[D-Leu1]MC-LR Has Lower PP1 Inhibitory Capability and Greater Toxic Potency than MC-LR in Animal and Plant Tissues



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
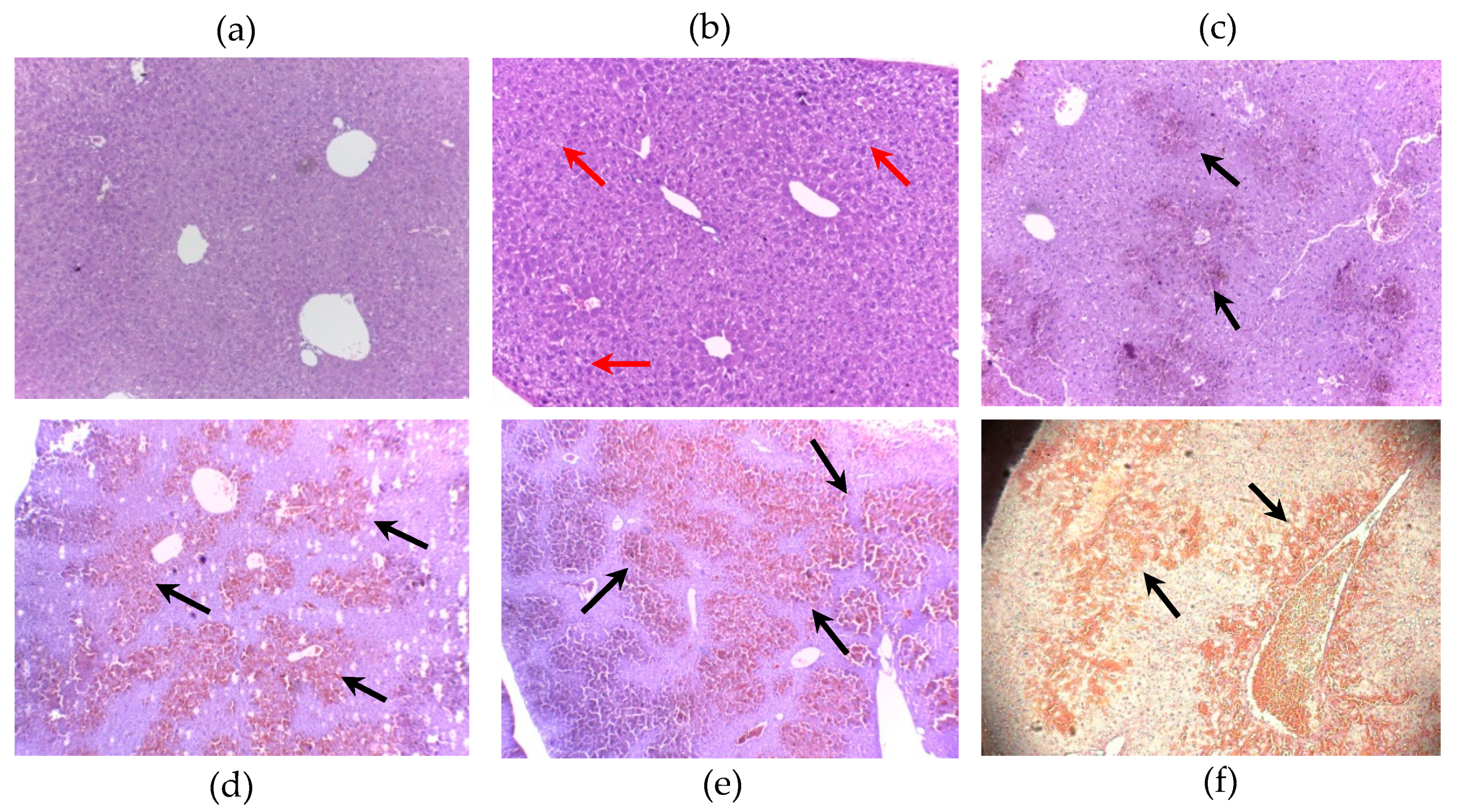
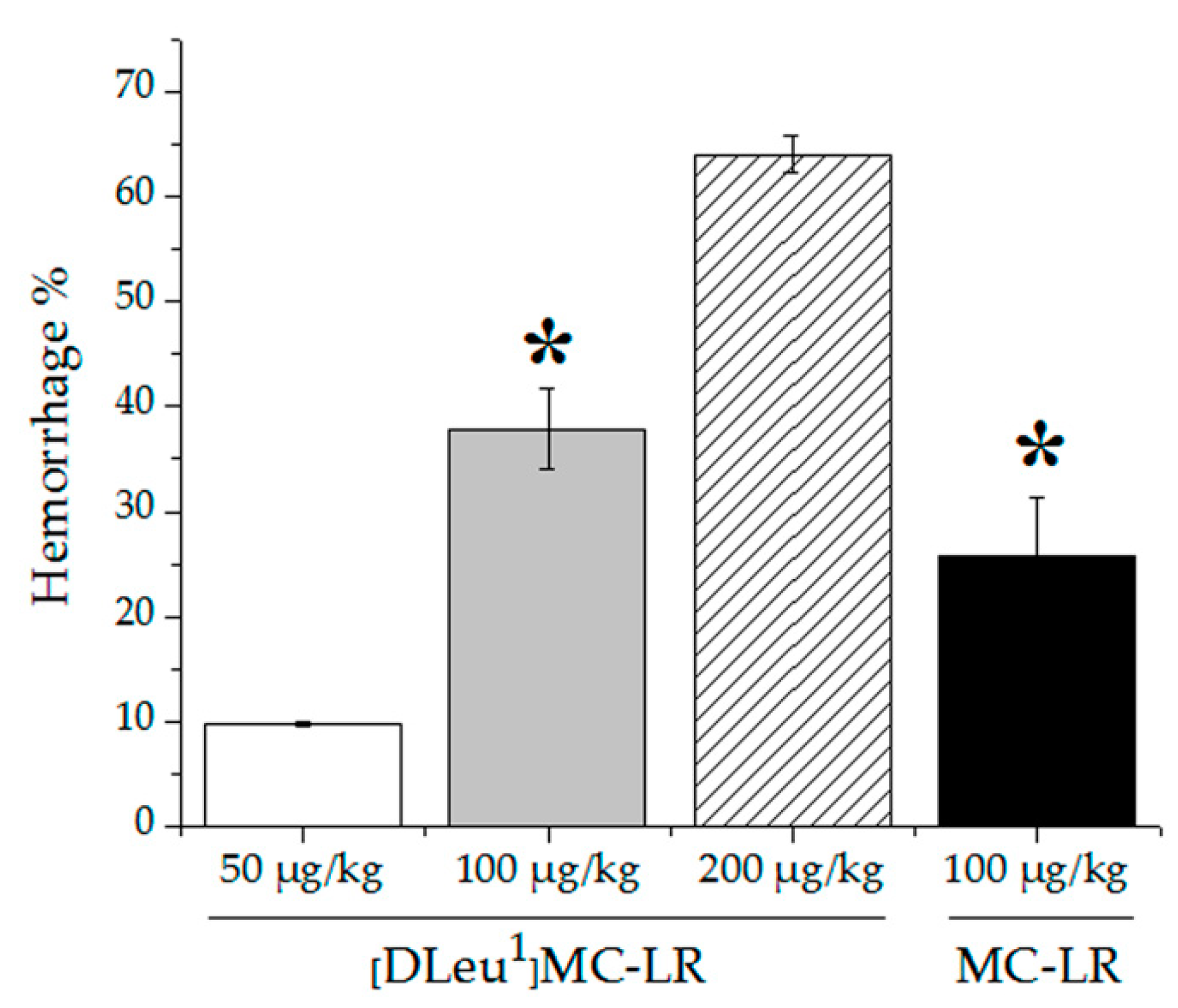
2.1. MC-LR and [DLeu1]MC-LR Acute Exposure in Mice
2.2. MC-LR and [DLeu1]MC-LR Acute Exposure in Phaseolus Vulgaris
2.2.1. Effects on Germination, Development and Morphology of Plants
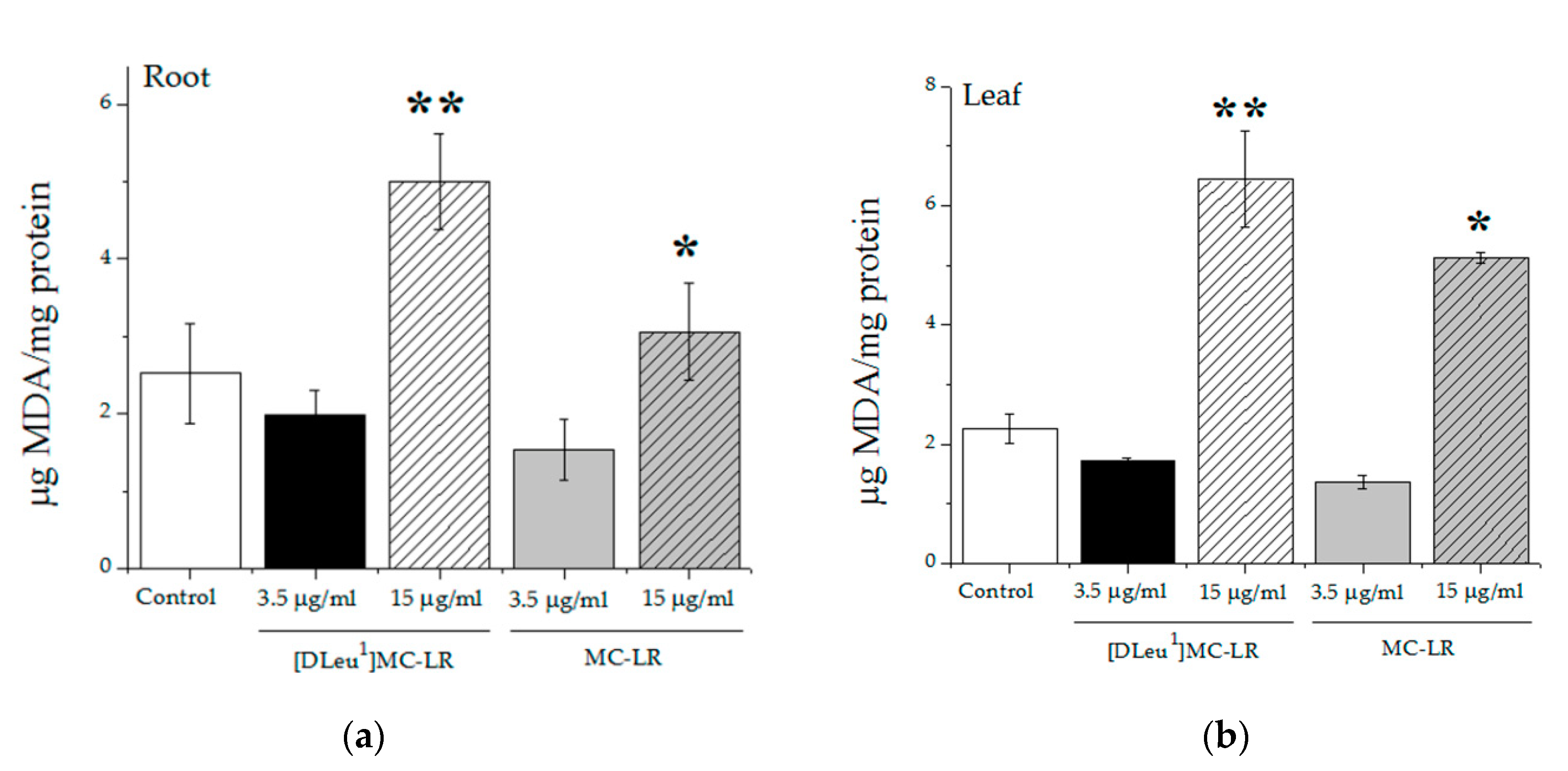
2.2.2. Biochemical Parameters in Treated Plants
2.3. MCLR and [D-Leu1]MC-LR Effects on Protein Phosphatase Activity
3. Discussion
4. Conclusions
5. Materials and Methods
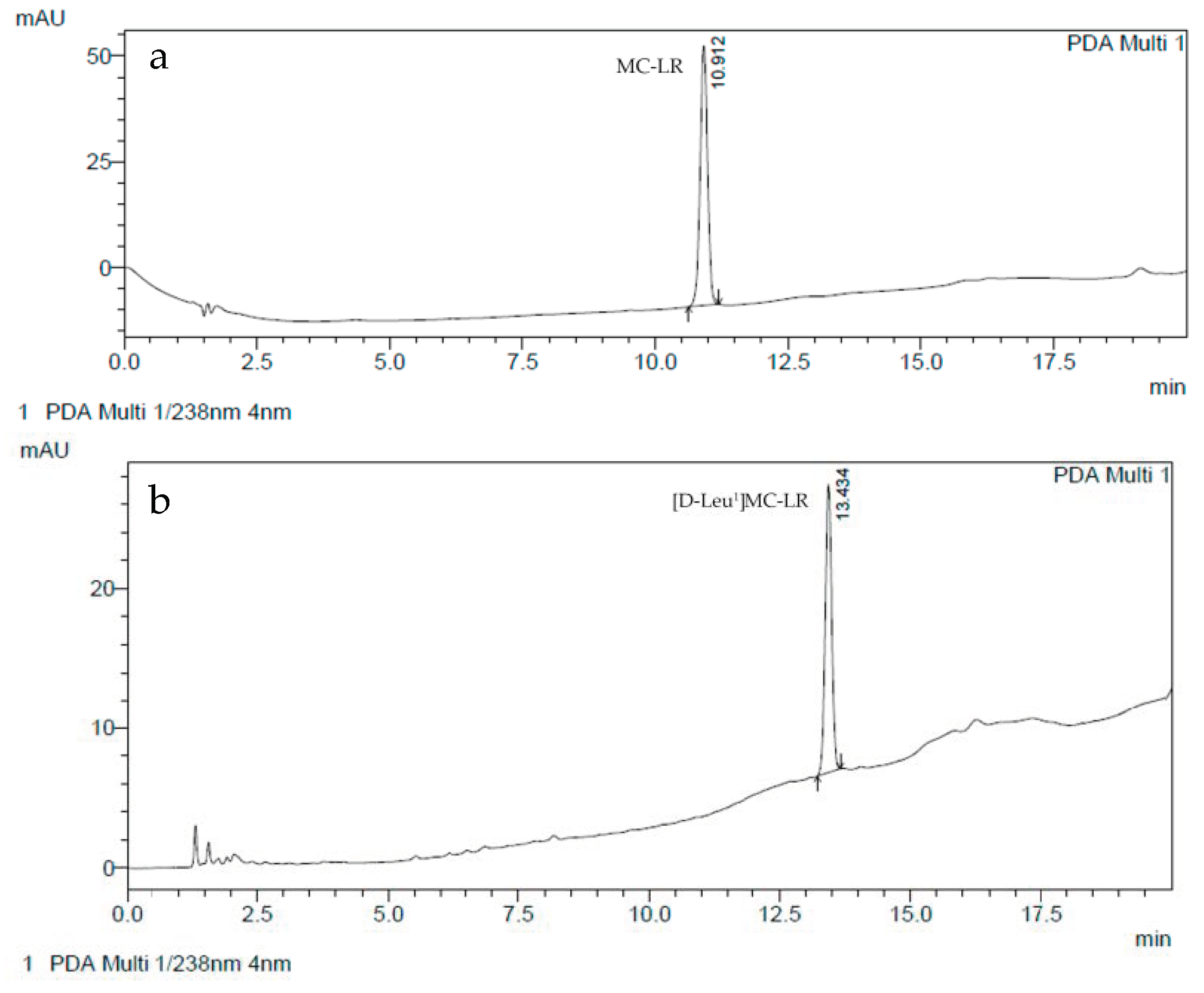
5.1. [D-Leu1]MC-LR and MC-LR Purification
5.2. Experimental Design
5.2.1. [D-Leu1] MC-LR and MC-LR Acute Exposure in Mice
5.2.2. [D-Leu1] MC-LR and MC-LR Acute Exposure in Beans
5.3. Analytical Determinations
5.3.1. Histological Studies
5.3.2. Biochemical Parameters Determined in Roots and Leaves
5.3.3. Protein Phosphatase Activity Assay
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pearl, H.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar]
- Carmichael, W.W. Fresh water cyanobacteria (blue-green algae) toxins. In Natural Toxins: Characterization, Pharmacology and Therapeutics; Ownby, C.A., Odell, G.V., Eds.; Pergamon Press: Oxford, UK, 1989; pp. 3–16. [Google Scholar]
- Codd, G.A.; Bell, S.G.; Brooks, W.P. Cyanobacterial toxins in water. Water Sci. Technol. 1989, 21, 1–13. [Google Scholar]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 1st ed.; World Health Organization: Geneva, Switzerland, 1999; pp. 55–161. ISBN 0-419-23930-8. [Google Scholar]
- Matthiensen, A.; Beattie, K.A.; Yunes, J.S.; Kaya, K.; Codd, G.A. [D-Leu1]-Microcystin-LR, from the cyanobacterium Microcystis RST 9501 and from a Microcystis bloom in the Patos Lagoon estuary, Brazil. Phytochemistry 2000, 55, 383–387. [Google Scholar]
- De León, L.; Yunes, J. First report of a Microcystis aeruginosa toxic bloom in La Plata River. Environ. Toxicol. 2001, 16, 110–112. [Google Scholar]
- Park, H.; Namikoshi, M.; Brittain, S.M.; Carmichael, W.W.; Murphy, T. [D-Leu1] microcystin-LR, a new microcystin isolated from waterbloom in a Canadian prairie lake. Toxicon 2001, 39, 855–862. [Google Scholar]
- Andrinolo, D.; Pereira, P.; Giannuzzi, L.; Aura, C.; Massera, S.; Caneo, M.; Caixach, J.; Braco, M.; Echenique, R. Ocurrence of microcystis aeruginosa and microcystins in Rio de La Plata River (Argentina). Acta Toxicol. Argent. 2007, 15, 8–14. [Google Scholar]
- Echenique, R.; Rodríguez, J.; Caneo, M.; Gianuzzi, L.; Barco, M.; Rivera, J.; Caixach, J.; Andrinolo, D. Microcystins in the drinking water supply in the cities of Ensenada and La Plata (Argentina). Anais XICBFic Livros Mus. Nac. Bras. 2008, 1, 125–129. [Google Scholar]
- Rosso, L.; Sedan, D.; Kolman, M.; Caixach, J.; Flores, C.; Oteiza, J.M.; Salerno, G.; Echenique, R.; Giannuzzi, L.; Andrinolo, D. Microcystis aeruginos strain [D-Leu1] Mcyst-LR producer, from Buenos Aires province, Argentina. J. Coast. Life Med. 2014, 2, 287–296. [Google Scholar]
- Craig, M.; Holmes, C.F.B. Freshwater hepatotoxins: Microcystin and nodularin, mechanisms of toxicity and effects on health. In Seafood and Freshwater Toxins; Botana, L.M., Ed.; Marcel Dekker: New York, NY, USA, 2000; pp. 643–671. [Google Scholar]
- Gehringer, M.M. Microcystin-LR and okadaic acid-induced cellular effects: A dualistic response. FEBS Lett. 2004, 57, 1–8. [Google Scholar]
- Falconer, I.R.; Beresford, A.M.; Runnegar, M.T.C. Evidence of liver damage by toxin from a bloom of the blue-green alga, Microcystis aeruginosa. Med. J. Aust. 1983, 1, 511–514. [Google Scholar] [PubMed]
- Carmichael, W.W. The toxins of cyanobacteria. Sci. Am. 1994, 270, 78–86. [Google Scholar] [CrossRef]
- Yoshida, T.; Makita, Y.; Nagata, S.; Tsutsumi, T.; Yoshida, F.; Sekijima, M.; Tamura, S.-I.; Ueno, Y. Acute oral toxicity of microcystin-LR, a cyanobacterial hepatotoxin, in mice. Nat. Toxins 1997, 5, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Ito, E.; Kondo, F.; Terao, K.; Harada, K. Neoplastic nodular formation in mouse liver induced by repeated intraperitoneal injections of MCLR. Toxicon 1997, 35, 1453–1457. [Google Scholar] [CrossRef]
- Solter, P.; Wollenberg, G.; Huang, X.; Chu, F.; Runnegar, M. Prolonged sublethal exposure to the protein phosphatase inhibitor, Microcystin-LR results in multiple dose-dependent hepatotoxic effects. Toxicol. Sci. 1998, 44, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Fawell, J.K.; Mitchell, R.E.; Everett, D.J.; Hill, R.E. The toxicity of cyanobacterial toxins in the mouse: I Microcystin-LR. Hum. Exp. Toxicol. 1999, 18, 162–167. [Google Scholar] [CrossRef]
- Guzman, R.E.; Solter, P.F. Hepatic oxidative stress following prolonged sublethal microcystin LR exposure. Toxicol. Pathol. 1999, 5, 582–588. [Google Scholar] [CrossRef]
- Guzman, R.E.; Solter, P.F. Characterization of sublethal MCLR exposure in mice. Vet. Pathol. 2002, 39, 17–26. [Google Scholar] [CrossRef]
- Milutinovic, A.; Zivin, M.; Zorc-Pleskovic, R.; Sedmak, B.; Suput, D. Nephrotoxic effects of chronic administration of microcystins-LR and -YR. Toxicon 2003, 42, 281–288. [Google Scholar] [CrossRef]
- Ding, W.; Ong, C. Role of oxidative stress and mitochondrial changes in cyanobacteria-induced apoptosis and hepatotoxicity. FEMS Microbiol. Lett. 2003, 220, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Xie, P.; Tang, R.; Zhang, X.; Li, L.; Li, D. In vivo studies on the toxic effects of microcystins on mitochondrial electron transport chain and ion regulation in liver and heart of rabbit. Comp. Biochem. Physiol. 2008, 148, 204–210. [Google Scholar] [CrossRef]
- Soares, R.; Cagido, V.; Ferraro, R.; Meyer-Fernandes, J.; Rocco, P.; Zin, W.; Azevedo, S. Effects of Microcystin-LR on mouse lungs. Toxicon 2007, 50, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Andrinolo, D.; Sedan, D.; Telese, L.; Aura, C.; Masera, S.; Giannuzzi, L.; Marra, C.; Alaniz, M.T. Recovery after damage produced by subchronic intoxication with the cyanotoxin microcystin LR. Toxicon 2008, 51, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Sedan, D.; Andrinolo, D.; Telese, L.; Giannuzzi, L.; Alaniz, M.T.; Marra, C.A. Alteration and recovery of the antioxidant system induced by sub-chronic exposure to Microcystin-LR in mice: Its relation to liver lipid composition. Toxicon 2010, 55, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Lezcano, N.; Sedan, D.; Lucotti, I.; Giannuzzi, L.; Vittone, L.; Andrinolo, D.; Mundiña-Weilenmann, C. Subchronic Microcystin-LR exposure increased hepatic apoptosis and induced compensatory mechanisms in mice. J. Biochem. Mol. Toxicol. 2012, 26, 131–138. [Google Scholar] [CrossRef]
- Sedan, D.; Giannuzzi, L.; Rosso, L.; Marra, C.A.; Andrinolo, D. Biomarkers of prolonged exposure to Microcystin-LR in mice. Toxicon 2013, 68, 9–17. [Google Scholar] [CrossRef]
- Sedan, D.; Laguens, M.; Copparoni, G.; Aranda, J.O.; Giannuzzi, L.; Marra, C.A.; Andrinolo, D. Hepatic and intestine alterations in mice after prolonged exposure to low oral doses of Microcystin-LR. Toxicon 2015, 104, 26–33. [Google Scholar] [CrossRef]
- Moreno, I.; Pichardo, S.; Jos, A.; Gomez-Amores, L.; Mate, A.; Vazquez, C.M.; Camean, A.M. Antioxidant enzyme activity and lipid peroxidation in liver and kidney of rats exposed to Microcystin-LR administered intraperitoneally. Toxicon 2005, 45, 395–402. [Google Scholar] [CrossRef]
- Kós, P.; Gorzó, G.; Surányi, G.; Borbély, G. Simple and efficient method for isolation and measurement of cyanobacterial hepatotoxins by plant tests (Sinapis alba L.). Anal. Biochem. 1995, 225, 49–53. [Google Scholar] [CrossRef]
- Abe, T.; Lawson, T.; Weyers, J.D.B.; Codd, G.A. Microcystin-LR inhibits photosynthesis of Phaseolus vulgaris primary leaves: Implications for current spray irrigation practice. New Phytol. 1996, 133, 651–658. [Google Scholar] [CrossRef]
- Kurki-Helasmo, K.; Meriluoto, J. Microcystin uptake inhibits growth and protein phosphatase activity in mustard (Sinapis alba L.) seedlings. Toxicon 1998, 36, 1921–1926. [Google Scholar] [CrossRef]
- Mc Elhiney, J.; Lawton, L.A.; Leifert, C. Investigations into the inhibitory effects of microcystins on plant growth, and the toxicity of plant tissues following exposure. Toxicon 2001, 39, 1411–1420. [Google Scholar] [CrossRef]
- Chen, J.Z.; Song, L.; Dai, E.; Gan, N.; Liu, Z. Effects of microcystins on the growth and the activity of superoxide dismutase and peroxidase of rape (Brassicanapus L.) and rice (Oryza sativa L.). Toxicon 2004, 43, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Malanga, G.; Giannuzzi, L.; Hernando, M. The possible role of microcystin (D-Leu1 MC-LR) as an antioxidant on Microcystis aeruginosa (Cyanophyceae). In vitro and in vivo evidence. Comp. Biochem. Physiol. Part C 2019, 225, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ramos, D.F.; Matthiensen, A.; Colvara, W.; Souza de Votto, A.P.; Trindade, G.S.; Almeida da Silva, P.E.; Yunes, J.S. Antimycobacterial and cytotoxicity activity of microcystins. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-M.; Lee, T.-H.; Lee, S.-J.; Huang, H.-B.; Huang, R.; Chou, H.-N. Comparison of protein phosphatase inhibition activities and mouse toxicities of microcystins. Toxicon 2006, 47, 742–746. [Google Scholar] [CrossRef]
- Goldberg, J.; Huang, H.-B.; Kwon, Y.-G.; Greengard, P.; Nairn, A.C.; Kuriyan, J. Three-dimensional structure of the catalytic subunit of protein serine/threonine phosphatase-1. Nature 1995, 376, 745–753. [Google Scholar] [CrossRef]
- Kaya, K. Toxicology of microcystins. In Toxic Microcystis, 1st ed.; Watanabe, M.F., Harada, K., Carmichael, W.W., Fujiki, H., Eds.; CRC Press: New York, NY, USA, 1996; pp. 175–202. [Google Scholar]
- Ito, E.; Takai, A.; Kondo, F.; Masui, H.; Imanishi, S.; Harada, K.I. Comparison of protein phosphatase inhibitory activity and apparent toxicity of microcystins and related compound. Toxicon 2002, 40, 1017–1025. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; The National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Marra, C.A.; de Alaniz, M.J.T.; Brenner, R.R. Modulation of D6 and D5 rat liver microsomal desaturase activities by dexamethasone-induced factor. Biochim. Biophys. Acta 1986, 879, 388–393. [Google Scholar] [CrossRef]
- Culling, C.F.A. Handbook of Histological Techniques, 3rd ed.; Butterworths: London, UK, 1975. [Google Scholar]
- Okawa, H.; Oshishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids, the pigments of photosynthetic biomembranes. In Methods in Enzymology; Douce, R., Packer, L., Eds.; Academic Press: Cambridge, MA, USA, 1987; pp. 350–382. [Google Scholar]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sedan, D.; Malaissi, L.; Vaccarini, C.A.; Ventosi, E.; Laguens, M.; Rosso, L.; Giannuzzi, L.; Andrinolo, D. [D-Leu1]MC-LR Has Lower PP1 Inhibitory Capability and Greater Toxic Potency than MC-LR in Animal and Plant Tissues. Toxins 2020, 12, 632. https://doi.org/10.3390/toxins12100632
Sedan D, Malaissi L, Vaccarini CA, Ventosi E, Laguens M, Rosso L, Giannuzzi L, Andrinolo D. [D-Leu1]MC-LR Has Lower PP1 Inhibitory Capability and Greater Toxic Potency than MC-LR in Animal and Plant Tissues. Toxins. 2020; 12(10):632. https://doi.org/10.3390/toxins12100632
Chicago/Turabian StyleSedan, Daniela, Luciano Malaissi, Cristian Adrián Vaccarini, Ezequiel Ventosi, Martín Laguens, Lorena Rosso, Leda Giannuzzi, and Darío Andrinolo. 2020. "[D-Leu1]MC-LR Has Lower PP1 Inhibitory Capability and Greater Toxic Potency than MC-LR in Animal and Plant Tissues" Toxins 12, no. 10: 632. https://doi.org/10.3390/toxins12100632
APA StyleSedan, D., Malaissi, L., Vaccarini, C. A., Ventosi, E., Laguens, M., Rosso, L., Giannuzzi, L., & Andrinolo, D. (2020). [D-Leu1]MC-LR Has Lower PP1 Inhibitory Capability and Greater Toxic Potency than MC-LR in Animal and Plant Tissues. Toxins, 12(10), 632. https://doi.org/10.3390/toxins12100632