewood, P.F.; Nicholson, G.M.; et al. Isolation of two insecticidal toxins from venom of the australian theraphosid spider Coremiocnemis tropix. Toxicon Off. J. Int. Soc. Toxinol. 2016, 123, 62–70. [Google Scholar] [CrossRef]
- Jiang, L.; Peng, L.; Chen, J.; Zhang, Y.; Xiong, X.; Liang, S. Molecular diversification based on analysis of expressed sequence tags from the venom glands of the chinese bird spider Ornithoctonus huwena. Toxicon 2008, 51, 1479–1489. [Google Scholar] [CrossRef]
- Lin, X.; Novotny, M.; Söderhäll, K.; Söderhäll, I. Ancient cytokines, the role of astakines as hematopoietic growth factors. J. Biol. Chem. 2010, 285, 28577–28586. [Google Scholar] [CrossRef]
- Oukkache, N.; Chgoury, F.; Lalaoui, M.; Cano, A.A.; Ghalim, N. Comparison between two methods of scorpion venom milking in morocco. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 5. [Google Scholar] [CrossRef]
- World Health Organization. Progress in the Characterization of Venoms and Standardization of Antivenoms; WHO Offset Publication: Geneva, Switzerland, 1981; pp. 1–44. [Google Scholar]
- Fernandez, J.; Gutierrez, J.M.; Angulo, Y.; Sanz, L.; Juarez, P.; Calvete, J.J.; Lomonte, B. Isolation of an acidic phospholipase a2 from the venom of the snake Bothrops asper of Costa Rica: Biochemical and toxicological characterization. Biochimie 2010, 92, 273–283. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage t4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Herzig, V.; Wood, D.L.A.; Newell, F.; Chaumeil, P.-A.; Kaas, Q.; Binford, G.J.; Nicholson, G.M.; Gorse, D.; King, G.F. Arachnoserver 2.0, an updated online resource for spider toxin sequences and structures. Nucl. Acids Res. 2011, 39, D653–D657. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from rna-seq using the trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Simao, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. Busco: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. Rsem: Accurate transcript quantification from rna-seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. Signalp 4.1: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Goujon, M.; McWilliam, H.; Li, W.; Valentin, F.; Squizzato, S.; Paern, J.; Lopez, R. A new bioinformatics analysis tools framework at embl-ebi. Nucl. Acids Res. 2010, 38, W695–W699. [Google Scholar] [CrossRef]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Nicholas, K.B.; Nicholas, H.B.; Deerfield, D.W. Genedoc: Analysis and visualization of genetic variation. Embnew News 1997, 4, 14. [Google Scholar]
- Ceroni, A.; Passerini, A.; Vullo, A.; Frasconi, P. DISULFIND: A Disulfide Bonding State and Cysteine Connectivity Prediction Server. Nucleic Acids Res. 2006, 34 (Suppl. S2), W177–W181. [Google Scholar] [CrossRef]
- Sairaman, A.; Cardoso, F.C.; Bispat, A.; Lewis, R.J.; Duggan, P.J.; Tuck, K.L. Synthesis and evaluation of aminobenzothiazoles as blockers of n- and t-type calcium channels. Bioorg. Med. Chem. 2018, 26, 3046–3059. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| rp-HPLC Fraction Number/Tris-Tricine Band | Mapp (kDa) | m/z | z | Peptide Sequence | Score | Best Transcriptomic Match | TPM |
|---|---|---|---|---|---|---|---|
| 21-22/a-b_1 | 6–10▼ | 444.36 | 2 | K.IKLCLKI.- | 40% | c7142_g1_i1 | 27806.19 |
| 21-22/a-b_2 | 773.98 | 2 | -.IFECVFSCDIEK.E | 100% | c7142_g1_i1 | 27806.19 | |
| 21-22/a-b_3 | 618.64 | 4 | -.IFECVFSCDIEKEGKPCKPK.G | 100% | c7142_g1_i1 | 27806.19 | |
| 21-22/c_1 | 10▼ | 579.10 | 2 | CVFSCDXEK | 81.9% | c7142_g1_i1 | 27806.19 |
| Superfamily | Cysteine Pattern | Number of Cysteines | Mature Peptide Length |
|---|---|---|---|
| SF1 | C-C-CC-C | 5 | 54 |
| SF2 | C-C-C-CC-C | 6 | 48 |
| SF3 | C-C-CC-C-C-C | 7 | 43–84 |
| SF4 | C-C-CC-C-C-C-C | 8 | 44–74 |
| SF5 | C-C-C-CC-C-C-C | 8 | 61–65 |
| SF6 | C-C-C-C-CC-C-C | 8 | 43–71 |
| SF7 | C-C-C-C-C-CC-C | 8 | 76–80 |
| SF8 | C-C-CC-C-C-C-C-C-C | 10 | 67–88 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estrada-Gomez, S.; Cardoso, F.C.; Vargas-Muñoz, L.J.; Quintana-Castillo, J.C.; Arenas Gómez, C.M.; Pineda, S.S.; Saldarriaga-Cordoba, M.M. Venomic, Transcriptomic, and Bioactivity Analyses of Pamphobeteus verdolaga Venom Reveal Complex Disulfide-Rich Peptides That Modulate Calcium Channels. Toxins 2019, 11, 496. https://doi.org/10.3390/toxins11090496
Estrada-Gomez S, Cardoso FC, Vargas-Muñoz LJ, Quintana-Castillo JC, Arenas Gómez CM, Pineda SS, Saldarriaga-Cordoba MM. Venomic, Transcriptomic, and Bioactivity Analyses of Pamphobeteus verdolaga Venom Reveal Complex Disulfide-Rich Peptides That Modulate Calcium Channels. Toxins. 2019; 11(9):496. https://doi.org/10.3390/toxins11090496
Chicago/Turabian StyleEstrada-Gomez, Sebastian, Fernanda Caldas Cardoso, Leidy Johana Vargas-Muñoz, Juan Carlos Quintana-Castillo, Claudia Marcela Arenas Gómez, Sandy Steffany Pineda, and Monica Maria Saldarriaga-Cordoba. 2019. "Venomic, Transcriptomic, and Bioactivity Analyses of Pamphobeteus verdolaga Venom Reveal Complex Disulfide-Rich Peptides That Modulate Calcium Channels" Toxins 11, no. 9: 496. https://doi.org/10.3390/toxins11090496
APA StyleEstrada-Gomez, S., Cardoso, F. C., Vargas-Muñoz, L. J., Quintana-Castillo, J. C., Arenas Gómez, C. M., Pineda, S. S., & Saldarriaga-Cordoba, M. M. (2019). Venomic, Transcriptomic, and Bioactivity Analyses of Pamphobeteus verdolaga Venom Reveal Complex Disulfide-Rich Peptides That Modulate Calcium Channels. Toxins, 11(9), 496. https://doi.org/10.3390/toxins11090496