The Histone Deacetylases HosA and HdaA Affect the Phenotype and Transcriptomic and Metabolic Profiles of Aspergillus niger

Abstract
:1. Introduction
2. Results
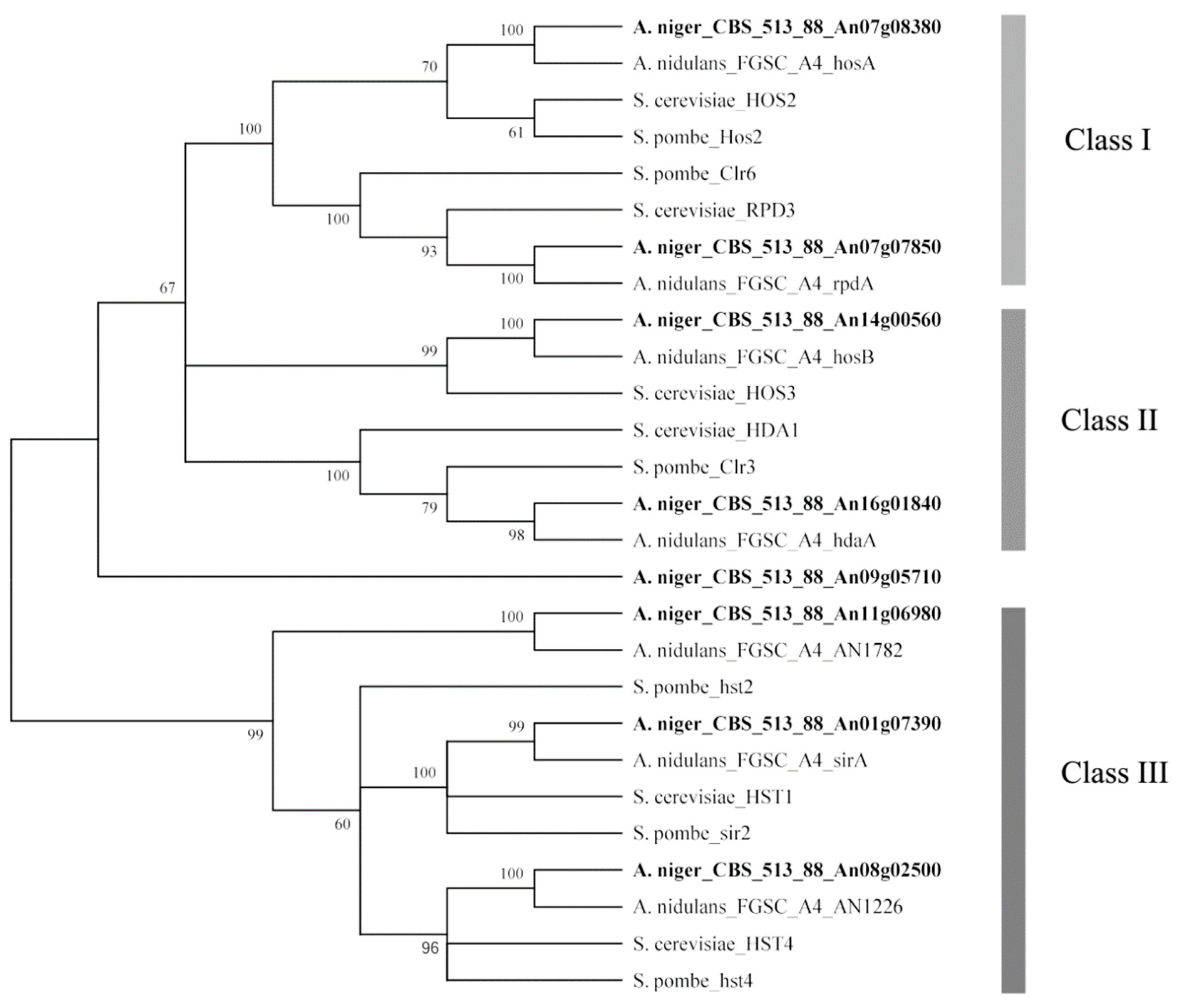
2.1. Phylogenetic Analysis Predicted the Amino Acid Sequences of HDACs in A. niger FGSC A1279 and Related Genes
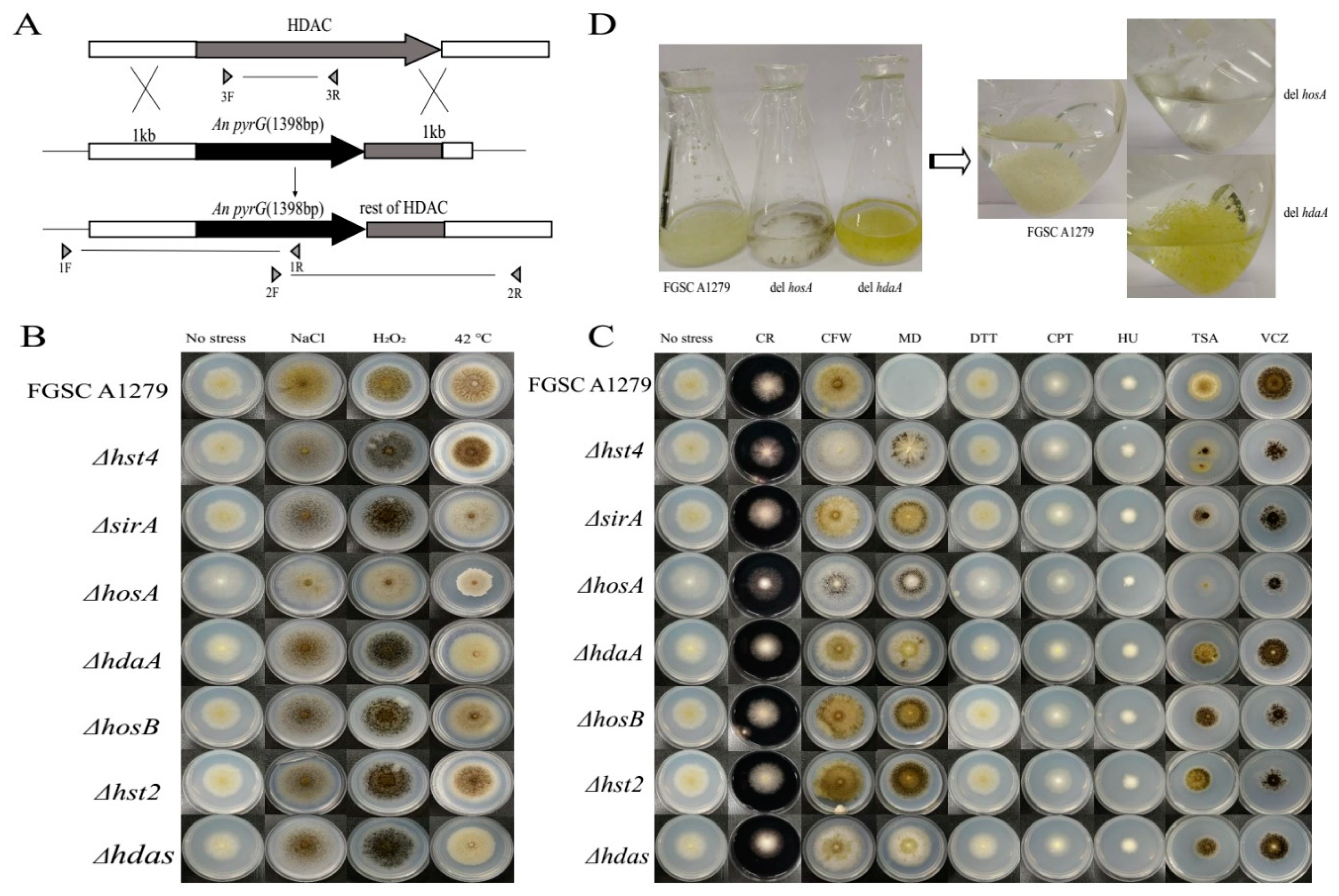
2.2. HDAC-Deficient Strain Construction and Phenotype Analysis
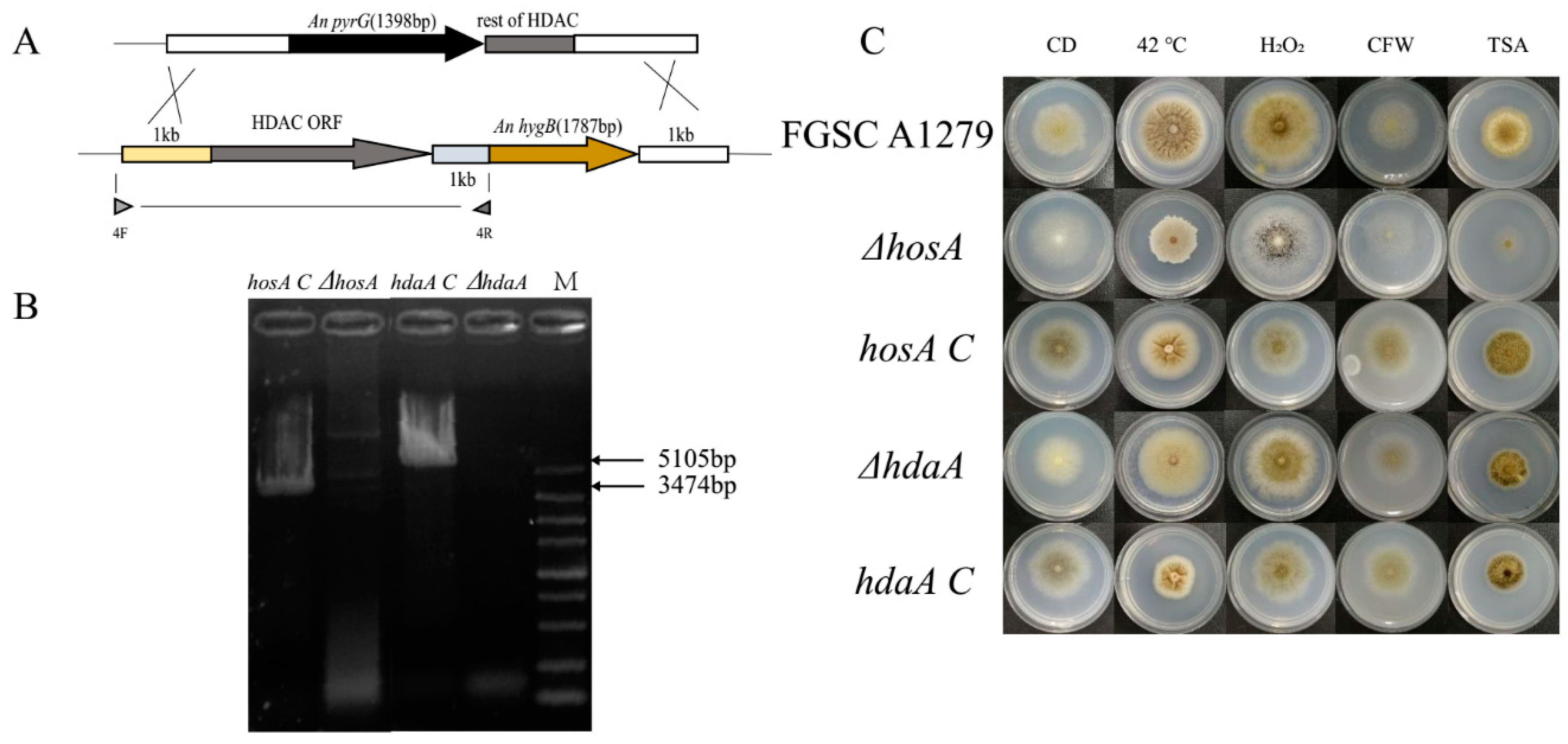
2.3. Phenotype Analysis of HosA and HdaA Complementation Strains
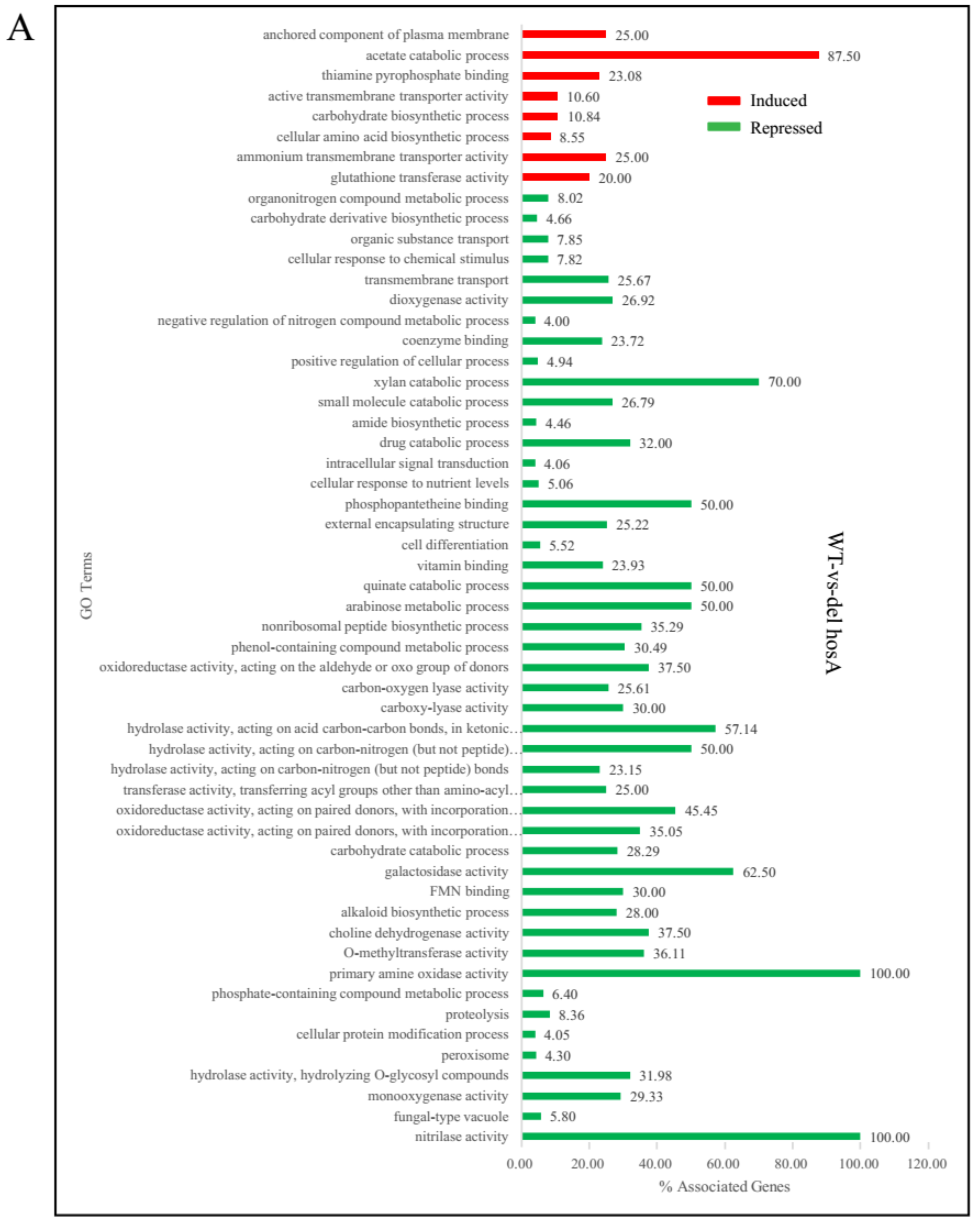
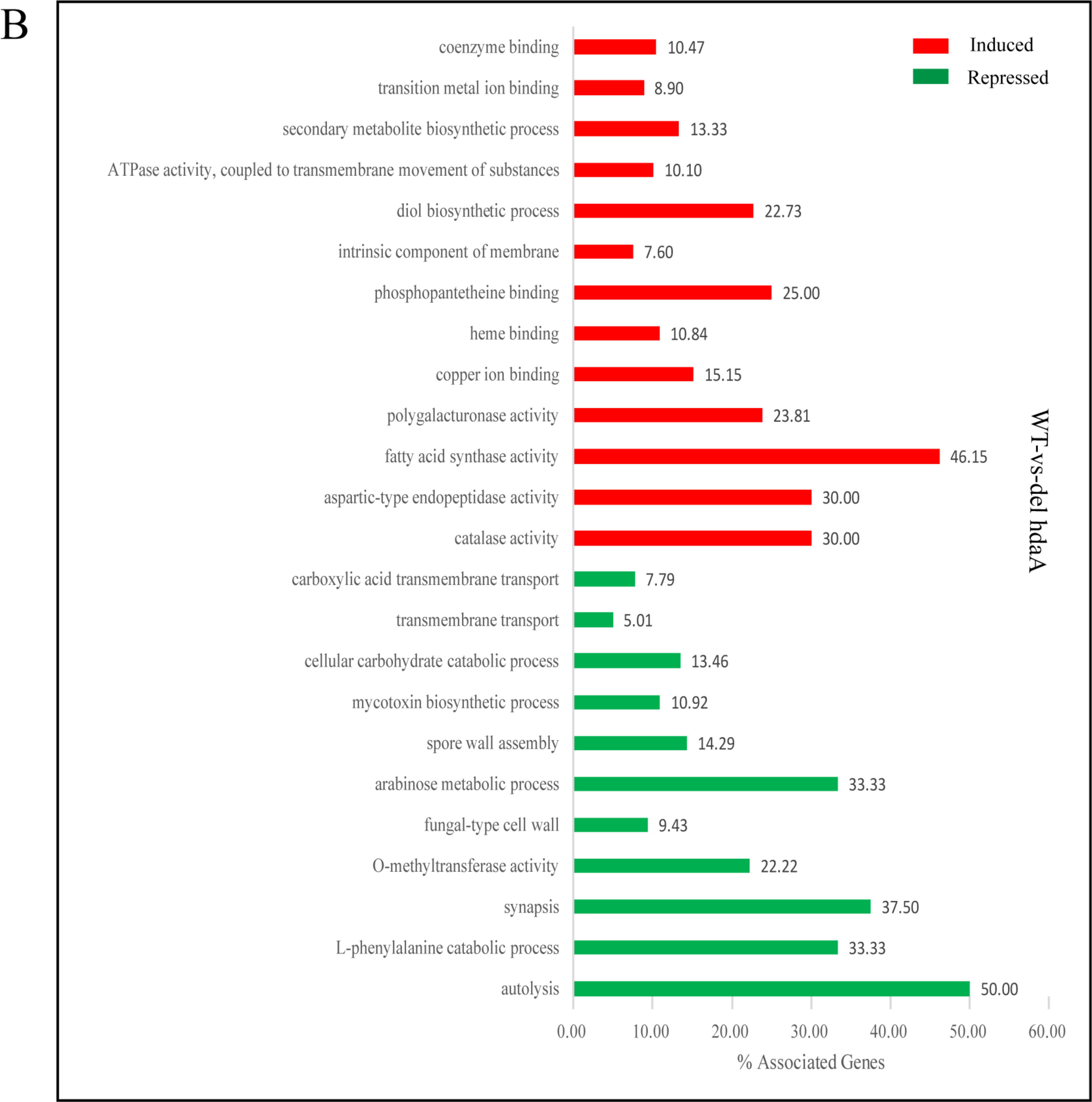
2.4. Transcriptomic Profiles of A. niger FGSC A1279, ΔHosA and ΔHdaA
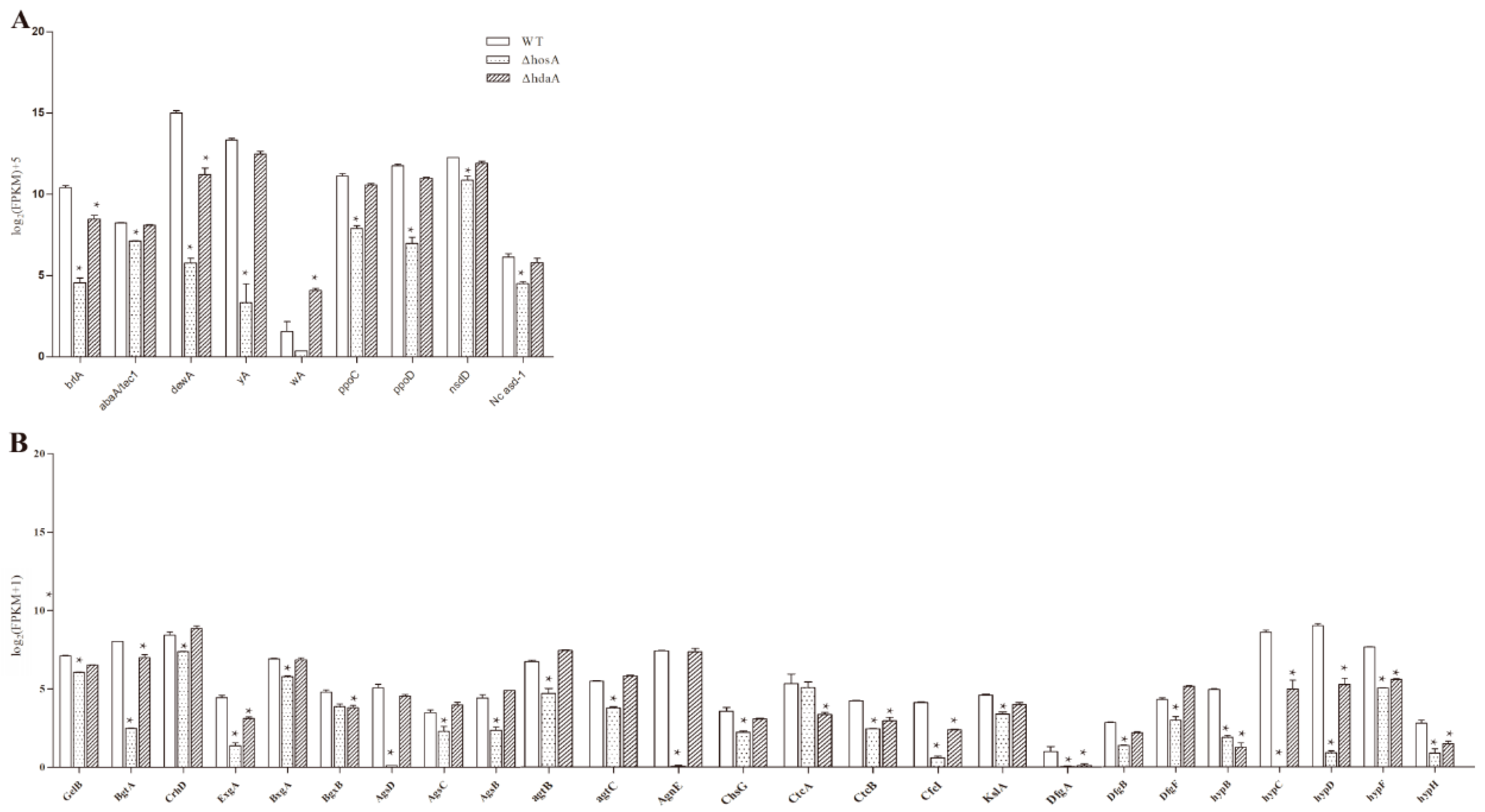
2.5. Effect of HosA and HdaA on Asexual or Sexual Reproduction, Pigmentation, and Cell Wall Synthesis
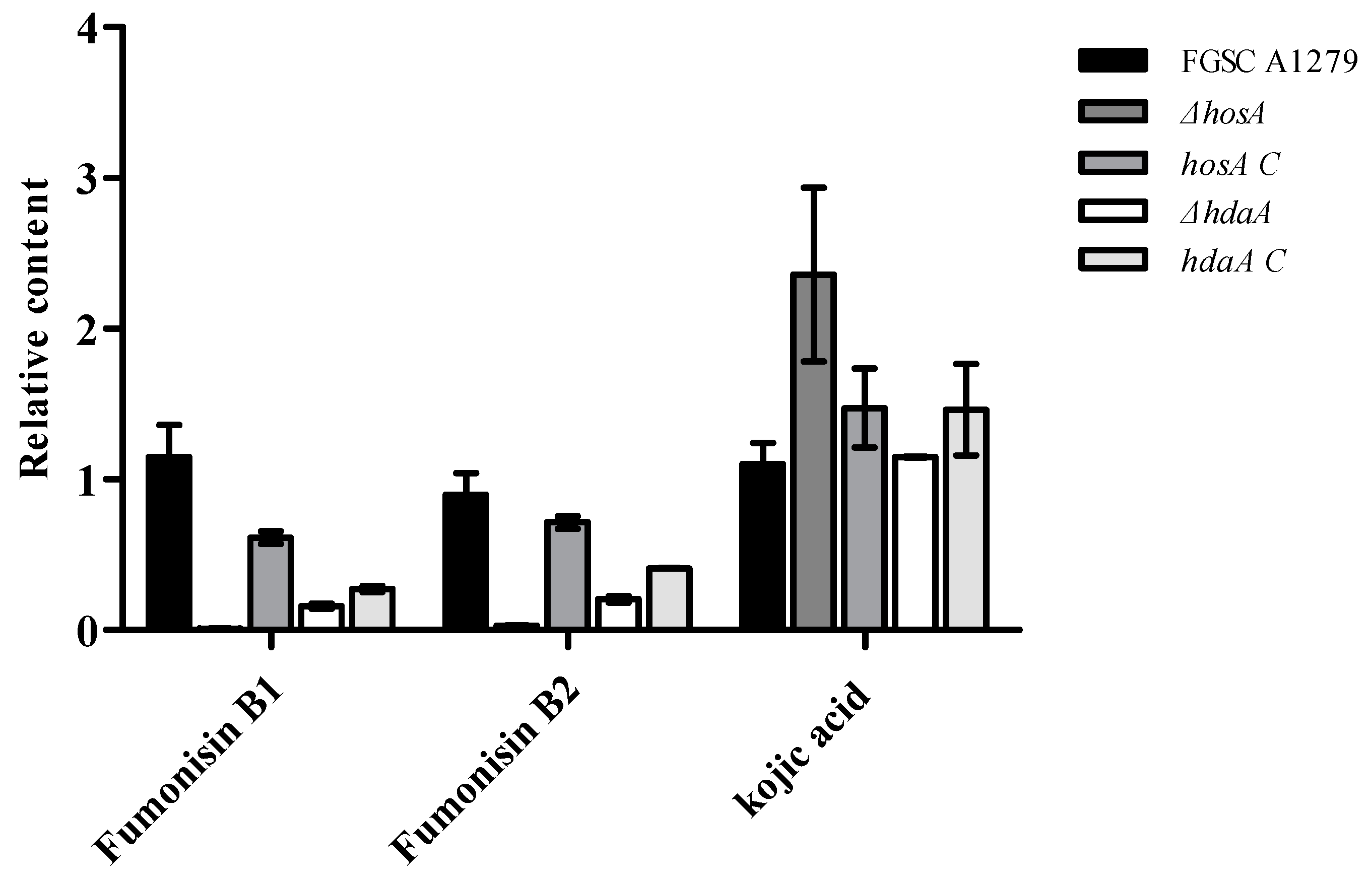
2.6. Effects of the Genes HosA and HdaA on Secondary Metabolites
3. Discussion
4. Materials and Methods
4.1. Phylogenetic Analysis of HDACs
4.2. Strains and Growth Conditions
4.3. Construction of A. niger Histone Deacetylase Mutants
4.4. Environmental Stress Resistance and Drug Resistance Assay
4.5. RNA Preparation and RNA-Seq Analysis
4.6. Secondary Metabolite Collection and Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Verdin, E.; Ott, M. 50 years of protein acetylation: From gene regulation to epigenetics, metabolism and beyond. Nat. Rev. Mol. Cell Biol. 2015, 16, 258–264. [Google Scholar] [CrossRef] [PubMed]
- West, A.C.; Johnstone, R.W. New and emerging HDAC inhibitors for cancer treatment. J. Clin. Investig. 2014, 124, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurdistani, S.K.; Grunstein, M. Histone acetylation and deacetylation in yeast. Nat. Rev. Mol. Cell Biol. 2003, 4, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, M.; Iwashita, K. Functional analysis of histone deacetylase and its role in stress response, drug resistance and solid-state cultivation in Aspergillus oryzae. J. Biosci. Bioeng. 2014, 118, 172–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawauchi, M.; Nishiura, M.; Iwashita, K. Fungus-specific sirtuin HstD coordinates secondary metabolism and development through control of LaeA. Eukaryot. Cell 2013, 12, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Shwab, E.K.; Bok, J.W.; Tribus, M.; Galehr, J.; Graessle, S.; Keller, N.P. Histone deacetylase activity regulates chemical diversity in Aspergillus. Eukaryot. Cell 2007, 6, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Oh, J.-H.; Shwab, E.K.; Dagenais, T.R.T.; Andes, D.; Keller, N.P. HdaA, a class 2 histone deacetylase of Aspergillus fumigatus, affects germination and secondary metabolite production. Fungal Genet. Biol. 2009, 46, 782–790. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Izawa, M.; Nakajima, Y.; Jin, Q.; Hirose, T.; Nakamura, T.; Koshino, H.; Kanamaru, K.; Ohsato, S.; Kamakura, T.; et al. Increased metabolite production by deletion of an HDA1-type histone deacetylase in the phytopathogenic fungi, Magnaporthe oryzae (Pyricularia oryzae) and Fusarium asiaticum. Lett. Appl. Microbiol. 2017, 65, 446–452. [Google Scholar] [CrossRef]
- Izawa, M.; Takekawa, O.; Arie, T.; Teraoka, T.; Yoshida, M.; Kimura, M.; Kamakura, T. Inhibition of histone deacetylase causes reduction of appressorium formation in the rice blast fungus Magnaporthe oryzae. J. Gen. Appl. Microbiol. 2009, 55, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, C.; Liu, W.; Wang, G.; Kang, Z.; Kistler, H.C.; Xu, J.-R. The HDF1 Histone Deacetylase Gene Is Important for Conidiation, Sexual Reproduction, and Pathogenesis in Fusarium graminearum. Mol. Plant-Microbe Interact. 2011, 24, 487–496. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Smedsgaard, J.; Samson, R.A.; Larsen, T.O.; Thrane, U. Fumonisin B 2 production by Aspergillus niger. J. Agric. Food Chem. 2007, 55, 9727–9732. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, X.; Yu, D.; Chen, X.; Tabudravu, J.; Deng, H.; Pan, L. Deletion of the epigenetic regulator GcnE in Aspergillus niger FGSC A1279 activates the production of multiple polyketide metabolites. Microbiol. Res. 2018, 217, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.E. Aspergillus niger genomics: Past, present and into the future. Med. Mycol. 2006, 441, S17–S21. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Lv, Y.; Li, X.; Lin, Y.; Deng, H.; Pan, L. Profiling of secondary metabolite gene clusters regulated by LaeA in Aspergillus niger FGSC A1279 based on genome sequencing and transcriptome analysis. Res. Microbiol. 2018, 169, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Graessle, S.; Dangl, M.; Haas, H.; Mair, K.; Trojer, P.; Brandtner, E.M.; Walton, J.D.; Loidl, P.; Brosch, G. Characterization of two putative histone deacetylase genes from Aspergillus nidulans. Biochim. Biophys. Acta-Gene Struct. Expr. 2000, 1492, 120–126. [Google Scholar] [CrossRef]
- Trojer, P.; Brandtner, E.M.; Brosch, G.; Loidl, P.; Galehr, J.; Linzmaier, R.; Haas, H.; Mair, K.; Tribus, M.; Graessle, S. Histone deacetylases in fungi: Novel members, new facts. Nucleic Acids Res. 2003, 31, 3971–3981. [Google Scholar] [CrossRef]
- Brosch, G.; Loidl, P.; Graessle, S. Histone modifications and chromatin dynamics: A focus on filamentous fungi. FEMS Microbiol. Rev. 2008, 32, 409–439. [Google Scholar] [CrossRef]
- Lamoth, F.; Juvvadi, P.R.; Steinbach, W.J. Histone deacetylase inhibition as an alternative strategy against invasive aspergillosis. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Tribus, M.; Bauer, I.; Galehr, J.; Rieser, G.; Trojer, P.; Brosch, G.; Loidl, P.; Haas, H.; Graessle, S. A Novel Motif in Fungal Class 1 Histone Deacetylases Is Essential for Growth and Development of Aspergillus. Mol. Biol. Cell 2010, 21, 345–353. [Google Scholar] [CrossRef]
- Tribus, M.; Galehr, J.; Trojer, P.; Brosch, G.; Loidl, P.; Marx, F.; Haas, H.; Graessle, S. HdaA, a major class 2 histone deacetylase of Aspergillus nidulans, affects growth under conditions of oxidative stress. Eukaryot. Cell 2005, 4, 1736–1745. [Google Scholar] [CrossRef]
- Brandao, F.; Esher, S.K.; Ost, K.S.; Pianalto, K.; Nichols, C.B.; Fernandes, L.; Bocca, A.L.; Pocas-Fonseca, M.J.; Alspaugh, J.A. HDAC genes play distinct and redundant roles in Cryptococcus neoformans virulence. Sci. Rep. 2018, 8, 5209. [Google Scholar] [CrossRef] [PubMed]
- Baidyaroy, D.; Brosch, G.; Ahn, J.H.; Graessle, S.; Wegener, S.; Tonukari, N.J.; Caballero, O.; Loidl, P.; Walton, J.D. A gene related to yeast HOS2 histone deacetylase affects extracellular depolymerase expression and virulence in a plant pathogenic fungus. Plant Cell 2001, 13, 1609–1624. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Pel, H.J.; de Winde, J.H.; Archer, D.B.; Dyer, P.S.; Hofmann, G.; Schaap, P.J.; Turner, G.; de Vries, R.P.; Albang, R.; Albermann, K.; et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat. Biotechnol. 2007, 25, 221–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terabayashi, Y.; Sano, M.; Yamane, N.; Marui, J.; Tamano, K.; Sagara, J.; Dohmoto, M.; Oda, K.; Ohshima, E.; Tachibana, K.; et al. Identification and characterization of genes responsible for biosynthesis of kojic acid, an industrially important compound from Aspergillus oryzae. Fungal Genet. Biol. 2010, 47, 953–961. [Google Scholar] [CrossRef]
- Marui, J.; Yamane, N.; Ohashi-Kunihiro, S.; Ando, T.; Terabayashi, Y.; Sano, M.; Ohashi, S.; Ohshima, E.; Tachibana, K.; Higa, Y.; et al. Kojic acid biosynthesis in Aspergillus oryzae is regulated by a Zn(II)(2)Cys(6) transcriptional activator and induced by kojic acid at the transcriptional level. J. Biosci. Bioeng. 2011, 112, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Steinfeld, I.; Shamir, R.; Kupiec, M. A genome-wide analysis in Saccharomyces cerevisiae demonstrates the influence of chromatin modifiers on transcription. Nat. Genet. 2007, 39, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Spikes, S.; Xu, R.; Nguyen, C.K.; Chamilos, G.; Kontoyiannis, D.P.; Jacobson, R.H.; Ejzykowicz, D.E.; Chiang, L.Y.; Filler, S.G.; May, G.S. Gliotoxin production in Aspergillus fumigatus contributes to host-specific differences in virulence. J. Infect. Dis. 2008, 197, 479–486. [Google Scholar] [CrossRef]
- Ferracin, L.M.; Fier, C.B.; Vieira, M.L.C.; Monteiro-Vitorello, C.B.; Varani, A.D.M.; Rossi, M.M.; Mueller-Santos, M.; Taniwaki, M.H.; Iamanaka, B.T.; Pelegrinelli Fungaro, M.H. Strain-specific polyketide synthase genes of Aspergillus niger. Int. J. Food Microbiol. 2012, 155, 137–145. [Google Scholar] [CrossRef]
- Bauer, I.; Varadarajan, D.; Pidroni, A.; Gross, S.; Vergeiner, S.; Faber, B.; Hermann, M.; Tribus, M.; Brosch, G.; Graessle, S. A Class 1 Histone Deacetylase with Potential as an Antifungal Target. MBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, M.; Jamnischek, A.; Weinzierl, G.; Ladendorf, O.; Huber, S.; Kahmann, R.; Kamper, J. The histone deacetylase Hda1 from Ustilago maydis is essential for teliospore development. Mol. Microbiol. 2002, 46, 1169–1182. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Wang, Z.-K.; Shao, W.; Ying, S.-H.; Feng, M.-G. Essential role of Rpd3-dependent lysine modification in the growth, development and virulence of Beauveria bassiana. Environ. Microbiol. 2018, 20, 1590–1606. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Tong, S.-M.; Shao, W.; Ying, S.-H.; Feng, M.-G. Pleiotropic effects of the histone deacetylase Hos2 linked to H4-K16 deacetylation, H3-K56 acetylation, and H2A-S129 phosphorylation in Beauveria bassiana. Cell. Microbiol. 2018, 20, e12839. [Google Scholar] [CrossRef] [PubMed]
- Pidroni, A.; Faber, B.; Brosch, G.; Bauer, I.; Graessle, S. A Class 1 Histone Deacetylase as Major Regulator of Secondary Metabolite Production in Aspergillus nidulans. Front. Microbiol. 2018, 9, 2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Studt, L.; Schmidt, F.J.; Jahn, L.; Sieber, C.M.K.; Connolly, L.R.; Niehaus, E.M.; Freitag, M.; Humpf, H.U.; Tudzynski, B. Two Histone Deacetylases, FfHda1 and FfHda2, Are Important for Fusarium fujikuroi Secondary Metabolism and Virulence. Appl. Environ. Microbiol. 2013, 79, 7719–7734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Xie, J.; Liu, X.; Wang, B.; Pan, L. Functional and transcriptomic analysis of the key unfolded protein response transcription factor HacA in Aspergillus oryzae. Gene 2016, 593, 143–153. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name * | Genotype | Disrupted Gene ID | HDAC Type (HDAC Class) |
|---|---|---|---|
| FGSC A1279 | Δku70::ptrA ΔpyrG:amds | ||
| ΔhosA | Δku70::ptrA ΔpyrG:amds ΔhosA::pyrG | An07g08380 | Classical HDAC (Class I) |
| ΔrpdA ht ** | Δku70::ptrA ΔpyrG:amds ΔrpdA::pyrG | An07g07850 | Classical HDAC (Class I) |
| ΔhdaA | Δku70::ptrA ΔpyrG:amds ΔhdaA::pyrG | An16g01840 | Classical HDAC (Class II) |
| ΔhosB | Δku70::ptrA ΔpyrG:amds ΔhosB::pyrG | An14g00560 | Classical HDAC (Class II) |
| Δhst2 | Δku70::ptrA ΔpyrG:amds Δhst2::pyrG | An11g06980 | sirtuins (Class III) |
| Δhst4 | Δku70::ptrA ΔpyrG:amds Δhst4::pyrG | An08g02500 | sirtuins (Class III) |
| ΔsirA | Δku70::ptrA ΔpyrG:amds ΔsirA::pyrG | An01g07390 | sirtuins (Class III) |
| Δhdas | Δku70::ptrA ΔpyrG:amds Δhdas::pyrG | An09g05710 |
| Name | Abbreviation | Concentration | Effects to Cellular Process |
|---|---|---|---|
| cargo red | CR | 1 mg/mL | cell walls stress |
| calcofluor white | CFW | 3 μg/mL | cell walls stress |
| menadione | MD | 45 μM | oxidative |
| dithiothreitol | DTT | 20 μM | reductive |
| camptothecin | CPT | 3 μM | obstruct DNA replication and transcription |
| hydroxy urea | HU | 10 M | obstruct DNA replication |
| trichostatin A | TSA | 5 μg/mL | histone inhibitor |
| voriconazole | VCZ | 20 ng/mL | cellular membrane stress |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Pan, L.; Wang, B.; Pan, L. The Histone Deacetylases HosA and HdaA Affect the Phenotype and Transcriptomic and Metabolic Profiles of Aspergillus niger. Toxins 2019, 11, 520. https://doi.org/10.3390/toxins11090520
Li X, Pan L, Wang B, Pan L. The Histone Deacetylases HosA and HdaA Affect the Phenotype and Transcriptomic and Metabolic Profiles of Aspergillus niger. Toxins. 2019; 11(9):520. https://doi.org/10.3390/toxins11090520
Chicago/Turabian StyleLi, Xuejie, Lijie Pan, Bin Wang, and Li Pan. 2019. "The Histone Deacetylases HosA and HdaA Affect the Phenotype and Transcriptomic and Metabolic Profiles of Aspergillus niger" Toxins 11, no. 9: 520. https://doi.org/10.3390/toxins11090520
APA StyleLi, X., Pan, L., Wang, B., & Pan, L. (2019). The Histone Deacetylases HosA and HdaA Affect the Phenotype and Transcriptomic and Metabolic Profiles of Aspergillus niger. Toxins, 11(9), 520. https://doi.org/10.3390/toxins11090520